
Триптофановый оперон кратко и понятно
Обновлено: 04.07.2024
Этот регуляторный механизм был описан Ф. Джейкобом и Дж. Моно в 1961 г., что принесло им Нобелевскую премию по физиологии и медицине в 1965 г. Эти исследователи предложили и продемонстрировали действие оперонов через гены, которые кодируют ферменты, необходимые для кишечная палочка для употребления лактозы.
Опероны отвечают за координацию синтеза белка в соответствии с потребностями каждой клетки, то есть они экспрессируются для генерации белков только в то время и в том месте, где они необходимы.
Гены, содержащиеся в оперонах, обычно являются структурными генами, что означает, что они кодируют важные ферменты, которые непосредственно участвуют в метаболических путях внутри клетки. Это может быть синтез аминокислот, энергии в виде АТФ, углеводов и т. Д.
Опероны также обычно встречаются у эукариотических организмов, однако, в отличие от прокариотических организмов, у эукариот область оперона не транскрибируется как единая молекула информационной РНК.
Открытие
Такой ответ клеток на субстраты наблюдается у бактерий в течение многих лет. Однако исследователи задались вопросом, как именно клетка определяет, какой фермент синтезировать для метаболизма этого субстрата.
Джейкоб и Монод наблюдали, что бактериальные клетки в присутствии галактозоподобных углеводов производили в 100 раз больше β-галактозидазы, чем в нормальных условиях. Этот фермент отвечает за расщепление β-галактозидов, так что клетка метаболически их использует.
Точно так же Джейкоб и Моно обнаружили генетическую область с тремя генами, которые контролировались скоординированным образом: ген Z, кодирующий фермент β-галактозидазу; ген Y, кодирующий фермент лактозопермеазу (транспорт галактозидов); и ген A, который кодирует фермент трансацетилазу, который также важен для ассимиляции галактозидов.
Модель оперона
Модель оперона была впервые точно описана в 1965 году Джейкобом и Моно для объяснения регуляции генов, которые транскрибируются и транслируются для ферментов, необходимых для кишечная палочка чтобы усваивать лактозу как источник энергии.
Эти исследователи предположили, что транскрипты гена или набора генов, которые расположены последовательно, регулируются двумя элементами: 1) регуляторным геном или геном-репрессором, 2) и геном-оператором или последовательностью оператора.
Транскрипция подавляется, когда репрессор связан с геном оператора. Таким образом, генетическая экспрессия генов, кодирующих ферменты, необходимые для ассимиляции лактозы, не экспрессируется, и, следовательно, он не может метаболизировать указанный дисахарид.
В настоящее время известно, что связывание репрессора с оператором стерическими механизмами предотвращает связывание РНК-полимеразы с сайтом промотора, так что она начинает транскрибировать гены.
Ген оператора находится между генетической областью последовательности, известной как промотор, и структурными генами. Однако Якоб и Моно не идентифицировали этот регион в свое время.
Классификация оперонов
Опероны классифицируются только по трем различным категориям, которые зависят от того, как они регулируются, то есть некоторые экспрессируются непрерывно (конститутивно), другим нужна определенная молекула или фактор для активации (индуцибельный), а другие экспрессируются непрерывно до тех пор, пока что индуктор выражен (подавляемый).
Три типа оперонов:
Индуцибельный оперон
Опероны этого типа регулируются молекулами окружающей среды, такими как аминокислоты, сахара, метаболиты и т. Д. Эти молекулы известны как индукторы. Если молекула, которая действует как индуктор, не обнаружена, гены оперона не транскрибируются активно.
В индуцибельных оперонах свободный репрессор связывается с оператором и предотвращает транскрипцию генов, обнаруженных в опероне. Когда индуктор связывается с репрессором, образуется комплекс, который не может связываться с репрессором, и, таким образом, гены оперона транслируются.
Репрессируемый оперон
Эти опероны зависят от конкретных молекул: аминокислот, сахаров, кофакторов или факторов транскрипции, среди прочего. Они известны как корепрессоры и действуют совершенно противоположно индукторам.
Только когда корепрессор связывается с репрессором, транскрипция останавливается, и, таким образом, транскрипция генов, содержащихся в опероне, не происходит. Тогда транскрипция репрессируемого оперона останавливается только в присутствии корепрессора.
Учредительный оперон
Эти типы оперонов не регулируются. Они постоянно активно транскрибируются, и в случае какой-либо мутации, затрагивающей последовательность этих генов, это может повлиять на жизнь содержащих их клеток и, в целом, вызвать запрограммированную гибель клеток.
Примеры
Самый ранний и наиболее известный пример функции оперона - это оперон. лак (лактоза). Эта система отвечает за преобразование лактозы, дисахарида, в моносахариды глюкозу и галактозу. В этом процессе действуют три фермента:
- β-галактозидаза, отвечающая за превращение лактозы в глюкозу и галактозу.
- Лактозопермеаза, ответственная за транспортировку лактозы из внеклеточной среды во внутреннюю часть клетки и
- Трансцетилаза, которая принадлежит системе, но имеет неизвестную функцию
Оперон trp (триптофан) из кишечная палочка контролирует синтез триптофана, имея в качестве предшественника хорисминовую кислоту. В этом опероне находятся гены пяти белков, которые используются для производства трех ферментов:
- Первый фермент, кодируемый генами E и D, катализирует первые две реакции пути триптофана и известен как антранилатсинтетаза.
- Второй фермент - это глицеринфосфат, который катализирует последующие этапы образования антранилатсинтетазы.
- Третий и последний фермент - триптофансинтетаза, ответственная за производство триптофана из индол-глицеринфосфата и серина (этот фермент является продуктом генов B и A)
Еще один регуляторный элемент был открыт Чарльзом Янофски (Charles Yanofsky) и его коллегами при изучении триптофано- вого оперона E. coli. Транскрибируемая с этого оперона мРНК длиной 7kb кодирует пять ферментов, превращающих хоризмат в триптофан (разд. 21.9). Эти пять белков синтезируются при трансляции полици- стронной trp-мРНК последовательно, координированно и в эквимолярных количествах. Трансляция начинается раньше, чем заканчивается транскрипция. trp-мРНК синтезируется примерно за 4 мин и затем быстро разрушается. Короткое время жизни trp-мРНК, составляющее всего около 3 мин, позволяет бактериям быстро реагировать на изменяющуюся потребность в триптофане. Е. coli может менять скорость образования ферментов биосинтеза триптофана более чем в 700 раз.
Как осуществляется эта регуляция? Первый уровень регуляции достигается путем взаимодействия специфического репрессора с trp-операторным участком ДНК. trp- репрессор - белок с массой 58 кДа, кодируемый геном trpR, удаленным от trp-оперо- на на довольно большое расстояние. Комплекс этого репрессора и триптофана прочно связывается с оператором, тогда как сам по себе репрессор с ним не связывается. Другими словами, триптофан является корепрессором. Мишень, на которую действует комплекс триптофана с репрессором, участок ДНК, обладающий симметрией второго порядка (рис. 28.11); и в этом случае симметрия играет важную роль во взаимодействии белка с ДНК. Этот операторный участок перекрывается с промоторным участком инициирования транскрипции. Таким образом, связывание trp-penpeccopa с оператором препятствует связыванию РНК-полимеразы с trp-промотором, и гены trp не транскрибируются.
Рис. 28.10. Схема trр-оперона. Показаны контрольные участки - промотор (р), оператор (о) и аттенюатор (а), а также гены, кодирующие лидерную последовательность (К), и ферменты пути биосинтеза триптофана (Е, Б, С, В и А)

Рис. 28.11. Последовательность оснований trp-оператора. Ось симметрии второго порядка обозначена зеленым. Пара оснований, отмеченная +1,- начало транскрибируемой части оперона

В течение некоторого времени считалось, что ингибирование конечным продуктом каталитической активности первого ферментативного комплекса в пути биосинтеза триптофана (разд. 21.11) и система ингибирования транскрипции с участием репрессора и оператора - основные регуляторные системы биосинтеза триптофана. Эта точка зрения была неожиданно опровергнута, когда было обнаружено, что у некоторых мутантов с делециями между оператором и геном первого фермента (trpE) в опероне происходит повышенное образование trp-мРНК. К тому же анализ 5'-концевой последовательности trp-мРНК показал, что там имеется лидерная последовательность длиной 162 нуклеотида, расположенная перед инициирующим кодоном trpE. Затем оказалось, что делеции, повышающие содержание trp-мРНК, картируются в этой лидерной области, примерно в 30-60 нуклеотидах от начала гена trpE. Следующее поразительное наблюдение состояло в том, что при высокой концентрации триптофана образовывался транскрипт, содержащий всего 130 нуклеотидов лидерной последовательности; при нехватке же триптофана синтезировалась trp-мРНК длиной 7000 нуклеотидов, включающая полную лидерную последовательность. Отсюда Янофски сделал вывод, что транскрипция trp-оперона должна регулироваться участком контролируемой терминации, называемым аттенюатором. Он локализован между оператором и геном первого фермента пути биосинтеза триптофана. Этот участок терминации, регулируемый физиологическими условиями, подобен участкам терминации в конце других оперонов (разд. 25.15); он содержит GC-богатую последовательность и следом за ней АТ-богатый участок. Каждый из этих участков аттенюатора обладает симметрией второго порядка (рис. 28.12). Кроме того, терминированный лидерный транскрипт заканчивается несколькими U подряд.
Рис. 28.12. Последовательность оснований аттенюаторного участка Связанные симметрией второго порядка пары оснований GC-богатой области показаны синим цветом; такие же пары AT-богатой области показаны желтым цветом

Аттенюаторный участок дополняет оператор в регуляции транскрипции trp-генов. При изобилии триптофана инициация транскрипции блокируется в результате связывания комплекса триптофан-репрессор с оператором. По мере снижения концентрации триптофана в клетке репрессия снижается и начинается транскрипция. Однако некоторые молекулы РНК-полимеразы покидают матрицу, дойдя до аттенюатора, тогда как другие продолжают синтезировать полную trp-матрицу. По мере исчерпания триптофана увеличивается доля молекул РНК- полимеразы, проходящих через аттенюаторный участок.
28.9. Аттенюация опосредуется трансляцией лидерной мРНК
Каким образом аттенюаторный участок trp- оперона улавливает концентрацию триптофана в клетке? В решении этого вопроса особенно важную роль сыграли данные о том, что часть лидерной мРНК транслируется. Весьма существенно, что в 14-членном лидерном полипептиде (рис. 28.13) имеются остатки триптофана в положениях 10 и 11. Когда триптофан находится в избытке, синтезируется полный лидерный пептид. Но если триптофана не хватает, рибосома задерживается на двух расположенных тандемом кодонах UGG, поскольку в этом случае оказывается недостаточным содержание триптофанил-тРНК. Застрявшая рибосома каким-то образом меняет структуру мРНК, так что РНК-полимераза транскрибирует оперон за пределами аттенюаторного участка. Ключевой аспект этого регуляторного механизма состоит в том, что трансляция и транскрипция тесно сопряжены между собой. Рибосомы, транслирующие лидерную trp-мРНК, следуют непосредственно за молекулой РНК-полимеразы, транскрибирующей ДНК-матрицу. Исследования, проведенные в последние годы, показали, что застрявшая рибосома изменяет вторичную структуру мРНК: конформация, при которой основания спариваются, благоприятствуя тем самым терминированию транскрипции, изменяется таким образом, что РНК-полимераза проскакивает аттенюатор (рис. 28.14). Мы начинаем понимать, что молекулы нуклеиновых кислот, как и белковые молекулы, могут принимать различные конформации и что изменения конформации регулируются и имеют далеко идущие физиологические последствия.
Рис. 28.13. Последовательность аминокислот в лидерном пептиде trp и последовательность оснований в соответствующей лидерной мРНК

Рис. 28.14. Схематическое изображение аттенюации trp-оперона E. coli. Когда триптофан имеется в избытке (А), лидерный участок (обозначен цифрой 1) trp-мРНК полностью транслируется. Участок 2 взаимодействует с рибосомой, что позволяет основаниям участков 5 и 4 спариваться. Эта спаренная область каким-то образом сигнализирует РНК-полимеразе о том, что следует закончить транскрипцию. Если же триптофана не хватает (Б), участки 3 и 4 не взаимодействуют, так как рибосома застревает на trp-кодонах участка 1. Участок 2. взаимодействует с участком 3. вместо того, чтобы входить в рибосому, и в результате участки 3 и 4 не могут спариваться. Вследствие этого транскрипция продолжается

28.10. Аттенюаторный участок гистидинового оперона содержит семь гистидиновых кодонов подряд
В настоящее время известны еще два оперона биосинтеза аминокислот у E. coli, содержащих аттенюаторные участки. Фенилала- ниновый оперон и гистидиновый оперон, подобно триптофановому оперону, содержат регулируемые участки терминации перед первым геном, кодирующим фермент. И в этих случаях лидерная область перед участком терминации транслируется. Удивительна последовательность аминокислот в лидерном пептиде фенилаланинового оперона: 7 из 15 остатков - фенилаланины (рис. 28.15). Еще поразительнее лидерный пептид гистидинового оперона: он содержит семь остатков гистидина подряд. Очевидно, что эти лидерные мРНК предназначены, чтобы улавливать концентрации фенилаланина и гистидина. Если соответствующих аминоацилированных тРНК не хватает, трансляция лидера останавливается. Как уже обсуждалось выше на примере trр-оперона, считается, что застрявшая рибосома таким образом изменяет конформацию мРНК, что у нее происходит спаривание оснований. Это дает возможность РНК-полимеразе проскакивать аттенюаторный участок 1 . Присутствие семи последовательно расположенных кодонов гистидина в лидерной мРНК гистидинового оперона существенно увеличивает чувствительность этого детектора. Действительно, падение концентрации гистидил-тРНК на 15% вызывает троекратное увеличение числа молекул мРНК, транскрибируемых с этого оперона.
1 Автор противоречит собственному утверждению, что застрявшая рибосома разворачивает мРНК (см. предыдущий разд.); рибосома действительно способствует именно разворачиванию мРНК. Прим. перев.
Рис. 28.15. Последовательность аминокислот в лидерном пептиде и последовательность оснований соответствующего участка мРНК фенилаланинового (А) и гистидинового (Б) оперонов


Рис. 28.16. Электронная микрофотография фагов
Триптофа́новый оперо́н — оперон, содержащий гены ферментов, задействованных в биосинтезе аминокислоты триптофан. Триптофановый оперон имеется у многих бактерий, впервые был описан у Escherichia coli. Триптофановый оперон является важной экспериментальной моделью для изучения регуляции экспрессии генов.
Триптофановый оперон был описан в 1953 году Жаком Моно и сотрудниками. Он стал первым опероном, для которого была показана регуляция посредством репрессии. В то время как лактозный оперон активируется веществом, на утилизацию которого он направлен (лактозой), триптофановый оперон подавляется триптофаном — соединением, за биосинтез которого ответственен данный оперон. Он содержит 5 структурных генов (цистронов): trpE, trpD, trpC, а также trpB и trpA, кодирующие субъединицы триптофансинтазы [en] . На значительном расстоянии от оперона находится ген trpR, кодирующий белок, подавляющий экспрессию триптофанового оперона. Продукт этого гена в присутствии триптофана связывается с оператором и блокирует транскрипцию оперона. В отличие от lac-оперона, в состав trp-оперона входит особая последовательность — аттенюатор [en] , необходимая для тонкой регуляции транскрипции оперона.
Содержание
Регуляция
Регуляция триптофаного оперона регулируется двумя способами: с помощью белка-репрессора (репрессия), а также с помощью особой последовательности — аттенюатора. При этом в каждом из этих случаев регуляция осуществляется по принципу отрицательной обратной связи.
Репрессия

Белок-репрессор (триптофановый репрессор) имеет молекулярную массу 58 кДа, кодируется геном trpR, расположенным на значительном расстоянии от самого оперона. Ген trpR непрерывно экспрессируется на невысоком уровне, образуя мономеры, которые затем объединяются в тетрамеры. В отсутствие триптофана эти тетрамеры неактивны и распадаются в нуклеоплазме. Однако если концентрация триптофана в клетке высока, то тетрамеры связываются с триптофаном. При этом происходит изменение конформации репрессора, позволяющее ему связаться с оператором. В данном случае существенно, что в триптофановом опероне нуклеотидные последовательности оператора и промотора перекрываются, так что присоединение комплекса L-триптофан•белок-репрессор автоматически блокирует связывание РНК-полимеразы с промотором. Таким образом, транскрипция триптофанового оперона блокируется [1] .
Аттенюация
Аттенюация является вторым механизмом регуляции trp-оперона. Этот способ регуляции возможен потому, что у прокариот, лишённых ядра, процессы транскрипции и трансляции не разделены во времени и пространстве, как у эукариот, и идут одновременно: пока РНК-полимераза синтезирует мРНК, синтезированный участок этой мРНК транслируется рибосомой. В связи с этим процесс трансляции может оказывать непосредственное влияние на транскрипцию оперона.
Сразу после оператора в триптофановом опероне располагается последовательность длиной 162 п. н. [2] , получившая название лидерной последовательности. Она кодирует так называемый лидерный пептид, который получил такое название, поскольку с полицистронной мРНК триптофанового оперона этот пептид синтезируется первым. В состав лидерной последовательности входит особая аттенюаторная последовательность (аттенюатор), которая, влияя на вторичную структуру синтезируемой мРНК, способна вызывать преждевременную терминацию транскрипции. Аналогичная последовательность имеется также у бактерий рода Salmonella [3] .
В trp-опероне Escherichia coli аттенюатор имеет 4 области с обращёнными повторами [en] . Транскрипция аттенюатора приводит к образованию шпилек в мРНК. Возможны 3 варианта шпилек, а именно между последовательностями: 1—2, 2—3, 3—4. При этом образование шпильки 1—2 блокирует образование шпильки 2—3, а образование шпильки 2—3, в свою очередь, препятствует образованию шпильки 3—4. Только шпилька 3—4 является терминаторной, то есть при её образовании РНК-полимераза с высокой вероятностью диссоциирует от ДНК, и транскрипция прерывается.
Схожий механизм аттенюации имеет место при синтезе других аминокислот: гистидина, фенилаланина и треонина [4] . В аттенюаторе гистидинового оперона Escherichia coli имеется 7 гистидиновых кодонов, фенилаланинового — 7 фенилаланиновых кодонов [5] .
Триптофановый оперон Bacillus subtilis
У Bacillus subtilis также имеется триптофановый оперон, транскрипция которого контролируется с помощью аттенюации, однако механизм его регуляции несколько отличается от такового у Escherichia coli. Шпильки могут образовываться в районах А—В и C—D аттенюатора, но лишь последняя вызывает терминацию транскрипции. В отсутствие триптофана образуется шпилька А—В. Так как области В и С частично перекрываются, образование такой шпильки препятствует образованию шпильки С—D, следовательно, транскрипция оперона идёт полностью. Ключевыми отличиями триптофанового оперона Bacillus subtilis от такового у Escherichia coli являются, во-первых, наличие 11 повторяющихся кодонов в лидерной мРНК ( GAG или UAG ), а также наличие особого связывающегося с РНК белка, называемого TRAP (от англ. trp RNA-binding Attenuation Protein ). При высокой концентрации триптофана TRAP связывается с вышеуказанными повторяющимися последовательностями. Так как GAG / UAG -повторы охватывают всю область А, а также частично область В, то шпилька А—В не может образоваться. Это позволяет образоваться шпильке С—D, которая, как говорилось выше, является терминаторной. Таким образом, при наличии триптофана транскрипция trp-оперона блокируется [6] .
См. также
Напишите отзыв о статье "Триптофановый оперон"
Примечания
- ↑Коничев, Севастьянова, 2012, с. 257—258.
- ↑Dale, Park, 2004, с. 88.
- ↑ 123Коничев, Севастьянова, 2012, с. 260.
- ↑Daniel J, Saint-Girons I. Attenuation in the threonine operon: effects of amino acids present in the presumed leader peptide in addition to threonine and isoleucine. // Mol Gen Genet.. — 1982. — Т. 188 , № 2 . — С. 225—227 .
- ↑Dale, Park, 2004, с. 89.
- ↑Dale, Park, 2004, с. 91—92.
Литература
Отрывок, характеризующий Триптофановый оперон
– Eh bien, Rapp, croyez vous, que nous ferons do bonnes affaires aujourd'hui? [Ну, Рапп, как вы думаете: хороши ли будут нынче наши дела?] – обратился он к нему.
– Sans aucun doute, Sire, [Без всякого сомнения, государь,] – отвечал Рапп.
Наполеон посмотрел на него.
– Vous rappelez vous, Sire, ce que vous m'avez fait l'honneur de dire a Smolensk, – сказал Рапп, – le vin est tire, il faut le boire. [Вы помните ли, сударь, те слова, которые вы изволили сказать мне в Смоленске, вино откупорено, надо его пить.]
Наполеон нахмурился и долго молча сидел, опустив голову на руку.
– Cette pauvre armee, – сказал он вдруг, – elle a bien diminue depuis Smolensk. La fortune est une franche courtisane, Rapp; je le disais toujours, et je commence a l'eprouver. Mais la garde, Rapp, la garde est intacte? [Бедная армия! она очень уменьшилась от Смоленска. Фортуна настоящая распутница, Рапп. Я всегда это говорил и начинаю испытывать. Но гвардия, Рапп, гвардия цела?] – вопросительно сказал он.
– Oui, Sire, [Да, государь.] – отвечал Рапп.
Наполеон взял пастильку, положил ее в рот и посмотрел на часы. Спать ему не хотелось, до утра было еще далеко; а чтобы убить время, распоряжений никаких нельзя уже было делать, потому что все были сделаны и приводились теперь в исполнение.
– A t on distribue les biscuits et le riz aux regiments de la garde? [Роздали ли сухари и рис гвардейцам?] – строго спросил Наполеон.
– Oui, Sire. [Да, государь.]
– Mais le riz? [Но рис?]
Рапп отвечал, что он передал приказанья государя о рисе, но Наполеон недовольно покачал головой, как будто он не верил, чтобы приказание его было исполнено. Слуга вошел с пуншем. Наполеон велел подать другой стакан Раппу и молча отпивал глотки из своего.
– У меня нет ни вкуса, ни обоняния, – сказал он, принюхиваясь к стакану. – Этот насморк надоел мне. Они толкуют про медицину. Какая медицина, когда они не могут вылечить насморка? Корвизар дал мне эти пастильки, но они ничего не помогают. Что они могут лечить? Лечить нельзя. Notre corps est une machine a vivre. Il est organise pour cela, c'est sa nature; laissez y la vie a son aise, qu'elle s'y defende elle meme: elle fera plus que si vous la paralysiez en l'encombrant de remedes. Notre corps est comme une montre parfaite qui doit aller un certain temps; l'horloger n'a pas la faculte de l'ouvrir, il ne peut la manier qu'a tatons et les yeux bandes. Notre corps est une machine a vivre, voila tout. [Наше тело есть машина для жизни. Оно для этого устроено. Оставьте в нем жизнь в покое, пускай она сама защищается, она больше сделает одна, чем когда вы ей будете мешать лекарствами. Наше тело подобно часам, которые должны идти известное время; часовщик не может открыть их и только ощупью и с завязанными глазами может управлять ими. Наше тело есть машина для жизни. Вот и все.] – И как будто вступив на путь определений, definitions, которые любил Наполеон, он неожиданно сделал новое определение. – Вы знаете ли, Рапп, что такое военное искусство? – спросил он. – Искусство быть сильнее неприятеля в известный момент. Voila tout. [Вот и все.]
Рапп ничего не ответил.
– Demainnous allons avoir affaire a Koutouzoff! [Завтра мы будем иметь дело с Кутузовым!] – сказал Наполеон. – Посмотрим! Помните, в Браунау он командовал армией и ни разу в три недели не сел на лошадь, чтобы осмотреть укрепления. Посмотрим!
Он поглядел на часы. Было еще только четыре часа. Спать не хотелось, пунш был допит, и делать все таки было нечего. Он встал, прошелся взад и вперед, надел теплый сюртук и шляпу и вышел из палатки. Ночь была темная и сырая; чуть слышная сырость падала сверху. Костры не ярко горели вблизи, во французской гвардии, и далеко сквозь дым блестели по русской линии. Везде было тихо, и ясно слышались шорох и топот начавшегося уже движения французских войск для занятия позиции.
Наполеон прошелся перед палаткой, посмотрел на огни, прислушался к топоту и, проходя мимо высокого гвардейца в мохнатой шапке, стоявшего часовым у его палатки и, как черный столб, вытянувшегося при появлении императора, остановился против него.
– С которого года в службе? – спросил он с той привычной аффектацией грубой и ласковой воинственности, с которой он всегда обращался с солдатами. Солдат отвечал ему.
– Ah! un des vieux! [А! из стариков!] Получили рис в полк?
– Получили, ваше величество.
Наполеон кивнул головой и отошел от него.
В половине шестого Наполеон верхом ехал к деревне Шевардину.
Начинало светать, небо расчистило, только одна туча лежала на востоке. Покинутые костры догорали в слабом свете утра.
Вправо раздался густой одинокий пушечный выстрел, пронесся и замер среди общей тишины. Прошло несколько минут. Раздался второй, третий выстрел, заколебался воздух; четвертый, пятый раздались близко и торжественно где то справа.
Еще не отзвучали первые выстрелы, как раздались еще другие, еще и еще, сливаясь и перебивая один другой.
Наполеон подъехал со свитой к Шевардинскому редуту и слез с лошади. Игра началась.
Вернувшись от князя Андрея в Горки, Пьер, приказав берейтору приготовить лошадей и рано утром разбудить его, тотчас же заснул за перегородкой, в уголке, который Борис уступил ему.
Когда Пьер совсем очнулся на другое утро, в избе уже никого не было. Стекла дребезжали в маленьких окнах. Берейтор стоял, расталкивая его.
– Ваше сиятельство, ваше сиятельство, ваше сиятельство… – упорно, не глядя на Пьера и, видимо, потеряв надежду разбудить его, раскачивая его за плечо, приговаривал берейтор.
– Что? Началось? Пора? – заговорил Пьер, проснувшись.
– Изволите слышать пальбу, – сказал берейтор, отставной солдат, – уже все господа повышли, сами светлейшие давно проехали.
Пьер поспешно оделся и выбежал на крыльцо. На дворе было ясно, свежо, росисто и весело. Солнце, только что вырвавшись из за тучи, заслонявшей его, брызнуло до половины переломленными тучей лучами через крыши противоположной улицы, на покрытую росой пыль дороги, на стены домов, на окна забора и на лошадей Пьера, стоявших у избы. Гул пушек яснее слышался на дворе. По улице прорысил адъютант с казаком.
– Пора, граф, пора! – прокричал адъютант.
Приказав вести за собой лошадь, Пьер пошел по улице к кургану, с которого он вчера смотрел на поле сражения. На кургане этом была толпа военных, и слышался французский говор штабных, и виднелась седая голова Кутузова с его белой с красным околышем фуражкой и седым затылком, утонувшим в плечи. Кутузов смотрел в трубу вперед по большой дороге.
Войдя по ступенькам входа на курган, Пьер взглянул впереди себя и замер от восхищенья перед красотою зрелища. Это была та же панорама, которою он любовался вчера с этого кургана; но теперь вся эта местность была покрыта войсками и дымами выстрелов, и косые лучи яркого солнца, поднимавшегося сзади, левее Пьера, кидали на нее в чистом утреннем воздухе пронизывающий с золотым и розовым оттенком свет и темные, длинные тени. Дальние леса, заканчивающие панораму, точно высеченные из какого то драгоценного желто зеленого камня, виднелись своей изогнутой чертой вершин на горизонте, и между ними за Валуевым прорезывалась большая Смоленская дорога, вся покрытая войсками. Ближе блестели золотые поля и перелески. Везде – спереди, справа и слева – виднелись войска. Все это было оживленно, величественно и неожиданно; но то, что более всего поразило Пьера, – это был вид самого поля сражения, Бородина и лощины над Колочею по обеим сторонам ее.
Над Колочею, в Бородине и по обеим сторонам его, особенно влево, там, где в болотистых берегах Во йна впадает в Колочу, стоял тот туман, который тает, расплывается и просвечивает при выходе яркого солнца и волшебно окрашивает и очерчивает все виднеющееся сквозь него. К этому туману присоединялся дым выстрелов, и по этому туману и дыму везде блестели молнии утреннего света – то по воде, то по росе, то по штыкам войск, толпившихся по берегам и в Бородине. Сквозь туман этот виднелась белая церковь, кое где крыши изб Бородина, кое где сплошные массы солдат, кое где зеленые ящики, пушки. И все это двигалось или казалось движущимся, потому что туман и дым тянулись по всему этому пространству. Как в этой местности низов около Бородина, покрытых туманом, так и вне его, выше и особенно левее по всей линии, по лесам, по полям, в низах, на вершинах возвышений, зарождались беспрестанно сами собой, из ничего, пушечные, то одинокие, то гуртовые, то редкие, то частые клубы дымов, которые, распухая, разрастаясь, клубясь, сливаясь, виднелись по всему этому пространству.
Эти дымы выстрелов и, странно сказать, звуки их производили главную красоту зрелища.
Пуфф! – вдруг виднелся круглый, плотный, играющий лиловым, серым и молочно белым цветами дым, и бумм! – раздавался через секунду звук этого дыма.
Что такое оперон в генетике — кто разработал концепцию
Оперон — это функциональная единица наследственной информации, которая содержится в прокариотических клетках (к ним относятся истинные бактерии и археи) и транскрибирует все гены, находящиеся под общим промотором.
Промотор — это последовательность цепочки ДНК, которую РНК-полимераза узнает как стартовую площадку для начала транскрипции.
В состав оперонов входят цистоны, т.е. гены, синтезирующие определенный белок. По количеству этих единиц транскрипции опероны делятся на такие виды, как:
Осторожно! Если преподаватель обнаружит плагиат в работе, не избежать крупных проблем (вплоть до отчисления). Если нет возможности написать самому, закажите тут.
- моноцистонные;
- олигоцистонные;
- полицистонные.
Одним из примеров оперонной организации генома является лактозный оперон. Эта группа генов, контролирующая синтез ферментов, отвечающих за расщепление молочного сахара (лактозы). Изучив данный механизм, французские ученые Ф.Жакоб и Ж.Моно, разработали концепцию оперона, и за это открытие в области биохимии в 1965 году получили Нобелевскую премию.
Структурная организация и основные функции
Строение оперона
Опероны состоят из промоторной области, оператора и структурных генов. В начале и конце оперона находятся регуляторные области: в начале — промотор, в конце — терминатор. Эти элементы может иметь в своем составе и каждый отдельно взятый цистон.
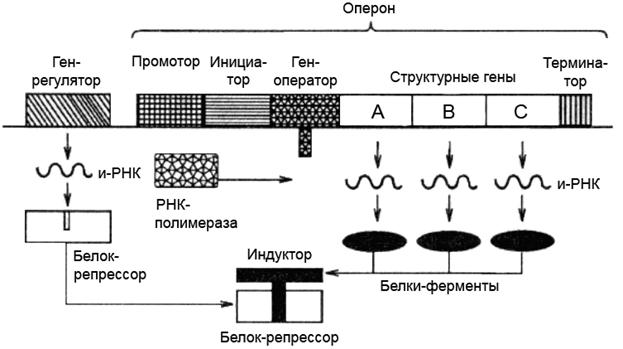
Функции оперона
Опероны участвуют в процессах синтеза полипептидов, необходимых для усвоения определенных питательных веществ. Опероны являются стимулом для того, чтобы в клетке живого организма начали вырабатываться необходимые ферменты. Например, фермент лактаза, за который отвечает оперон Lac, расщепляет молочный сахар. Оперон Trp способствует выработке ферментов, синтезирующих аминокислоту триптофан.
Оперон и транскриптон как единицы транскрипции
Оперон является видом транскриптона, но может присутствовать только у прокариотов. Транскриптоны эукариот содержат, как правило, только один ген. В прокариотических клетках транскрипторные единицы содержат несколько генов и производят полицистронные мРНК. В результате их трансляции рибосомы синтезируют необходимый белок.
Когда происходит включение оперона
Механизм выработки клеткой определенного фермента включается только при попадании вещества в культурную среду. Для включения оперона и начала экспрессии нужного гена необходимо наличие индуктора.
Индуктор (или эффектор) — это вещество-стимулятор экспрессии гена.
Процесс регуляции своевременного включения оперона обозначается понятием индукция. Если в клетке наблюдается недостаток синтезирующего вещества, то индуктор придает белку-регулятору способность присоединяться к оператору, либо препятствует присоединению белка-репрессора. Способ индукции зависит от того, какое вещество находится в среде. Каждый из них по-своему включает нужные участки ДНК.
Когда в клетке наблюдается избыток полипептида, индуктор включает аттенуацию, процесс регуляции оперона путем репрессии стимулятора. Транскрипция гена прекращается, и фермент больше не синтезируется.
Как найти длину оперона
Оперон является участком ДНК, поэтому для вычисления его длины необходимо знать количество нуклеотидов, входящих в его состав. Длина одного нуклеотида составляет 0,34 нм. Нужно умножить их количество на указанное число.
Читайте также: