
Биоэлектрические явления общая характеристика классификация реферат
Обновлено: 05.07.2024
Возбуждение представляет собой сложную совокупность физических, химических и физико-химических процессов, в результате которых происходит быстрое и кратковременное изменение электрического потенциала мембраны.
Позднее Дюбуа-Реймоном было установлено, что поврежденный участок мышцы имеет отрицательный заряд, а неповрежденный участок – положительный. При набрасывании нерва между поврежденным и неповрежденным участками мышцы возникает ток, который раздражает нерв и вызывает сокращение мышцы. Этот ток был назван током покоя, или током повреждения. Так было показано, что наружная поверхность мышечных клеток заряжена положительно по отношению к внутреннему содержимому.
Первая теория возникновения и поддержания мембранного потенциала была разработана Ю. Бернштейном (1902). Исходя из того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он показал, что величину мембранного потенциала можно определить, используя формулу Нернста.
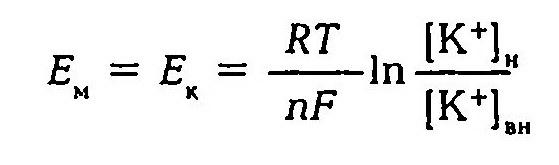
В 1949–1952 гг. А. Ходжкин, Э. Хаксли, Б. Катц создали со-временную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 -50 раз больше ионов калия, в 8–10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость. Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциалозависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией по обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле.
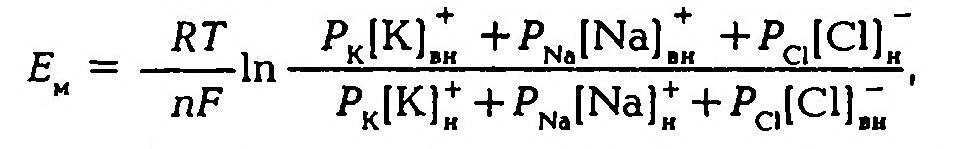
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т. е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы – крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд. Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде. Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки. То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей
ЛЕКЦИЯ № 2. Физиологические свойства и особенности функционирования возбудимых тканей 1. Физиологическая характеристика возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и
1. Физиологическая характеристика возбудимых тканей
1. Физиологическая характеристика возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.Раздражители – это
2. Законы раздражения возбудимых тканей
2. Законы раздражения возбудимых тканей Законы устанавливают зависимость ответной реакции ткани от параметров раздражителя. Эта зависимость характерна для высоко организованных тканей. Существуют три закона раздражения возбудимых тканей:1) закон силы
3. Понятие о состоянии покоя и активности возбудимых тканей
3. Понятие о состоянии покоя и активности возбудимых тканей О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма, нет
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.Процессы метаболизма в
2. Основные характеристики и законы возбудимых тканей
2. Основные характеристики и законы возбудимых тканей Основным свойством любой ткани является раздражимость, т. е. способность ткани изменять свои физиологические свойства и проявлять функциональные отправления в ответ на действие раздражителей.Раздражители – это
3. Понятие о состоянии покоя О и активности возбудимых тканей
3. Понятие о состоянии покоя О и активности возбудимых тканей О состоянии покоя в возбудимых тканях говорят в том случае, когда на ткань не действует раздражитель из внешней или внутренней среды. При этом наблюдается относительно постоянный уровень метаболизма.Основные
2 глава Патологические изменения в тканях
2 глава Патологические изменения в тканях При заболеваниях различных органов и систем в коже, соединительной ткани, миофасциальных структурах, надкостнице образуются патологические изменения , которые выявляются при наружном осмотре, поверхностной и глубокой
2 глава Патологические изменения в тканях
2 глава Патологические изменения в тканях При заболеваниях различных органов и систем в коже, соединительной ткани, миофасциальных структурах, надкостнице образуются патологические изменения, которые выявляются при наружном осмотре, поверхностной и глубокой
Глава 2. Физиология возбудимых тканей
Глава 2. Физиология возбудимых тканей Способность адаптироваться к постоянно изменяющимся условиям внешней среды является одним из основных признаков живых систем. В основе приспособительных реакций организма лежит раздражимость – способность реагировать на
Законы раздражения возбудимых тканей
Газообмен в легких и тканях
Газообмен в легких и тканях В легких происходит газообмен между поступающим в альвеолы воздухом и протекающей по капиллярам кровью. Интенсивному газообмену между воздухом альвеол и кровью способствует малая толщина описанного аэрогематического барьера. Альвеолярный
Инородное тело в тканях
Инородное тело в тканях Рыбья кость, иголки, осколки костей —
Абсцесс (скопление гноя в тканях)
Абсцесс (скопление гноя в тканях) • Взять по 20 г порошка клубней топинамбура, семян льна и травы донника желтого, по 10 г листьев мальвы, топинамбура и алтея; 2 ст. л. смеси залить 300 г кремниевой воды, варить 5 мин, настоять, укутав, 1 час, процедить, добавить 1 ст. л. меда.
Абсцесс, скопление гноя в тканях
Абсцесс, скопление гноя в тканях ? Взять по 20 г семян льна и травы донника, по 10 г листьев мальвы и алтея; 2 ст. л. смеси залить 300 мл воды, всыпать 1 ст. л. пищевой соды, варить 5 мин. Для компрессов.? Взять по 1 ст. л. луковиц лилии чисто-белой, корневищ купены душистой и корневищ
Абсцесс, скопление гноя в тканях
Абсцесс, скопление гноя в тканях — Взять по 20 г семян льна и травы донника лекарственного, по 10 г листьев мальвы и алтея; 2 ст. ложки смеси залить 300 г воды, варить 5 мин., охладить, процедить, добавить 3 ст. ложки яблочного уксуса. Для
Способность отвечать возбуждением на действие раздражителя является одним из наиболее характерных свойств живых клеток. Поэтому изучению проблемы возбудимости всегда уделялось большое внимание. Наблюдающийся в настоящее время стремительный рост наших знаний в этой области связан с широким использованием в физиологической практике новых, весьма тонких и точных приемов исследования, позволяющих раздражать одиночные возбудимые образования, регистрировать их электрическую активность, фиксировать на заданном уровне мембранный потенциал, непосредственно измерять ионные потоки через мембрану.
Содержание
Введение
1.Раздражимость. Классификация раздражителей
2. Потенциал покоя
3.Методы регистрации
4.Ионно-мембранная теория происхождения
5.Ионный механизм возбуждения
6.Потенциал действия, его компоненты
7.Изменение возбудимости при возбуждении
8.Отличия местного и распространяющегося возбуждения
9.Параметры возбудимости
Заключение
Список использованной литературы
Работа содержит 1 файл
доклад по физиологии.docx
1.Раздражимость. Классификация раздражителей
2. Потенциал покоя
4.Ионно-мембранная теория происхождения
5.Ионный механизм возбуждения
6.Потенциал действия, его компоненты
7.Изменение возбудимости при возбуждении
8.Отличия местного и распространяющегося возбуждения
Список использованной литературы
Способность отвечать возбуждением на действие раздражителя является одним из наиболее характерных свойств живых клеток. Поэтому изучению проблемы возбудимости всегда уделялось большое внимание. Наблюдающийся в настоящее время стремительный рост наших знаний в этой области связан с широким использованием в физиологической практике новых, весьма тонких и точных приемов исследования, позволяющих раздражать одиночные возбудимые образования, регистрировать их электрическую активность, фиксировать на заданном уровне мембранный потенциал, непосредственно измерять ионные потоки через мембрану.
Особенно выдающихся успехов за последние десятилетия добились исследователи, изучающие механизмы генерации биопотенциалов в нервных волокнах и клетках. Вершиной этих достижений, несомненно, являются исследования английских ученых Hodgkin и Huxley, которые развили выдвинутую еще в начале этого столетия концепцию Bernstein и сформулировали современную мембранную теорию возбуждения. Теория Hodgkin–Huxley получила всеобщее признание, и ее авторы были удостоены в 1964 г. Нобелевской премии.
Раздражимость. Классификация раздражителей
Основным свойством живых клеток является раздражимость, т. е. их способность реагировать изменением обмена веществ в ответ на действие раздражителей. Раздражением обозначается процесс воздействия на живую ткань агентов внешней по отношению к этой ткани среды — изменение среды существования. Раздражитель — это причина, способная вызвать возбуждение, т. е. агент внешней среды существования организма или внутренней среды организма, который, действуя на ткани, органы организма или на организм в целом, вызывает активную реакцию живого образования.
Возбудимость — свойство клеток отвечать на раздражение возбуждением. К возбудимым относят нервные, мышечные и некоторые секреторные клетки. Возбуждение — ответ ткани на ее раздражение, проявляющийся в специфической для нее функции (проведение возбуждения нервной тканью, сокращение мышцы, секреция железы) и неспецифических реакциях (генерация потенциала действия, метаболические изменения). Одним из важных свойств живых клеток является их электрическая возбудимость, т.е. способность возбуждаться в ответ на действие электрического тока. Высокая чувствительность возбудимых тканей к действию слабого электрического тока впервые была продемонстрирована Л. Гальвани в опытах на нервно-мышечном препарате задних лапок лягушки. Если к нервно-мышечному препарату лягушки приложить две соединенные между собой пластинки из различных металлов, например медь—цинк, таким образом, что бы одна пластинка касалась мышцы, а другая — нерва, то мышца будет сокращаться (первый опыт Л. Гальвани).
В конце XIX века благодаря работам Л. Германа, Э. Дюбуа-Раймона, Ю. Бернштейна стало очевидно, что электрические явления, которые возникают в возбудимых тканях, обусловлены электрическими свойствами клеточных мембран.
Реакция живой ткани зависит не только от силы раздражения извне, но и от времени действия этого раздражителя. Зависимости, существующие между длительностью раздражения и ответом раздражаемой ткани, могут быть сформулированы следующим образом: чем длительнее раздражение, тем сильнее (до известных пределов) и ответная реакция живой ткани. А общая сумма физиологических сдвигов, составляющих ответ ткани на раздражение, нарастает с нарастанием длительности раздражения без ограничений. Что касается возбуждения, связанного с функциональным отправлением ткани, то для проявления минимальной функциональной реакции требуется определенная критическая, пороговая длительность раздражителя. Увеличение длительности внешнего ''воздействия за пределами порога ведет к увеличению и функциональных отправлении ткани. Дальнейшее увеличение длительности раздражения уже не ведет к нарастанию функциональных отправлений ткани.
Таким образом, для того чтобы вызвать ответную реакцию органа, раздражители должны иметь достаточную силу. Возбудимость определяется минимальной (пороговой) силой раздражителя, вызывающей ответную реакцию. Причем чем выше возбудимость, тем меньше может быть пороговая сила раздражителя.
· физические (электричество, свет, звук, механические воздействия и т.д.);
· химические (кислоты, щелочи, гормоны и т.д.);
· физико-химические (осмотическое давление, парциальное давление газов);
· биологические (пища для животного, особь другого пола);
· социальные (слово для человека).
2. По месту воздействия:
· подпороговые (не вызывающие ответной реакции);
· пороговые (раздражители мин. силы, при которой возникает возбуждение);
· сверхпороговые (силой выше пороговой).
4. По физиологическому характеру:
- адекватные (физиологичные для данной клетки или рецептора, которые, приспособились к нему в процесс эволюции, например, свет для фоторецепторов глаза). Адекватными раздражителями называются агенты, действующие на определенные возбудимые системы организма в естественных условиях существования этого последнего. Так, например, для органа зрения адекватным раздражителем будет определенный участок шкалы электромагнитных волн – видимые световые лучи, для органа слуха - колебания воздушных волн известной частоты (от 16 до 20000 Гц), для температурных рецепторов кожи—изменения температуры внешней среды
- неадекватные. К неадекватным раздражителям относятся агенты внешней среды, не являющиеся в натуральных условиях существования организма средством возбуждения различных органов чувств, но, тем не менее, способные при достаточной силе и длительности своего воздействия вызвать возбуждение в раздражаемой ткани.
Если реакция на раздражитель является рефлекторной, то выделяют также:
- безусловно-рефлекторные раздражители;
- условно-рефлекторные.
Мембранный потенциал (или потенциал покоя) – это разность потенциалов между наружной и внутренней поверхностью мембраны в состоянии относительного физиологического покоя. Потенциал покоя возникает в результате двух причин:
1) неодинакового распределения ионов по обе стороны мембраны;
2) избирательной проницаемости мембраны для ионов. В состоянии покоя мембрана неодинаково проницаема для различных ионов. Клеточная мембрана проницаема для ионов K, малопроницаема для ионов Na и непроницаема для органических веществ.
Содержимое клетки заряжено отрицательно по отношению к внеклеточному пространству. Основная причина возникновения на мембране электрического потенциала (мембранного потенциала) — существование специфических ионных каналов. Транспорт ионов через каналы происходит по градиенту концентрации или под действием мембранного потенциала. В невозбужденной клетке часть К+-каналов находится в открытом состоянии и ионы К+ постоянно диффундируют из нейрона в окружающую среду (по градиенту концентрации). Покидая клетку, ионы К+ уносят положительный заряд, что создает потенциал покоя равный примерно -60 мВ. Из коэффициентов проницаемости различных ионов видно, что каналы, проницаемые для Na+ и Cl-, преимущественно закрыты. Ионы фосфата и органические анионы, например белки, практически не могут проходить через мембраны. Мембранный потенциал нервной клетки в первую очередь определяется ионами К+, которые вносят основной вклад в проводимость мембраны.
1. Мембранный потенциал быстро определяют, измеряя разность потенциалов между двумя одинаковыми электродами, один из которых введен в клетку, другой помещен в омывающую ее жидкость. Электроды соединены с усилителем, увеличивающим амплитуду регистрируемого потенциала; эта амплитуда определяется при помощи измерителя напряжения типа осциллоскопа.
2. О существовании электрического заряда на поверхностной мембране в физиологии известно очень давно, но только обнаруживали его другим способом – в виде так называемого тока покоя.
Ток покоя возникает в любой живой структуре между поврежденным ее участком и неповрежденной поверхностью.
Если перерезать нерв или мышцу, и один электрод приложить к поперечному разрезу, а другой – к поверхности, соединив их с гальванометром, то гальванометр покажет ток, который всегда течет от нормальной, неповрежденной поверхности к поперечному разрезу.
Ток покоя и мембранный потенциал – проявление одного и того же свойства мембраны; причина появления тока покоя заключается в том, что при повреждении клетки фактически возникает возможность соединить один электрод с внутренней стороной мембраны, а другой – с наружной ее поверхностью.
В идеальных условиях при повреждении должна была бы регистрироваться разность потенциалов = мембранному потенциалу. Этого, как правило, не происходит, т.к. часть тока не идет через гальванометр, а шунтируется по межклеточным пространствам, окружающей жидкости и т.п.
Величина трансмембранной разности потенциалов, которая может быть создана таким процессом, предсказывается уравнением Нернста:
где R – газовая постоянная, T – абсолютная температура, F – число Фарадея, [K]вн:[K]нар – отношение концентрации калия внутри и снаружи клетки.
Концентрация калия снаружи – в межклеточной жидкости – примерно = таковой в крови. Внутриклеточную концентрацию можно примерно определить, пользуясь некоторыми аналитическими приемами либо измерениями с помощью калий-селективных электродов.
Ионы натрия в небольшом количестве проникают внутрь клетки и заряжают внутреннюю поверхность мембраны положительно, создавая встречную разность потенциалов. Хотя эта разность незначительна, она может снизить истинную величину мембранного потенциала.
Ионно-мембранная теория происхождения
Сущность этой теории заключается в том, что потенциал покоя и потенциал действия являются по своей природе мембранными потенциалами, обусловленными полупроницаемыми свойствами клеточной мембраны и неравномерным распределением ионов между клеткой и средой, которое поддерживается механизмами активного переноса, локализованными в самой мембране. Основные положения.
Электрические процессы возникают на плазматической мембране клетки, которая состоит из бимолекулярного слоя липидов (остов мембраны) и белков, которые выполняют различные функции в мембране: рецепторную, ферментативную, образуют в ней каналы и насосы.

Лекции
Лабораторные
Справочники
Эссе
Вопросы
Стандарты
Программы
Дипломные
Курсовые
Помогалки
Графические
Доступные файлы (1):
МИНИСТЕРСТВО ОБРАЗОВАНИЯ И НАУКИ РК
ВОСТОЧНО-КАЗАХСТАНСКИЙ ГОСУДАРСТВЕННЫЙ УНИВЕРСИТЕТ им. С. АМАНЖОЛОВА
Тема: БИОЭЛЕКТРИЧЕСТВО В ЖИВОТНЫХ КЛЕТКАХ
Выполнила: Свечникова Татьяна
Проверила: Сапронова Е.А
г. Усть-Каменогорск 2008
1. Биоэлектрические потенциалы. 4
1.1. Краткие исторические сведения. 4
1.2. Потенциал покоя (ПП, мембранный потенциал покоя). 6
1.3. Потенциал действия (ПД). 7
1.4. Постсинаптические потенциалы (ПСП) 9
1.5. Генераторные потенциалы 9
2. Мембранная теория возбуждения 11
3. Натрий-калиевый насос. 14
Список литературы. 17
Введение.
Биоэлектричество - электрические явления и процессы, возникающие в живых тканях организма. Также - воздействие электрического тока на живые ткани.
Биоэлектричество это естественные электрические процессы в живых организмах, лежащие в основе многих физиологических и поведенческих реакций. К проблемам биоэлектричества относят также все эффекты, возникающие в организме на различных его уровнях при воздействии электричества от внешних источников.
Исследование биоэлектричества имеет большое значение для понимания физико-химических и физиологических процессов в живых системах.
Возбуждение характеризуется совокупностью электрических, температурных, химических, функциональных и структурных изменений живой клетки. Среди них особо важное значение имеют биоэлектрические явления.
^
1. Биоэлектрические потенциалы.
Биоэлектрические потенциалы - биоэлектрические явления, возникающие в тканях и отдельных клетках человека, животных и растений, важнейшие компоненты процессов возбуждения и торможения.
^
1.1. Краткие исторические сведения.
Первые данные о существовании биоэлектрических потенциалов ("животного электричества") были получены в 3-й четверти 18 в. при изучении природы "удара", наносимого некоторыми рыбами с электрическими органами при защите или нападении.
К этому же времени относится начало исследований итальянского физиолога и врача Л. Гальвани, заложивших основу учения о биоэлектрических потенциалах. Многолетний научный спор (1791—97) между Л. Гальвани и физиком А. Вольта о природе "животного электричества" завершился двумя крупными открытиями: были получены факты о существовании биоэлектрических явлений в живых тканях и открыт новый принцип получения электрического тока с помощью разнородных металлов — создан гальванический элемент (вольтов столб).
Правильная оценка наблюдений Гальвани стала возможной лишь после применения достаточно чувствительных электроизмерительных приборов — гальванометров. Первые такие исследования были проведены итальянским физиком К. Маттеуччи (1837). Систематическое изучение биоэлектрических потенциалов было начато немецким физиологом Э. Дюбуа-Реймоном (1848), который доказал существование биоэлектрических потенциалов в нервах и мышцах в покое и при возбуждении.
В 1886 немецкий физиолог Ю. Бернштейн проанализировал форму потенциала действия; французский учёный Э. Ж. Марей (1875) применил для записи колебаний потенциалов бьющегося сердца капиллярный электрометр. Русский физиолог Н. Е. Введенский использовал (1883) для прослушивания ритмических разрядов импульсов в нерве и мышце телефон, а голландский физиолог В. Эйнтховен (1903) ввёл в эксперимент и клиническую практику струнный гальванометр — высокочувствительный и малоинерционный прибор для регистрации электрических токов в тканях.
Значительный вклад в изучение биоэлектрических потенциалов внесли русские физиологи: В. В. Правдич-Неминский (1913—21) впервые зарегистрировал электроэнцефалограмму, А. Ф. Самойлов (1929) исследовал природу нервно-мышечной передачи возбуждения, а Д. С. Воронцов (1932) открыл следовые колебания биоэлектрических потенциалов, сопровождающие потенциал действия в нервных волокнах.
Дальнейший прогресс в изучении биоэлектрических потенциалов был тесно связан с успехами электроники, позволившими применить в физиологическом эксперименте электронные усилители и осциллографы.
Важное значение для выяснения механизмов генерации биоэлектрических потенциалов имело использование гигантских нервных волокон головоногих моллюсков, главным образом кальмара. Диаметр этих волокон в 50 — 100 раз больше, чем у позвоночных животных, он достигает 0,5—1 мм, что позволяет вводить внутрь волокна микроэлектроды, инъецировать в протоплазму различные вещества и т.п. Изучение ионной проницаемости мембраны гигантских нервных волокон позволило английским физиологам А. Ходжкину, А. Хаксли и Б. Катцу (1947—1952) сформулировать современную мембранную теорию возбуждения.
Различают следующие основные виды Б. п. нервных и мышечных клеток: потенциал покоя, потенциал действия, возбуждающие и тормозные постсинаптические потенциалы, генераторные потенциалы.
^
1.2. Потенциал покоя (ПП, мембранный потенциал покоя).
У живых клеток в покое между внутренним содержимым клетки и наружным раствором существует разность потенциалов (ПП) порядка 60—90мв, которая локализована на поверхностной мембране. Внутренняя сторона мембраны заряжена электроотрицательно по отношению к наружной. ПП обусловлен избирательной проницаемостью покоящейся мембраны для ионов К + . Концентрация К + в протоплазме примерно в 50 раз выше, чем во внеклеточной жидкости, поэтому, диффундируя из клетки, ионы выносят на наружную сторону мембраны положительные заряды, при этом внутренняя сторона мембраны, практически не проницаемой для крупных органических анионов, приобретает отрицательный потенциал. Поскольку проницаемость мембраны в покое для Na + примерно в 100 раз ниже, чем для К + , диффузия натрия из внеклеточной жидкости (где он является основным катионом) в протоплазму мала и лишь незначительно снижает ПП, обусловленный ионами К + . В скелетных мышечных волокнах в возникновении потенциала покоя важную роль играют также ионы Cl - , диффундирующие внутрь клетки. Следствием ПП является ток покоя, регистрируемый между поврежденным и интактным участками нерва или мышцы при приложении отводящих электродов. Мембраны нервных и мышечных клеток (волокон) способны изменять ионную проницаемость в ответ на сдвиги мембранного потенциала. При увеличении ПП (гиперполяризация мембраны) проницаемость поверхностных клеточных мембран для Na + и К + падает, а при уменьшении ПП (деполяризация) она возрастает, причём скорость изменений проницаемости для Na + значительно превышает скорость увеличения проницаемости мембраны для К + .
1.3. Потенциал действия (ПД).
Все раздражители, действующие на клетку, вызывают в первую очередь снижение ПП; когда оно достигает критического значения (порога), возникает активный распространяющийся ответ — ПД. Во время восходящей фазы ПД кратковременно извращается потенциал на мембране: её внутренняя сторона, заряженная в покое электроотрицательно, приобретает в это время положительный потенциал. Достигнув вершины, ПД начинает падать (нисходящая фаза ПД), и потенциал на мембране возвращается к уровню, близкому к исходному, — ПП. Полное восстановление ПП происходит только после окончания следовых колебаний потенциала — следовой деполяризации или гиперполяризации, длительность которых обычно значительно превосходит продолжительность пика ПД. Согласно мембранной теории, деполяризация мембраны, вызванная действием раздражителя, приводит к усилению потока Na + внутрь клетки, что уменьшает отрицательный потенциал внутренней стороны мембраны — усиливает её деполяризацию. Это, в свою очередь, вызывает дальнейшее повышение проницаемости для Na + и новое усиление деполяризации и т.д. В результате такого взрывного кругового процесса, т. н. регенеративной деполяризации, происходит извращение мембранного потенциала, характерное для ПД. Повышение проницаемости для Na + очень кратковременно и сменяется её падением, а следовательно, уменьшением потока Na + внутрь клетки. Проницаемость для К + , в отличие от проницаемости для Na + , продолжает увеличиваться, что приводит к усилению потока К + из клетки. В результате этих изменений ПД начинает падать, что ведёт к восстановлению ПП. Таков механизм генерации ПД в большинстве возбудимых тканей. Существуют, однако, клетки (мышечные волокна ракообразных, нервные клетки у ряда брюхоногих моллюсков, некоторые растительные клетки), у которых восходящая фаза ПД обусловлена повышением проницаемости мембраны не для ионов Na + , а для ионов Ca + . Своеобразен также механизм генерации ПД в мышечных волокнах сердца, для которых характерно длительное плато на нисходящей фазе ПД. Неравенство концентраций ионов К + и Na + (или Ca + ) внутри и снаружи клетки (волокна) поддерживается специальным механизмом (т. н. "натриевым насосом" ), выталкивающим ионы Na + из клетки и нагнетающим ионы К + в протоплазму, требующим затраты энергии, которая черпается клеткой в процессах обмена веществ.
Амплитуда ПД большинства нервных и мышечных волокон примерно одинакова: 110—120 мв. Длительность ПД варьирует в широких пределах: у теплокровных животных длительность ПД нервных волокон, наиболее быстро проводящих возбуждение, — 0,3—0,4 мсек, у волокон же мышц сердца — 50—600 мсек. В растительных клетках пресноводной водоросли хара ПД продолжается около 20 сек. Характерной особенностью ПД, отличающей его от других форм ответа клетки на раздражение, является то, что он подчиняется правилу "всё или ничего", т. е. возникает только при достижении раздражителем некоторого порогового значения, и дальнейшее увеличение интенсивности раздражителя уже не сказывается ни на амплитуде, ни на продолжительности ПД. Потенциал действия — один из важнейших компонентов процесса возбуждения. В нервных волокнах он обеспечивает проведение возбуждения от чувствительных окончаний ( рецепторов ) к телу нервной клетки и от неё — к синаптическим окончаниям (см. Синапсы ), расположенным на различных нервных, мышечных или железистых клетках. Поступая в эффекторные окончания, ПД вызывает выделение (секрецию) определённой порции специфических химических веществ, т. н. медиаторов , оказывающих возбуждающее или тормозящее влияние на соответствующие клетки. В мышечных волокнах распространяющийся ПД вызывает цепь физико-химических реакций, лежащих в основе процесса сокращения мышц. Проведение ПД вдоль нервных и мышечных волокон осуществляется т. н. локальными токами, или токами действия, возникающими между возбуждённым (деполяризованным) и соседними с ним покоящимися участками мембраны (см. Возбуждение ). Токи действия регистрируются обычными внеклеточными электродами; при этом кривая имеет двухфазный характер: первая фаза соответствует приходу ПД под ближний электрод, вторая — под дальний электрод.
1.4. Постсинаптические потенциалы (ПСП)
Возникают в участках мембраны нервных или мышечных клеток, непосредственно граничащих с синаптическими окончаниями. Они имеют амплитуду порядка нескольких мв и длительность 10—15 мсек. ПСП подразделяются на возбуждающие (ВПСП) и тормозные (ТПСП). ВПСП представляют собой местную деполяризацию постсинаптической мембраны, обусловленную действием соответствующего медиатора (например, ацетилхолина в нервно-мышечном соединении). При достижении ВПСП некоторого порогового (критического) значения в клетке возникает распространяющийся ПД. ТПСП выражается местной гиперполяризацией мембраны, обусловленной действием тормозного медиатора. В отличие от ПД, амплитуда ПСП постепенно увеличивается с увеличением количества выделившегося из нервного окончания медиатора. ВПСП и ТПСП суммируются друг с другом при одновременном или последовательном поступлении нервных импульсов к окончаниям, расположенным на мембране одной и той же клетки.
1.5. Генераторные потенциалы
Возникают в мембране чувствительных нервных окончаний — рецепторов. Они внешне сходны с ВПСП — их амплитуда порядка нескольких мв и зависит от силы приложенного к рецептору раздражения. Когда генераторный потенциал достигает порогового (критического) значения, в соседнем участке мембраны нервного волокна возникает распространяющийся ПД. Ионный механизм генераторных потенциалов ещё недостаточно изучен.
Наряду с перечисленными относительно быстро развивающимися Б. п., в нервных клетках, волокнах гладких мышц и некоторых растительных клетках регистрируются также очень медленные колебания мембранного потенциала неизвестной природы, причём на гребне волны деполяризации мембраны часто возникают разряды импульсов.
2. Мембранная теория возбуждения
Общепринятая в физиологии теория возбуждения мышечных и нервных клеток. Основа мембранной теории возбуждения — представление о том, что при раздражении возбудимой клетки в её поверхностной мембране происходит молекулярная перестройка, которая приводит к изменению проницаемости мембраны и появлению трансмембранных ионных токов. Источником энергии для этих токов служит постоянно существующее неравномерное распределение основных неорганических ионов между цитоплазмой и внеклеточной средой: накопление в клетке ионов K + и выведение из неё ионов Na + и Cl - .
Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю. Бернштейном (1902) и развиты английскими учёными: П. Бойлом и Э. Конуэем (1941) и А. Ходжкином, Б. Кацем, А. Хаксли (1949).
Описанные ионные процессы ведут (помимо появления распространяющегося импульса нервного ) к накоплению в клетке некоторого количества Na + и потере ею части K + . Эти изменения столь незначительны по сравнению с существующими между цитоплазмой и внеклеточной средой ионными градиентами, что клетка может генерировать огромное число импульсов без немедленного восстановления нарушенных ионных соотношений за счёт активного транспорта ионов , удаляющего из клетки избыток Na + и насасывающего в неё недостающее количество K + .
При химическом раздражении специфических изменения ионной проницаемости мембраны также приводят к развитию трансмембранных ионных токов. Такие изменения развиваются в межнейронных и нервно-мышечных синапсах и лежат в основе синаптической передачи с помощью медиаторов .
Существо перестроек в мембране, обеспечивающих появление ионных токов, — наименее ясная часть М. т. в. Полагают, что перенос ионов через мембрану происходит либо по системе пор (входы в которые в состоянии покоя закрыты, возможно ионами Ca 2+ , и открываются под действием внешнего раздражения), либо при помощи особых молекул-переносчиков, которые связывают ион на одной стороне мембраны и освобождают его на другой.
3. Натрий-калиевый насос.
Попытки выяснить связь обмена веществ с движением ионов через мембрану привели к открытию так называемого натрий-калиевого насоса.
Проведение серии импульсов по нервному волокну сопровождается обогащением протоплазмы ионами Nа и потерей ионов К. Для гигантского аксона кальмара подсчитано, что во время одиночного нервного импульса через каждый квадратный микрон мембраны внутрь протоплазмы поступает около 20 000 ионов Nа и столько же ионов К покидает волокно.
Такой механизм и получил название натрий-калиевого насоса. Деятельность его связана с затратой энергии обмена веществ. Действительно, для того чтобы выводить ионы Nа из протоплазмы в наружный раствор, где их концентрация значительно превышает внутриклеточную, необходимо совершить определенную работу. В покое эта работа невелика, так как натриевая проницаемость покоящейся мембраны имеет очень низкие величины. При возбуждении же усиленное поступление ионов Nа внутрь протоплазмы активирует работу насоса, что обеспечивает восстановление нарушенных концентрационных градиентов. Следует, однако, подчеркнуть, что этот восстановительный процесс протекает очень медленно — в течение многих минут и даже часов.
Непосредственным источником энергии для работы насоса являются богатые энергией фосфорные соединения — АТФ и фосфаген (креатин или аргининфосфат). При расщеплении одной грамм-молекулы АТФ выделяется 8000—10 000 кал. Расщепление это осуществляется ферментом аденозинтрифосфатазой, локализованной в мембране, причем указанный фермент активируется ионами Nа и К.
Нарушение синтеза АТФ и фосфагена при отравлении нервных волокон ядами, выключающими гликолиз, дыхание (цианиды) или окислительное фосфолирование (динитрофенол, азид натрия), приводит к нарушению работы натрий-калиевого насоса. В результате протоплазма отравленного нервного волокна обогащается натрием и теряет калий. Вместе с тем происходит снижение, вплоть до полнлго исчезновения, потенциала покоя. Волокно утрачивает возбудимость и проводимость. Если же в нервное волокно ввести некоторое количество аденозинтрифосфата и фосфагена, то активный транспорт ионов Nа и К восстанавливается.
Заключение.
Живой организм является не только генератором биопотенциалов, но и проводником электрического тока, причем изменение степени электропроводности живых тканей в зависимости от их жизнедеятельности может служить показателем жизнеспособности (состояния) клеток или тканей.
Список литературы.
2. Гальвани Л. и Вольта А., Избр. работы о животном электричестве, М. — Л., 1937;
Первая теория возникновения и поддержания мембранног потенциала была разработана Ю.Бернштейном (1902). Исходя и того, что мембрана клеток обладает высокой проницаемостью для ионов калия и малой проницаемостью для других ионов, он пока зал, что величину мембранного потенциала можно определить используя формулу Нернста

где Е — разность потенциалов между внутренней и наружной сторонами мембраны; £ — равновесный потенциал для ионов калия; R — газовая постоянная; Т — абсолютная температура; л — валентность иона; F — число Фарадея; [К*]вд — внутренняя и [К*] — наружная концентрация ионов калия.
В 1949—1952 гг. А.Ходжкин, Э.Хаксли, Б.Катц создали современную мембранно-ионную теорию, согласно которой мембранный потенциал обусловлен не только концентрацией ионов калия, но и натрия и хлора, а также неодинаковой проницаемостью для этих ионов мембраны клетки. Цитоплазма нервных и мышечных клеток содержит в 30 —50 раз больше ионов калия, в 8—10 раз меньше ионов натрия и в 50 раз меньше ионов хлора, чем внеклеточная жидкость.
Проницаемость мембраны для ионов обусловлена ионными каналами, макромолекулами белка, пронизывающими липидный слой. Одни каналы открыты постоянно, другие (потенциалозависимые) открываются и закрываются в ответ на изменения МП. Потенциал зависимые каналы подразделяются на натриевые, калиевые, кальциевые и хлорные. В состоянии физиологического покоя мембрана нервных клеток в 25 раз более проницаема для ионов калия, чем для ионов натрия.
Таким образом, согласно обновленной мембранной теории асимметричное распределение ионов по обе стороны мембраны и связанное с этим создание и поддержание мембранного потенциала обусловлено как избирательной проницаемостью мембраны для различных ионов, так и их концентрацией но обе стороны от мембраны, а более точно величину мембранного потенциала можно рассчитать по формуле:

где PK, PN4, PCl— проницаемость для ионов калия, натрия и хлора.
Поляризация мембраны в покое объясняется наличием открытых калиевых каналов и трансмембранным градиентом концентраций калия, что приводит к выходу части внутриклеточного калия в окружающую клетку среду, т.е. к появлению положительного заряда на наружной поверхности мембраны. Органические анионы — крупномолекулярные соединения, для которых мембрана клетки непроницаема, создают на внутренней поверхности мембраны отрицательный заряд.
Поэтому чем больше разница концентраций калия по обе стороны от мембраны, тем больше его выходит и тем выше значения МП. Переход ионов калия и натрия через мембрану по их концентрационному градиенту в конечном итоге должен был бы привести к выравниванию концентрации этих ионов внутри клетки и в окружающей ее среде.
Но в живых клетках этого не происходит, так как в клеточной мембране имеются натрий-калиевые насосы, которые обеспечивают выведение из клетки ионов натрия и введение в нее ионов калия, работая с затратой энергии. Они принимают и прямое участие в создании МП, так как за единицу времени ионов натрия выводится из клетки больше, чем вводится калия (в соотношении 3:2), что обеспечивает постоянный ток положительных ионов из клетки.
То что выведение натрия зависит от наличия метаболической энергии, доказывается тем, что под действием динитрофенола, который блокирует метаболические процессы, выход натрия снижается примерно в 100 раз. Таким образом, возникновение и поддержание мембранного потенциала обусловлено избирательной проницаемостью мембраны клетки и работой натрий-калиевого насоса.
Биоэлектрические явления
В частности, Б. я. обусловливают возникновение возбуждения и его проведение по нервным волокнам, являются причиной процессов сокращения мышечных волокон скелетных, гладких и сердечных мышц, выделительной функции железистых клеток и т. д. Б. я. лежат в основе процессов всасывания в жел.-киш. тракте, в основе восприятия вкуса, запаха (см. Обоняние), в основе деятельности всех анализаторов и т. д. Нет физиологич. процесса в живом организме, к-рый в той или иной форме не был бы связан с Б. я.
Но что же такое собственно Б. я., откуда они берутся, каково их участие в процессах жизнедеятельности?
Для облегчения понимания сущности Б. я. любой живой организм можно представить в виде сложной смеси жидкостей и различных химич. соединений. Многие из этих соединений (и поступающие в организм в виде пищи, и выделенные из него в процессе обмена веществ, и промежуточные вещества, образующиеся при обмене веществ) находятся в виде положительно или отрицательно заряженньгх частиц — ионов.
Перераспределение этих ионов и их транспорт, постоянно имеющие место в процессе Жизнедеятельности,— вот причина возникновения Б., я.
Эту разность потенциалов называют потенциалом покоя, или мембранными потенциалом. Наличие его обусловлено неравномерным распределением ионов (в первую очередь ионов натрия и калия) между внутренним содержимом клетгки (ее цитоплазмой) и окружающей клетку средой.
Величина мембранного потенциала различна: для нервной клетки она составляет 60—80 милливольт (.мв), для поперечнополосатых мышечных волокон — 80—90 мв, для воло- кон сердечной мышцы — 90—95 мв, причем для каждого типа клетки в покое величина потенциала строго определенная и отражает интенсивность обменных процессов, протекающих в этой клетке.
В возбужденной клетке регистрируется еще один вид потенциала — так наз. потенциал действия, к-рый, в отличие от потенциала покоя, передвигается в форме волны возбуждения по поверхности клетки со скоростью до нескольких десятков метров в секунду. В каждом возбужденном участке потенциал приобретает обратный знак. Возникновение потенциала действия связано с избирательным увеличением проницаемости клеточной мембраны для ионов натрия.
Существуют и другие виды потенциалов, в частности так наз. потенциал повреждения, или демаркационный потенциал. Этот вид электрич. активности регистрируется между поврежденным и интактным (неповрежденным) участками ткани.
Можно предположить, что его возникновение как бы стимулирует восстановительные (регенерационные) резервы клетки (ткани).
Возбудимые ткани
Эти вещества мы называем медиаторами, а в том случае, когда они передаются в пределах организма в целом на большие расстояния, их традиционно называют гормонами. Но такой способ общения не обеспечивает возможность быстрой передачи информации (напр., при необходимости отдернуть руки при внезапном прикосновенли к огню).
Поэтому природой был выработан другой, более совершенный способ сигнализации и передачи информации — с помощью электрич. импульсов, возникающих в клетке. Особенно отчетливо выражен этот способ в деятельности центральной нервной системы высших животных и человека.
Живой организм является не только генератором биопотенциалов, но и проводником электрич. тока, причем изменение степени электропроводности живых тканей в зависимости от их жизнедеятельности может служить показателем жизнеспособности (состояния) клеток или тканей.
Особой формой Б. я. служит так низ, электрокинетический потенциал, возникающий, напр., при движении крови по кровеносным сосудам. В этом случае появляется разность потенциалов между стенкой сосуда и движущейся кровью. Величина этого потенциала изменяется при некрых патологич. состояниях, что может быть использовано в диагностич. целях.
Возникновением различных форм биоэлектрич. активности сопровождается любой акт жизнедеятельности (мышечное сокращение, работа головного мозга, деятельность сердца и т. д.).
Компоненты потенциала действия и механизм их возникновения
При трансмембранном способе регистрации возникает потенциал действия, состоящий из 3-х основных компонентов:
1 компонент: местный (локальный ответ);
2 компонент: пик (спайк);
3 компонент: следовые потенциалы (отрицательный и положительный).
Спайк (пик) — самая постоянная часть.
Он состоит из восходящего колена (фаза деполяризации) и нисходящего колена (реполяризация). Остальные компоненты изменчивы и могут отсутствовать.
Местный (локальный) ответ возникает и продолжается до тех пор, пока раздражитель не достигнет пороговой величины. Если раздражитель (его сила) меньше 50-75 % пороговой величины проницаемость мембраны изменяется незначительно и равновесно для всех ионов (неспецифично).
После достижения силы раздражителя 50-75 % начинает преобладать натриевая проницаемость, т. к. натриевые каналы освобождаются от ионов Са2+. Происходит снижение мембранного потенциала при достижении пороговой величины разность потенциалов достигает критического уровня деполяризации.
Критический уровень деполяризации (Ек) — это та разность потенциалов, которая должна быть достигнута, чтобы местные изменения перешли в пик потенциала действия.
Ек — пороговая величина, при которой местные изменения переходят в распространенные. Ек величина практически постоянная и равна — 40 — -50 мВ. Разность между мембранным потенциалом и пороговой величиной характеризует порог раздражения и отражает возбудимость ткани.
Пик потенциала действия состоит из следующих фаз.
Фаза деполяризации возникает в результате лавинообразного движения Nа+ внутрь клетки.
Этому способствуют две причины: открываются потенциалзависимые Nа+-каналы. В этом случае происходит деполяризация по типу процесса с положительной обратной связью (самоподкрепляющийся процесс).
Освобождение натриевых каналов от Са2+.
Заряд клеточной мембраны сначала снижается до 0 (это собственно деполяризация), а затем меняется на противоположный (инверсия или овершут).
Для характеристики фазы деполяризации вводится понятие реверсии — это та разность потенциалов, на которую потенциал действия превышает потенциал покоя.
Р=(потенциал действия) — (мембранный потенциал) 20-30 = 50-60 мВ.
Р (реверсия) — это то количество мВ на которое произошла перезарядка мембраны.
Фаза деполяризации продолжается до достижения электрохимического равновесия по Nа+. Затем наступает следующая фаза. Амплитуда потенциала действия не зависит от силы раздражителя. Она зависит от концентрации Nа+ (как снаружи, так и внутри клетки), от количества натриевых каналов, особенностей натриевой проницаемости.
Фаза реполяризации характеризуется:
- снижением проницаемости клеточной мембраны для Nа+ (Nа-инактивация).
Натрий накапливается на наружной поверхности клеточной мембраны;
- возрастает проницаемость мембраны для К+, в результате повышается выход К+ из клетки с увеличением положительного заряда на мембране;
- изменение активности Nа+-К+ насоса.
Реполяризация — это процесс восстановления заряда мембраны.
Но полного восстановления нет, т. к. возникают следовые потенциалы.
Следовые потенциалы делятся на:
- Отрицательный следовой потенциал — замедление реполяризации клеточной мембраны. Это результат проникновения внутрь клетки какого-то количества Nа+, таким образом, отрицательный следовой потенциал — это следовая деполяризация.
- Положительный следовой потенциал — увеличение разности потенциалов.Это результат повышенного выхода ионов К+ из клетки. Положительный следовой потенциал — это следовая гиперполяризация. Как только калиевая проницаемость возвращается к исходному уровню — регистрируется мембранный потенциал.
Электрические явления в живых тканях
Впервые электрические явления в живых тканях были обнаружены Л. Гальвани (1786 г). Во время своих экспериментов он заметил, что лапки лягушки, подвешенные на медном крючке к железным перилам балкона, при каждом соприкосновении с перилами сокращались.
Однако А.Вольта(1792г) доказал, то в этом эксперименте разность потенциалов возникает между двумя разнородными металлами в момент замыкания цепи влажным проводником, роль которого выполняют ткани лягушки.
В настоящее время установлено, что между наружной и внутренней поверхностью клеточной мембраны постоянно существует разность потенциалов вследствие неодинаковой концентрации ионов внутри и вне клетки. В условиях относительного физиологического покоя внутренняя поверхность мембраны несёт отрицательный заряд по отношению к наружной.
Разность потенциалов между наружной и внутренней поверхностью мембраны в условиях покоя получила название -мембранный потенциал покоя. Его можно зарегистрировать при внутриклеточном способе отведения биопотенциалов на экране катодного осциллографа, используя микроэлектроды. При возбуждении изменяется проницаемость мембраны по отношению к иона.
Возникает перемещение ионов, что приводит к перезарядке мембраны и появлению потенциалов действия
Потенциал действия — это быстрое колебание мембранного потенциала, сопровождающееся перезарядкой клеточной мембраны, возникающее при возбуждении.
При регистрации с помощью катодного осциллографа потенциала действия нерва можно обнаружить три его составные части:
- местный потенциал (предспайковый)
- высоковольтный пиковый потенциал (спайк)
- следовые колебания:
- отрицательный следовый потенциал;
- положительный следовый потенциал.
Потенциал действия распространяется по возбудимой ткани при помощи малых круговых токов.
Спайк потенциала действия на всём протяжении его распространения сохраняет одинаковую амплитуду. За счёт спайка осуществляется передача информации по нервному или мышечному волокну.
Следовые потенциалы являются результатом последующих восстановительных процессов в нервном и мышечном волокне
Обратите внимание на то, что в возбудимых тканях животного организма при внеклеточном отведении можно наблюдать два вида биоэлектрических явлений: токи покоя и токи действия.
Ток покоя всегда однофазный, имеет значительную длительность.
Биоэлектрические явления в живых тканях
Ток, сопровождающий деятельное состояние органов и тканей, возникающий при возбуждении, носит название – ток действия. Он обычно двухфазный, малой продолжительности.
Двухфазность тока действия, связана с распространением процесса возбуждения по возбудимым тканям. Ток действия может быть однофазным, если один из отводящих электродов приложен к поврежденному участку возбудимой ткани.
Классический опыт, доказывающий возникновение токов действия — опыт вторичного сокращения (К.Маттеуччи).
Для этой цели приготовляют два нервно-мышечных препарата. Нерв второго препарата помещают продольно на мышцу первого. При раздражении нерва первого препарата сокращается не только его мышца, но и мышца второго нервно-мышечного препарата.
Это явление объясняется тем, что при раздражении нерва в мышце возникает биоток, который возбуждает нерв второго препарата.
Читайте также: