
Трансформация у бактерий кратко
Обновлено: 04.07.2024
Впервые эта форма изменчивости бактерий была описана в 1935 г. Е. Клинебергер. Она обнаружила и выделила из культуры Streptobacillus moniliformis необычные варианты, которые росли в виде маленьких характерных колоний с врастающей в агар центральной и фестончатой полупрозрачной периферической зонами. В этих колониях обнаруживались самые разнообразные по морфологии структуры: нитевидные, волокнистые, колбасовидные, шаровидные образования и мелкие гранулы размером 0,1 – 0,15 мкм (фильтрующиеся формы бактерий). Поскольку этот феномен был обнаружен в институте имени Листера, то таким необычным вариантам бактерий дали название L-форм, а такую изменчивость бактерий назвали L-трансформацией. Она может быть обратимой и необратимой. В случае если генетический контроль синтеза клеточной стенки сохраняется, L-формы при благоприятных условиях могут возвращаться в исходную бактериальную форму с восстановлением всех основных биологических свойств, включая патогенность. Если же генетический контроль синтеза клеточной стенки нарушен необратимо, L-трансформация приобретает необратимый характер, а такие L-трансформанты по своим морфологическим, культуральным и иным свойствам становятся неотличимыми от микоплазм. L-трансформации могут подвергаться, по-видимому, все бактерии, имеющие клеточную стенку, а все образующиеся L-формы, независимо от вида бактерий, из которого они возникли, обладают следующими общими для них особенностями:
1. Сходство морфологических изменений: образование нитевидных, волокнистых, колбасовидных, шаровидных и гранулярных форм.
3. Постепенное (по мере нарушения синтеза клеточной стенки) превращение из грамположительных в грамотрицательные структуры.
4. Образование стабильных и нестабильных L-форм (в зависимости от степени полноты утраты способности синтезировать клеточную стенку).
5. Изменение антигенных свойств (утрата К– и О-антигенов как следствие нарушения синтеза клеточной стенки).
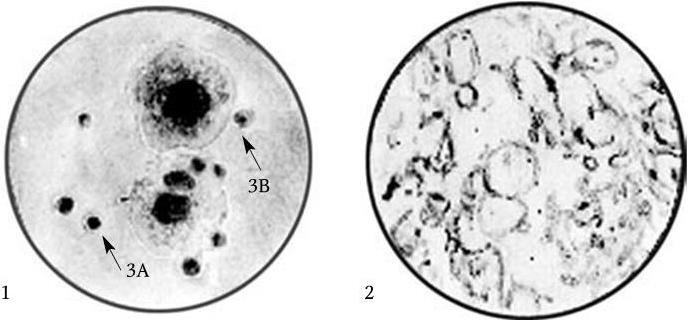
Рис. 8
1 – колонии L-форм типа 3А и 3В; 2 – пузыревидные, грушевидные и субмикроскопические элементы L-форм дифтерийной палочки
6. Снижение вирулентности по сравнению с исходными родительскими формами в связи с утратой различных факторов патогенности (адгезии, инвазии, эндотоксина и т. п.).
7. Способность длительно персистировать (переживать) в организме. Утрата клеточной стенки делает L-формы нечувствительными к различным химиопрепаратам и антителам.
8. Способность при неполной утрате синтеза клеточной стенки возвращаться в исходную бактериальную форму.
L-трансформация происходит как in vitro, так и in vivo (в организме человека и животных). Факторами, индуцирующими ее, являются различные антибиотики, угнетающие биосинтез клеточной стенки (пенициллин, цефалоспорины, циклосерин, ванкомицин и т. п.); ферменты (лизоцим, амидаза, эндопептидаза); антимикробные антитела; высокие концентрации некоторых аминокислот, особенно глицина и фенилаланина.
Исключительное значение L-трансформации патогенных бактерий заключается в том, что она является частой причиной перехода острых форм заболеваний в хронические и их обострений. L-трансформацию надо рассматривать не просто как одно из проявлений изменчивости бактерий, а как своеобразную, присущую всем бактериям форму приспособления к неблагоприятным условиям существования (подобно спорообразованию), которая способствует сохранению вида бактерий в природе. Клеточная стенка и ее синтез чувствительны к действию антител и различных химиопрепаратов. Освобождение от нее не лишает бактерии жизнеспособности, но позволяет переживать действие этих неблагоприятных для них факторов, а по их устранении – возвращаться в свое исходное состояние.
Принимая во внимание особую роль клеточной стенки в жизни бактерий, ей можно дать такое определение. Клеточная стенка – сложный структурный элемент, встречающийся только у эубактерий (кроме микоплазм) и характеризующийся наличием в его составе уникального химического соединения – пептидогликана, наделяющего клетку важными иммунобиологическими свойствами и определяющего ее постоянную форму; нарушение его синтеза приводит к превращению бактерий в L-формы, с помощью которых и обеспечивается, главным образом, длительное персистирование возбудителя в организме – одна из основных причин перехода заболевания из острой в хроническую форму. Соответственно L-трансформация, как и спорообразование, является важнейшей формой приспособления бактерий к неблагоприятным условиям существования.
11. Механизмы передачи генетического материала у бактерий: конъюгация, трансдукция, трансформация.
Рекомбинация между геномами бактерий осуществляется 3-мя механизмами: конъюгацией, трансдукцией и трансформацией
Трансформация – передача генетической информации через выделенную из клетки-донора ДНК.
1928г. Ф Гриффит (опыт с вирулентнотью пневмококков)
По происхождению ДНК может быть плазмидной или хромосомной и нести гены, трансформирующие реципиента. Подобным путем среди бактериальных популяций могут распространять гены, кодирующие факторы вирулентности, однако в обмене генетической информацией трансформация играет незначительную роль.
Трансформирующей активностью обладает только двунитчатая высокоспирализованная ДНК.
В клетку-реципиент проникает только одна нить ДНК, другая – в клеточной мембране подвергается деградации с освобождением энергии, необходимой для проникновения в клетку второй нити ДНК.
Интеграция с хромосомой требует наличия гомологичных участков с трансформирующей ДНК.
Процесс трансформации зависит от компетентности клетки-реципиента и состояния трансформирующей донорской ДНК.
Компетентность клеток бактерий (способность воспринимать трансформирующую ДНК) зависит от присутствия в ЦПМ особых белков, обладающих специфическим аффинитетом к ДНК. Компетентность зависит от фазы роста бактериальной культуры, вида бактерий. Может увеличиваться при обработке культуры раствором хлорида кальция при пониженной температуре.
Трансформация служит хорошим инструментом для картирования хромосом, поскольку трансформированные клетки включают различные фрагменты ДНК. Определение частоты одновременного приобретения двух заданных характеристик (чем ближе расположены гены, тем более вероятно, что они оба включатся в один и тот же участок ДНК) дает информацию о взаиморасположении соответствующих генов в хромосоме.
Трансформация является основным методом генной инженерии, используемым при конструировании рекомбинантных штаммов с заданным геномом.
Трансдукция – передача бактериальной ДНК посредством бактериофага.
1952г. Дж. Ледебург и Н. Циндер (опыты с сальмонеллами)
Так как трансдуцируются лишь небольшие фрагменты ДНК, вероятность рекомбинации, затрагивающей какой-то определенный признак, очень мала: она составляет от 10 -6 до 10 -8 .
· Общая (неспецифическая) трансдукция – перенос бактериофагом фрагмента любой части бактериальной хромосомы. Феномен неспецифической трансдукции может быть использован для картирования бактериальной хромосомы.
● Специфическая трансдукция – фаговая ДНК интегрирует в бактерию с образованием профага. При исключении ДНК фага из бактериальной хромосомы захватывается прилегающий к месту включения фаговой ДНК фрагмент бактериальной хромосомы. Специфическая трансдукция может служить механизмом переноса вирулентных генов среди бактерий при условии, что эти гены локализованы в непосредственной близости от мест интеграции профага.
● Абортивная трансдукция . При абортивной трансдукции внесенный фрагмент ДНК донора не встраивается в хромосому реципиента, а остается в цитоплазме и там самостоятельно функционирует. Впоследствии он передается одной из дочерних клеток (т.е. наследуется однолинейно) и затем теряется в потомстве.
Обязательное условие – трансмиссивная плазмида ( F , R ), обладающая tra -опероном .
Интегративной трансмиссивной плазмидой является F -фактор . Донорские клетки, имеющие встроенный в хромосому F -фактор, называются Hfr -клетками.
Биологическое значение конъюгации хорошо видно на примере распространения резистентности бактерий к антибиотикам. Устойчивость к антибиотикам бактерия может получить в результате мутации, что происходит 1 раз на каждые 106 клеточных делений. Изменившаяся генетическая информация может быстро распространяться среди сходных бактерий посредством конъюгации.
Трансформация — поглощение изолированной ДНК бактерии донора клетками бактерии реципиента. Явление трансформации кратко освещено при изложении доказательств роли ДНК в наследственности. В процессе трансформации принимают участие две бактериальные клетки: донор и реципиент. Трансформирующий агент представляет собой часть молекулы ДНК донора, которая внедряется в геном реципиента, изменяя его фенотип. В процессе трансформации клетки донора и реципиента не соприкасаются друг с другом. Механизм переноса генетического материала заключается в том, что из клеток донора выделяются в окружающую среду молекулы или фрагменты молекул ДНК.
Сначала эта ДНК адсорбируется на оболочке клетки реципиента. Затем через определенные рецепторные участки ее стенки при помощи специальных клеточных белков ДНК втягиваются внутрь клетки. Проникающая донорская ДНК должна быть двух-цепочной. В реципиентной клетке она становится одноцепоч-ной. В ДНК реципиента включается одна из цепей трансформирующего фрагмента. Эта цепь вступает в синапсис с гомологичным участком хромосомы реципиента и встраивается в нее посредством кроссинговера. При этом участок ДНК реципиента замещается фрагментом донора. Молекула ДНК со вставкой трансформирующего участка оказывается гибридной. При следующем удвоении возникают одна нормальная дочерняя молекула ДНК, другая — трансформированная. Установлено, что способность бактерий-реципиентов к трансформации определяется их физиологическим состоянием. Такое физиологическое состояние было названо компетентностью. Состояние компетентности краткосрочно и приурочено к определенному времени клеточного цикла. Было обнаружено, что трансформирующей способностью обладают только крупные молекулы ДНК с молекулярной массой не менее 5-Ю 5 Д. У бактерий трансформация имеет место чаще в пределах одного вида, но наблюдается и между разными близкими видами. Это указывает на то, что у них сохранилась гомологичность некоторых участков ДНК
Изучение процессов рекомбинации у бактерий имеет важное значение для ветеринарного врача, так как ведет к пониманию причин высокой изменчивости бактерий, их способности к приобретению свойств патогенности, устойчивости к лекарственным веществам.
Контрольные вопросы. 1. Каково строение бактерий и вирусов? 2. Как размножаются вирусы и бактгр:ш? 3. Что такое профаг, умеренные фаги и лизоге-ния? 4. Что такое геногип и фенотип у микроорганизмов? 5. Как происходит обмен генетическим материалом у бактерий при конъюгации, трансдукции и трансформации?
Трансформация — поглощение изолированной ДНК бактерии донора клетками бактерии реципиента. Явление трансформации кратко освещено при изложении доказательств роли ДНК в наследственности. В процессе трансформации принимают участие две бактериальные клетки: донор и реципиент. Трансформирующий агент представляет собой часть молекулы ДНК донора, которая внедряется в геном реципиента, изменяя его фенотип. В процессе трансформации клетки донора и реципиента не соприкасаются друг с другом. Механизм переноса генетического материала заключается в том, что из клеток донора выделяются в окружающую среду молекулы или фрагменты молекул ДНК.
Сначала эта ДНК адсорбируется на оболочке клетки реципиента. Затем через определенные рецепторные участки ее стенки при помощи специальных клеточных белков ДНК втягиваются внутрь клетки. Проникающая донорская ДНК должна быть двух-цепочной. В реципиентной клетке она становится одноцепоч-ной. В ДНК реципиента включается одна из цепей трансформирующего фрагмента. Эта цепь вступает в синапсис с гомологичным участком хромосомы реципиента и встраивается в нее посредством кроссинговера. При этом участок ДНК реципиента замещается фрагментом донора. Молекула ДНК со вставкой трансформирующего участка оказывается гибридной. При следующем удвоении возникают одна нормальная дочерняя молекула ДНК, другая — трансформированная. Установлено, что способность бактерий-реципиентов к трансформации определяется их физиологическим состоянием. Такое физиологическое состояние было названо компетентностью. Состояние компетентности краткосрочно и приурочено к определенному времени клеточного цикла. Было обнаружено, что трансформирующей способностью обладают только крупные молекулы ДНК с молекулярной массой не менее 5-Ю 5 Д. У бактерий трансформация имеет место чаще в пределах одного вида, но наблюдается и между разными близкими видами. Это указывает на то, что у них сохранилась гомологичность некоторых участков ДНК

Изучение процессов рекомбинации у бактерий имеет важное значение для ветеринарного врача, так как ведет к пониманию причин высокой изменчивости бактерий, их способности к приобретению свойств патогенности, устойчивости к лекарственным веществам.
Контрольные вопросы. 1. Каково строение бактерий и вирусов? 2. Как размножаются вирусы и бактгр:ш? 3. Что такое профаг, умеренные фаги и лизоге-ния? 4. Что такое геногип и фенотип у микроорганизмов? 5. Как происходит обмен генетическим материалом у бактерий при конъюгации, трансдукции и трансформации?
Изучение бактерий открыло целый ряд явлений, осветивших с новой стороны источники наследственной изменчивости и механизмы наследственной передачи.
Одним из первых успехов в этой области было открытие явления трансформации у бактерий в 1928 г.
Известно несколько штаммов пневмококка Diplococcus pneumoniae: штамм S — с полисахаридной капсулой и гладкими колониями и штамм R — без капсулы и с шероховатыми колониями. Оба эти признака наследственны. Бактериолог Ф. Гриффитс инъецировал мышам вместе с убитым нагреванием штаммом пневмококка, обладающим капсулой (S), штамм живого пневмококка, лишенного капсулы (R). Спустя некоторое время ему удалось выделить из зараженных мышей живых пневмококков, обладающих капсулой. Таким образом, оказалось, что свойство убитого пневмококка — способность образовывать капсулу — перешло к живой бактерии. Поскольку признак наличия капсулы является наследственным, то следовало предположить, что какая-то часть наследственного вещества от бактерий штамма S перешла к клеткам штамма R. Но как это могло произойти, если клетки штамма S были убиты? Можно было предполагать, что в этом случае либо возникла мутация, либо произошла своеобразная гибридизация между живыми и мертвыми бактериями. Первое объяснение было наиболее вероятным, однако вопреки здравому смыслу второе объяснение оказалось ближе к истине.
В 1944 г. О. Эвери с сотрудниками удалось выяснить природу этого загадочного явления. Они взяли те же два штамма — R и S. Перед началом решающих опытов было изучено спонтанное мутирование обеих форм. Оказалось, что гладкая S-форма хотя и очень редко, но спонтанно мутирует в R-форму, а R-форма практически вовсе не мутирует в S-форму, т. е. мутации происходят почти исключительно в одном направлении: S→R. Но если R-форму помещали в экстракт из убитых клеток S-формы, то частота изменений R→S увеличивалась в 10 000 раз. Стало очевидным, что признак одного штамма (S) через какое-то вещество экстракта передавался другому штамму (R), т. е. возникало направленное наследственное изменение. Далее была произведена тщательная очистка — выделение этого вещества из экстракта клеток S-формы. Вещество было названо трансформирующим фактором (ТФ), а само явление — трансформацией.
Трансформирующий фактор по своей биохимической природе представлял собой не что иное, как дезоксирибонуклеиновую кислоту, входящую в состав хромосом. При этом было установлено, что он обладает некоторыми характерными свойствами. Его можно экстрагировать из клеток, очищать, воздействовать на него in vitro химическими и физическими факторами и затем снова вводить в живые клетки и изучать вызываемые им изменения.
Сначала к этим исследованиям отнеслись скептически. Но вскоре многие исследователи поняли, что открыто не только новое явление, но и один из новых методов исследования наследственности. В последующих генетических и биохимических исследованиях было показано, что явление трансформации широко распространено у бактерий. Оно твердо установлено у самых различных видов и родов бактерий: Diplococcus, Staphyloccocus, Hemophilus, Neisseria, Agrobacterium, Rhizobium, Bacillus, Xantomonas.
Схема опыта, демонстрирующего явление трансформации
Активность трансформирующего фактора оказалась чрезвычайно высокой. Так, у Hemophilus трансформация осуществляется в течение 15 мин при концентрации ДНК 0,00015γ (γ = 10 -6 г) в 1 мл среды. С помощью меченого фосфора (Р 32 ) было показано, что не вся ДНК донора включается в геном реципиента, а лишь фрагменты с молекулярным весом около 3·10 5 . В то же время под действием фермента дезоксирибонуклеазы (ДНК-азы), разрушающей ДНК, активность трансформирующего агента падает до нуля.
Трансформации могут подвергаться различные признаки. У пневмококков, например, трансформируется наличие капсулы, специфичность белков, размер и морфология колоний, устойчивость к антибиотикам (пенициллину и стрептомицину), способность к окислению определенных веществ и др.
Как правило, трансформируются отдельные свойства, но иногда одновременно несколько признаков в сцепленном состоянии. Р. Хочкис и Дж. Мармур при помощи ДНК, выделенной из штамма пневмококка, устойчивого к стрептомицину и способного сбраживать маннит, трансформировали оба эти свойства другому штамму пневмококка, не обладавшему ими. Одновременная передача обоих признаков от донора к реципиенту происходила в 50 раз чаще, чем это ожидалось, если бы трансформация по обоим признакам осуществлялась независимо. Дополнительная проверка показала, что действительно в рассматриваемом примере имеет место сцепленная передача обоих признаков.
Как правило, трансформация возможна между различными штаммами одного и того же вида, однако недавно была показана возможность межвидовой трансформации. В этом случае донором трансформирующего фактора были виды Hemophilus parainfluenzae или Н. aegypti, а реципиентом — Н. influenzae. Характерной особенностью межвидовой трансформации оказалась низкая частота ее осуществления в сравнении с внутривидовой.
При изучении действия мутагенов на ДНК, обладающую трансформирующей активностью, обнаружена различная чувствительность к мутагенам отдельных наследственных факторов этого трансформирующего фактора. Так, например, облучение ультрафиолетом значительно чаще инактивирует фактор, определяющий форму капсулы у пневмококков, чем фактор, обусловливающий устойчивость к стрептомицину.
Таким образом, трансформация обеспечивает генетическую рекомбинацию у бактерий. В этом может заключаться ее значение для эволюции бактериальных организмов. Обнаружение трансформации и изучение биохимической природы трансформирующего фактора явились вескими аргументами в пользу генетической роли ДНК как материального носителя наследственной информации.
После открытия явления трансформации у бактерий были сделаны попытки обнаружить это явление у высших животных. Получив экстракты ДНК из определенных тканей организма одного генотипа, их вводили другому в надежде на то, что специфическая ДНК донора вызовет в ДНК половых клеток реципиента направленное наследственное изменение. Хотя в этом плане было сделано несколько интересных попыток, убедительных фактов трансформации у высших организмов пока неизвестно. Впрочем, в принципе осуществление трансформации на соматических клетках животных и человека вполне возможно. Так, показано, что клетки в культуре тканей могут усваивать, включать меченую ДНК из среды. Возможно, метод культуры тканей откроет новые перспективы исследований в этой области.
Читайте также: