
Строение стебля однодольных растений кратко
Обновлено: 04.07.2024
Стебель – это осевой вегетативный орган высших растений, часть побега, которая состоит из междоузлий и узлов, его анатомия зависит от выполняемых функций. В результате эволюции он появился не сразу. Спорофиты древнейших сосудистых растений представляли собой дихотомически ветвящиеся оси без листьев и корней. В ходе эволюционной специализации возникли морфологические и физиологические различия между отдельными частями их тел, приведшие к обособлению органов – корней, стеблей и листьев.
Корни в совокупности образовали корневую систему, закрепляющую растение в почве и всасывающую оттуда воду и минеральные вещества. Система побега включила листья со стеблями, пронизанные проводящими тканями, транспортирующими воду и минеральные вещества к листьям, а конечные продукты фотосинтеза – от них.
Стебель нарастает за счёт верхушечных (апекальных), боковых и вставочных меристем (образовательных тканей). В апекальных меристемах появляются новые стебли и органы. Обычно в сечении стебель имеет цилиндрическую форму. Но встречаются уплощённые, ребристые, крылатые, бороздчатые, трехгранные, четырёхгранные и многогранные их разновидности.

Функции стебля
Стебли бывают травянистыми чаще однолетними и одревесневающими: у деревьев они называются стволами, а у кустарников – стволиками. Этот орган выполняет следующие функции:
- опорную: соединяет между собой корень, листья и другие части растения в единое целое;
- механическую — выносит листья к свету;
- запасающую: хранит продукты обмена – жиры, крахмал и другие органические вещества, у кактусов, баобабов, мхов и других растений он запасает воду;
- проводящую;
- служит для опоры и лазания (вьющиеся растения);
- для вегетативного размножения;
- у многих растений его клетки способны к фотосинтезу.
Классификация стеблей
Стебли группируют по разным признакам. Например, по характеру и направлению роста :
- прямостоячие – имеют прямые стебли (пшеница, кукуруза);
- ползучие (стелящиеся, лежачие) – стелются по земле в разные стороны от корня (лапчатка, земляника, камнеломка). Стебель таких растений с длинными междоузлиями и конечными почками, которые называют усами. У огурцов, тыквы образуются плети – облиственные побеги с короткими междоузлиями, стелящиеся по земле;
- вьющиеся стебли поднимаются вверх, обвиваясь вокруг опоры – вьюнок, хмель, лимонник китайский;
- лазающие – имеют усики или придаточные корни, которые отрастают от стебля и при их помощи цепляются за опоры (горох, плющ). Растения с вьющимися и лазающими стеблями ещё называют лианами;
- у некоторых растений только по листьям можно определить наличие стебля, к которому они прикрепляются. Такие стебли получили название укороченных (одуванчик, подорожник). Перед цветением у некоторых растений он развивается в цветущую стрелку.
По месту расположения :
- надземные;
- подземные. Стебель, как часть побега, присутствует в клубнях картофеля, луковицах (плоский стебель-донце), в корневищах пырея, ландыша, крапивы, ириса и др.
Как устроен стебель?
Стебли содержат четыре типа тканей: покровные, механические, основные, проводящие. Внутреннее строение стебля у травянистых растений и деревьев неодинаково. Например, у трав стебель состоит из основной ткани, в которой в беспорядке расположены многочисленные проводящие сосудисто-волокнистые пучки. Стебли однодольных растений не имеют камбия (образовательной ткани), поэтому они почти не растут в толщину. У древесных двудольных растений (липа, клён) пучки сливаются, образуя три концентрических слоя: древесину, камбий и луб. Центральную часть древесного стебля занимает сердцевина. Она может быть рыхлой, как у бузины, и очень плотной и плохо различимой, как у берёзы, дуба.
Развитие зародышевого стебелька начинается при прорастании семени. Он хорошо заметен между семядолями и первыми листочками. Образуется он из меристемы конуса нарастания, в основании которого вырастают бугорки зачаточных листьев. Под ними появляются протодерма, прокамбий и основная меристема. Протодерма в будущем станет эпидермой (покровной тканью), прокамбий превратится в проводящую ткань, а из основной меристемы сформируется основная ткань. У стебля появится первичное строение.
Стебель: первичное строение
Во всех стеблях молодых растений выделяют 3 анатомо-функциональные части: покров, кору и стелу. Первичная флоэма, первичная ксилема, а также сердцевина (если она имеется) образуют центральный цилиндр, или стелу стебля и корня молодого растения. Существует несколько типов стел, основные из них:
- Протостела – наиболее примитивный тип, состоит из сплошного тяжа проводящих тканей, в которой флоэма либо окружает ксилему, либо рассеяна в ней. Этот тип стелы характерен для псилофитов и других вымерших растений, для корней большинства современных растений, для стеблей плауновидных и некоторых других групп представителей флоры.
- Сифоностела, или трубчатая стела. Отличается наличием центрального столба паренхимной основной ткани (сердцевины), окружённой проводящей тканью. Флоэма формируется только снаружи от ксилемного цилиндра или с обеих сторон от него. Такой тип стелы имеют большинство папоротников.
- Эвстела– проводящий цилиндр представлен системой отдельных тяжей, расположенных вокруг сердцевины. У хвощей, многих голосеменных и покрытосеменных.
В разрезе первичное строение стебля растений класса однодольных имеет следующие слои:
- зачаточная эпидерма (протодерма) становится эпидермой. Эпидерма однослойная, покрытая кутикулой;
- кольцо механической ткани– склеренхимы;
- из прокамбия (первичная меристема) – находящегося на границе листового бугорка и будущей оси побега появляются закрытые (лишённые камбия) проводящие пучки.Другими словами – камбий полностью превращается в проводящие элементы. Замкнутые проводящие пучки беспорядочно разбросаны по всей толщине стебля. Каждый такой пучок состоит из древесины (ксилемы), окружённой лубом (флоэмой) и клетками механической ткани – склеренхимы;
- между протодермойи прокамбием лежит основная меристема, она становится первичной корой стебля. В ней есть только паренхимные клетки, покрытые одревесневшими стенками;
- иногда под эпидермой в молодом стебле развивается слой хлоренхимы – ассимиляционной паренхимной ткани;
- основная меристема – внутри прокамбия – будущая сердцевина. Состоит из живых тонкостенных клеток.

Первичное строение стебля двудольных растений
Молодой стебель подсолнечника снаружи покрыт эпидермой. Под ней лежит первичная кора, а в центре расположен осевой цилиндр.
- Эпидерма имеет типичное строение, с малым количеством устьиц. Она выполняет роль газообмена и транспирации. У подсолнечника на выростах эпидермы расположены жёсткие волоски.
- Первичная кора. Состоит из разных элементов. Сразу под эпидермой лежит механическая ткань (у двудольных – колленхима), клетки которой содержат хлоропласты. Ниже её расположена паренхима из клеток с тонкими стенками. Самый нижний слой коры – эндодерма, или крахмалоносное влагалище.
- Центральный осевой цилиндр. Вокруг сердцевины расположены проводящие пучки первичной ксилемы и первичной флоэмы, между которыми лежат клетки камбия. Проводящие пучки разделены паренхимными сердцевинными лучами. Лучи играют важную роль в горизонтальном передвижении растворов по стеблю. Они соединяют сердцевину с корой. Сердцевина состоит из неспециализированной паренхимы, которая может содержать воздушные полости, идиобласты, запасные вещества.
Таблица 1. Строение стеблей однодольных и двудольных растений
| Признаки | Однодольные растения | Двудольные растения |
| Первичная кора | Состоит из паренхимы и склеренхимы | Состоит из паренхимы и колленхимы |
| Тип проводящих пучков | Закрытые (камбий отсутствует) | Открытые (между первичной флоэмой и первичной ксилемой прослойка камбия) |
| Расположение пучков | Беспорядочное | Упорядоченное по кругу |
| Паренхима | Занимает основную часть центрального осевого цилиндра и первичной коры | Образует часть первичной коры, сердцевину и сердцевинные лучи |

Стебель моховидных
Он может состоять из однородных клеток, но у большинства мхов они дифференцированы. В них можно выделить склеродерму (кору), сверху покрытую несколькими слоями мёртвой эпидермы. Настоящие покровные ткани у мхов отсутствуют, так же как и истинная проводящая система. Устьиц в эпидерме у всех листостебельных мхов нет. У некоторых из них снаружи расположен другой тип покровной ткани – гиалодерма, клетки которой лишены хлоропластов, имеют тонкие стенки и расположены так, что между ними образуется расстояние – просвет, в которое проникает вода. Мхи могут впитывать воду всей поверхностью тела.
Внутренняя часть коры состоит из механической ткани, клетки которой имеют толстые стенки, пронизанные водопроводящими порами. Эти клетки могут набухать, запасая воду. Основная функция внутренней коры – обеспечение прочности стебля.
Ниже лежат клетки основной фотосинтезирующей и запасающей ткани. Они хранят воду, масла, крахмал, оксалат кальция.
Проводящий, или центральный пучок проходит по оси стебля, состоит из вытянутых тонкостенных клеток с косыми перегородками. Реже эти клетки толстостенные. У многих мохообразных проводящего пучка нет, тогда на его месте в центре стебля располагаются клетки основной ткани, выполняющей ту же функцию – запасания и проведения органических и минеральных веществ и воды.

Стебель мха Catoscopium nigritum
Автор: Hermann Schachner
Вторичное строение стебля
Вторичная меристема есть только у двудольных растений, поэтому и вторичное строение стебля характерно только для них. Далее мы будем говорить о строении стебля двудольных покрытосеменных растений.
С момента начала деления камбиальных клеток начинается формирование вторичной структуры стебля. Клетки камбия образуют паренхиму, вторичную флоэму, вторичную ксилему и механические элементы.
Эпидермис – первичная кроющая ткань, характерная главным образом для однолетних органов растений. Только у листьев и стеблей вечнозелёных растений эпидермис сохраняется несколько лет. У большинства же многолетних растений уже к концу вегетационного периода эпидермис заменяется вторичной кроющей тканью – перидермой, образующейся или непосредственно под клетками эпидермиса, или на некотором расстоянии от него из более глубоко лежащих паренхимных клеток. Общая последовательность слоёв вторичного строения стебля дерева от наружного до внутреннего выглядит следующим образом:
- Эпидерма отмирает, но сверху часто бывают заметны её остатки. У многолетних древесных растений снаружи развивается третичная ткань – корка.
- Пробка (феллема) – вторичная покровная ткань. Состоит из мёртвых клеток, заполненных воздухом, коричневыми смолистыми или дубильными веществами. В процессе опробковения на её первичную оболочку откладывается суберин, а со стороны клетки на субериновый слой налегает вторичная целлюлозная оболочка. В стенках клеток пор нет.
- Пробковый камбий – феллоген. Однослойная меристема, состоит из коротких клеток, таблитчатых в поперечном сечении.
- Феллодерма построена из живых клеток, внешне похожих на клетки феллогена. Они обычно содержат запасные вещества, которые используются феллогеном. Феллодерма, феллема и феллоген вместе составляют перидерму – комплекс защитных образований.
- Вторичная кора состоит из колленхимы и основной запасающей ткани
- Вторичная флоэма (вторичная кора, или луб). Состоит из лубяных волокон, ситовидных трубок, клеток-спутниц и мягкой паренхимы. Её функции – проведение продуктов фотосинтеза от листьев (нисходящий ток) и запасание веществ.
- Вторичный камбий – камбиальное кольцо.
- Вторичная ксилема (древесина). Состоит из сосудов, либриформа (механических волокон) и основной паренхимной ткани. Основные функции древесины – проведение воды с растворёнными минеральными веществами (восходящий ток).
- Сердцевина состоит из рыхлой паренхимы, служит для запасания веществ.

На срезе ветки или ствола дерева хорошо заметны следующие слои стебля: кора (в составе которой выделяют пробку и луб), камбий, древесина и сердцевина. Стебель многолетнего древесного растения состоит главным образом из древесины, все другие его части сильно уменьшаются.
На поперечном срезе ветки липы выделяют верхний наружный слой коры – кожицу, которая с возрастом заменяется более толстым слоем – пробкой. В пробковом слое есть чечевички – маленькие бугорки с отверстиями, через которые в растение проникает кислород для дыхания. Кора защищает внутренние слои стебля от испарения влаги, в холодное время года – от промерзания, от проникновения атмосферной пыли и др.
Внутренняя часть коры, примыкающая к камбию, представлена лубом. В нём находятся лубяные волокна и ситовидные трубки. Проводящие и механические элементы древесины и луба расположены вдоль стебля, а в поперечном направлении через древесину и луб проходят сердцевинные лучи, состоящие из рядов живых клеток.
Между древесной (ксилемой) и лубом (флоэмой) залегает особый слой живых клеток образовательной ткани, называемый камбием. Клетки камбия делятся в плоскости, но увеличения камбия в толщину не происходит, потому что из двух дочерних клеток, возникающих при делении, только одна сохраняет способность делиться, тогда как вторая становится элементом постоянной ткани. Если такие клетки расположены к периферии от камбия, они становятся клетками луба. Если же к центру стебля – клетками древесины, которых образуется значительно больше и поэтому слой древесины бывает намного шире слоя луба.
Клетки древесины, образовавшиеся за весну, лето и осень, составляют слой, называемый годичным кольцом прироста. Годичные кольца образуются в результате того, что весной с началом сокодвижения камбий откладывает крупные клетки с тонкими оболочками. Осенью у большинства деревьев новые клетки древесины мелкие с более толстыми оболочками. С глубокой осени и до весны следующего года деятельность камбия прекращается.

Видоизменённые надземные стебли
В отличие от почки – зачаточного побега, колючки, усики, кладодии и туберидии являются изменёнными побегами со своими специфическими функциями.
- колючки – одревесневшие части побега с острой верхушкой. Они нужны растениям для самозащиты от поедания животными. Колючки бывают листового и стеблевого происхождения. Из стебля образуются колючки дикой яблони, крушины, боярышника;
- кладодии – стебли, выполняющие функцию фотосинтеза. Если они внешне похожи на листья, то их называют филлокладиями (у акации);
- усики нужны растениям для прикрепления к опоре. Они также бывают стеблевого и листового происхождения. Стеблевые усики есть у винограда, тыквы;
- ложная луковица, псевдобульба, или туберидий, также носит название надземного или воздушного клубня. Утолщённая околосубстратная часть стебля эпифитных растений (орхидных). В ней растения запасают воду и питательные вещества.
Вам будет интересно
Строение растений очень разнообразно и отличаются даже в пределах одного вида. Древнейшие представители флоры, многие…
Хлопковые, льняные, синтетические — это ткани, из которых люди шьют себе одежду. Она нужна им…
В ботанике листья – это вегетативные органы, части побега сосудистых растений. В норме они развиваются…
Листья – основные фотосинтезирующие органы растений. На развитом побеге они инициируются как листовые примордии в…
Хотя представители флоры могут абсорбировать воду всеми частями тела, основным её источником является почва. Но…
Стебель — осевая, опорная часть побега, образующаяся из зародышевого стебелька, обладающая неограниченным верхушечным ростом и положительным гелиотропизмом. Стебель связывает надземные и подземные органы; удерживает в пространстве всю надземную массу; обеспечивает передвижение восходящего и нисходящего токов веществ; ассимилирует, пока зеленый; запасает питательные вещества; участвует в вегетативном размножении. Стебель метамерен, т. е. состоит из повторяющихся частей — узла с листьями, почками, побегами и междоузлия. В стебле скелетно-проводящая система сложнее, чем в корне. Первичное анатомическое строение стебля появляется в конусе нарастания на уровне зачатков листьев — примордиев, где закладывается прокамбий. После смены прокамбия камбием формируется вторичное анатомическое строение. У сформировавшихся стеблей строение разнообразно, но специфично для конкретных систематических групп (однодольных, двудольных, голосеменных, папоротникообразных) и жизненных форм (травянистых, древесных), своеобразно у определенных экологических групп (ксерофитов, гидро- и гигрофитов).
СТЕБЛИ ОДНОДОЛЬНЫХ РАСТЕНИЙ
Строение стеблей однодольных травянистых растений первичное, пучковое. Характеризуется отсутствием боковых меристем — прокамбия, камбия и феллогена, наличием множества листовых следов — закрытых коллатеральных пучков, внедряющихся из листьев в стебель (рис. 1.31). Они распределены по всему сечению центрального цилиндра, причем, на его периферии пучки мелкие, многочисленные, а в срединной части они крупнее и малочисленнее. Сердцевина или морфологически не выражена, или полая.
Рис. 1.31. Схемы прохождения и расположения проводящих пучков в стебле однодольного растения: А — продольный срез; Б — поперечный срез; 1 — индивидуальные листовые следы; 2 — слившиеся листовые следы

Первичная кора чаще всего развита слабо, но может совсем отсутствовать, состоять из нескольких рядов или участков паренхимы, содержать кольцо или отдельные тяжи склеренхимы, а у водных растений — аэренхиму (рис. 1.32). Расположение тканей у злаков показано на примере выполненного стебля кукурузы (рис. 1.32, А) и полых стеблей типа соломины (рожь, тростник), где проводящие пучки располагаются в 2—3 круга (рис. 1.32, Б, В). Строение выполненных стеблей типа лилейных с развитой, однородной корой показано на примере стебля спаржи (рис. 1.33).
Рис. 1.32. Анатомическое строение стеблей представителей сем. злаки: А — кукурузы; Б — ржи; В — тростника; 1 — эпидерма; 2 — хлоренхима; 3 — склеренхима; 4 — основная паренхима; 5 — проводящие пучки; 6 — флоэма; 7 — сосуды ксилемы; 8 — воздухоносные ходы; 9 — полость

Рис. 1.33. Стебель спаржи: 1 — эпидерма; 2 — хлоренхима первичной коры; 3 — перициклическая склеренхима; 4 — основная паренхима центрального цилиндра; 5 — закрытые коллатеральные пучки: а — ксилема; б — флоэма

КОРНЕВИЩА ОДНОДОЛЬНЫХ
Корневище — подземное видоизменение побега, приспособленное к накоплению питательных веществ и к вегетативному возобновлению растения. Преобладающей тканью корневищ является запасающая паренхима (рис. 1.34). Анатомическое строение корневищ однодольных и двудольных несколько различается.
Рис. 1.34. Корневища однодольных растении: А — ландыша; Б — пырея; 1 — эпидерма; 2 — запасающая паренхима коры; 3 — эндодерма; 4 — склеренхима; 5 — закрытый коллатеральный пучок; 6 — центрофлоэмный пучок; 7 — запасающая паренхима центрального цилиндра; а — ксилема; б — флоэма

К характерным анатомическим признакам корневищ однодольных можно отнести следующие: покровная ткань — многолетняя, иногда одревесневающая эпидерма без устьиц, трихом и кутикулы; первичная кора хорошо развита, представлена многорядной запасающей паренхимой и 1-, 2-, реже многорядной эндодермой с U-образными утолщениями клеточных оболочек; центральный цилиндр содержит коллатеральные закрытые или центрофлоэмные пучки, расположенные по всему цилиндру; сердцевина более или менее выражена, не выражена или полая.
СТЕБЛИ ДВУДОЛЬНЫХ ТРАВЯНИСТЫХ РАСТЕНИЙ
У двудольных растений первичное строение стеблей быстро сменяется вторичным в результате появления и деятельности камбия. В зависимости от характера заложения и дальнейшей дифференциации камбия у травянистых растений центральный цилиндр может быть пучкового, беспучкового и переходного типа строения (рис. 1.35,1.36). Первичная кора обычно хорошо развита, включает коровую хлорофиллоносную и запасающую паренхиму, а зачастую и колленхиму. Эндодерма, зачастую, хорошо выражена и отделяет кору от центрального цилиндра. В клетках эндодермы может находиться оберегаемый крахмал, тогда она называется крахмалоносным влагалищем, или кристаллы оксалата кальция, и тогда она называется кристаллоносным влагалищем. Сердцевина стебля бывает выполненной однородной или неоднородной, а также может быть полой (сем. зонтичные, гвоздичные, губоцветные).
Пучковый тип строения стебля (рис. 1.35, 1.36, Б, В, 1.37, А, 1.38) характеризуется наличием постоянных коллатеральных или биколлатеральных пучков, образованных сначала тяжами прокамбия, а затем — пучкового камбия. Разделены пучки живой или одревесневшей паренхимой, или склеренхимой. В междоузлиях пучки располагаются упорядоченно — по кругу, так как листовые следы в узлах объединяются в синтетические пучки и сливаются с пучками стебля (рис. 1.35). Пучки открытые и постоянно пополняются тканями.
Рис. 1.35. Схемы прохождения и расположения проводящих пучков в стебле двудольного травянистого растения: А — поперечный срез; Б — продольные срезы (объемный и развернутый в плоскости); 1 — первичная кора; 2 — центральный цилиндр; 3 — листовые следы: 4 — стеблевые пучки

В стеблях переходного типа (рис. 1.36, Г, Д, 1.37, Б) из лучевой паренхимы появляется межпучковый камбий, дающий дополнительные проводящие пучки. Они находятся между основными пучками и всегда мельче последних. Основные и дополнительные пучки при разрастании могут сливаться, образуя неравномерное по толщине кольцо ксилемы. Флоэма или сливается в более тонкое неравномерное кольцо, или располагается участками, разделенными лучевой паренхимой, склеренхимой или склереидами. Такой переход от пучкового строения стебля через переходной тип к непучковому характерен для представителей сложноцветных, бобовых, губоцветных, капустных и др.

Изначально непучковый, или беспучковый, тип (рис. 1.36, Е, 1.37, В, 1.38, В) характеризуется отсутствием проводящих пучков, поскольку прокамбий, а затем и камбий закладываются кольцом. Проводящая система состоит из внутреннего широкого по толщине цилиндра ксилемы и более узкого наружного цилиндра флоэмы. На поперечных срезах ксилема и флоэма имеют кольцевое расположение (сем. гвоздичные, льновые, мареновые и др.).
Рис. 1.36. Развитие тканей в стеблях двудольных растении и формирование различных типов анатомического строения (схемы): А — продольный разрез; Б — Е — поперечные срезы стеблей разных типов строения в трех зонах — заложения прокамбия (а), появления камбия (б), вторичного утолщения (в); Б — первичное пучковое строение; В — вторичное пучковое строение с паренхимой и склеренхимой между пучками; Г — переходной тип; Д — беспучковый тип, сформированный из переходного: Е — беспучковый тип, сформированный однородным кольцом камбия; 1 — верхушечная меристема; 2 — протодерма; 3 — основная меристема; 4 — сердцевина; 5 — прокамбий; 6 — эпидерма; 7 — первичная кора; 8 — первичная флоэма; 9 — первичная ксилема; 10 — камбий; 11 — перицикл; 12 — вторичная флоэма; 13 — вторичная ксилема; 14 — паренхима сердцевинных лучей; 15 — склеренхима

Различные систематические группы растений и виды имеют определенные индивидуальные признаки анатомического строения (рис. 1.37, 1.38), которые используются как диагностические в систематике и при идентификации растений и растительного сырья.
Рис. 1.37. Примеры строения стеблей двудольных травянистых растений: А — пучкового типа (клевер); Б — переходного типа (подсолнечник); В — беспучкового типа (лен); 1 — эпидерма: 2 — пластинчатая колленхима; 3 — коровая паренхима; 4 — схизогенные каналы; 5 — крахмалоносная эндодерма; 6 — открытый коллатеральный пучок; 7 — толстостенная флоэма; 8 — тонкостенная флоэма; 9 — пучковый камбий; 10— межпучковый камбий; 11 — дополнительный пучок; 12 — ксилема; 13 — одревесневшая межпучковая паренхима; 14 — лучевая паренхима; 15 — сердцевина; а — склеренхимные волокна; б — лубяные волокна; в — паренхима; г — ситовидные трубки с клетками-спутницами; д, е, ж, з, и — сосуды: пористые, сетчатый, лестничный, спиральные и кольчатый; к — паренхима; л — либриформ

Рис. 1.38. Стебель тыквы: А — схема; Б — фрагмент поперечного среза; 1 — эпидерма; 2,3— первичная кора: 2 — уголковая колленхима в ребрах и межреберьях; 3 — хлоренхима; 4 —7 — центральный цилиндр: 4 — перициклическая склеренхима; 5 — основная паренхима; 6 — полость; 7 — биколлатеральный проводящий пучок: а — флоэма наружная и внутренняя; б — многорядный камбий; в — ксилема

КОРНЕВИЩА ДВУДОЛЬНЫХ
Корневища двудольных могут иметь пучковое, беспучковое и переходное строение. Они во многом сходны с надземными стеблями, но являются запасающими подземными органами и отличаются от надземного стебля следующим: покрыты чаще всего перидермой, не содержат в коре хлоренхимы, в них слабее развиты механические и проводящие ткани, а особенно хорошо представлена во всех частях паренхима (рис. 1.39). Она накапливает не только резервные питательные продукты, но и биологически активные вещества, что обусловливает использование корневищ многих растений в качестве пищевого и лекарственного растительного сырья. Довольно часто в корневищах имеются специализированные эндогенные секреторные структуры, что учитывается при диагностике растений и растительного сырья.
Рис. 1.39. Корневище мать-и-мачехи (схема части поперечного среза): 1 — перидерма; 2 — запасающая паренхима коры; 3 — полости со слизью; 4 — проводящие пучки коры; 5 — эндодерма; 6 — открытые коллатеральные пучки; 7 — сердцевина с полостью в центре

СТЕБЛИ ДРЕВЕСНЫХ ДВУДОЛЬНЫХ РАСТЕНИЙ
Для стеблей древесных характерно вторичное беспучковое строение. Покровная ткань — перидерма с чечевичками или корка. В древесине вследствие сезонной деятельности камбия образуются годичные кольца прироста, состоящие из осенних трахеальных элементов — трахеид и либриформа, и весенних трахеальных элементов — сосудов, трахеид с окаймленными порами, древесинной паренхимы (рис. 1.40).
Рис. 1.40. Фрагменты поперечных срезов стебля липы: А — схема; Б — сегмент в плане; В — фрагмент луба (вторичной коры); Г — фрагмент древесины; 1 — перидерма; 2 — колленхима; 3 — паренхима первичной коры; 4 — друзы; 5 — лубяные волокна; 6 — тонкостенные элементы луба — ситовидные трубки с клетками-спутницами; 7 — сердцевинные лучи; 8 — камбий; 9 — годичные кольца древесины; 10 — весенние элементы годичного кольца; 11 — осенние элементы годичного кольца; 12 — сердцевина

Сердцевинные лучи одно- или многорядные, первичные (соединяющие сердцевину с корой) и вторичные (не доходящие до сердцевины). Диагностическое значение имеет толщина перидермы, состав и расположение гистологических элементов древесины, вторичной коры — твердого (толстостенного) и мягкого (тонкостенного) луба. Древесина может быть лучистой, кольце-сосудистой и рассеянно-сосудистой (рис. 1.41).
Рис. 1.41. Типы древесины: 1 — кольце-сосудистая; 2 — рассеянно-сосудистая

СТЕБЛИ ХВОЙНЫХ РАСТЕНИЙ
Анатомическое строение стеблей хвойных отличается от покрытосеменных следующим: перидерма обычно без чечевичек, но с трещинами; на стволе рано начинает формироваться корка; в коре отсутствуют или слабо развиты механические ткани; луб включает ситовидные клетки или ситовидные трубки без клеток-спутниц, паренхиму, иногда — лубяные волокна или склереиды; годичные кольца древесины состоят из весенних трахеид с окаймленными порами, выполняющими проводящую функцию, и осенних волокнистых трахеид без окаймленных пор, выполняющих опорную функцию; все части стебля содержат, как правило, схизогенные смоляные ходы. Типичное строение имеет стебель сосны обыкновенной (рис. 1.42).
Рис. 1.42. Анатомическое строение стебля сосны: А, Б, В — поперечные срезы; Г, Д — древесина на продольных срезах — радиальном и тангентальном; 1 — перидерма; 2 — коровая паренхима; 3 — смоляные ходы; 4 —11 — центральный цилиндр; 4 — луб; 5 — камбий; 6 — годичные кольца древесины; 7 — весенние трахеиды, 8 — осенние трахеиды; 9 — сердцевинные лучи; 10 — сердцевина

Биологическая библиотека - материалы для студентов, учителей, учеников и их родителей.
Наш сайт не претендует на авторство размещенных материалов. Мы только конвертируем в удобный формат материалы, которые находятся в открытом доступе и присланные нашими посетителями.
Если вы являетесь обладателем авторского права на любой размещенный у нас материал и намерены удалить его или получить ссылки на место коммерческого размещения материалов, обратитесь для согласования к администратору сайта.
Разрешается копировать материалы с обязательной гипертекстовой ссылкой на сайт, будьте благодарными мы затратили много усилий чтобы привести информацию в удобный вид.
Posted in Биология Tags: Ботаника

Стебель — это осевая структура побега. И его анатомическое строение вытекает из главных функций. Во-первых, для стебля характерно развитие механической и проводящей тканей. Во-вторых, стебель имеет сложную систему меристем: верхушечные, боковые и вставочные, которые определяют его нарастание в течение длительного времени, а также возникновение новых органов. Возникает стебель из апикальной меристемы, из которой дифференцируются три слоя тканей: основная, покровная, проводящая.
Строение однодольных растений
Весь прокамбий или первичная меристема у однодольных растений дифференцируется в элементы первичных проводящих тканей. Их стебли, особенно, если рассматривать травянистые (злаки), имеют более простое строение по сравнению со стеблями двудольных растений. Для них также в основном характерно первичное строение. У однодольных растений сосудисто-волокнистые проводящие пучки закрытые (без камбия), состоят только из первичных тканей ирасположены беспорядочно в основной паренхиме стебля.

Структура стебля однодольных растений
Строение двудольных растений
В средней части прокамбиального тяжа у двудольных растений происходит образование камбия и начинается формирование вторичных проводящих тканей (метафлоэмы и метаксилемы). Объём этих вторичных проводящих тканей увеличивается за счёт деления клеток камбия, что в итоге приводит к сильному утолщению стебля.
Для двудольных характерны открытые пучки с камбием. Проводящие ткани в стеблях двудольных растений располагаются кольцом вокруг сердцевины. Пучки разделяются сердцевинными лучами, которые состоят из паренхимы и соединяют сердцевину или с перициклом, или с первичной корой. По периферии в стеблях двудольных растений располагаются механические ткани, при этом склеренхима входит в состав перицикла, колленхима, вместе с основной паренхимой, входит в состав первичной коры.
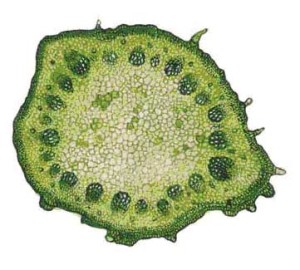
Строение стебля двудольных растений
Строение центрального цилиндра двудольных может быть пучковым и непучковым.
По своей анатомической структуре однолетние стебли древесных растений сходны со стеблями травянистых растений: их проводящие системы имеют непучковое строение. Их отличительной чертой является активная деятельность камбия и раннее формирование вторичной покровной ткани — пробки.
Благодаря деятельности камбия в стебле образуются различные элементы вторичного происхождения, а первичные элементы при этом постепенно исчезают. Камбий формирует элементы ксилемы и флоэмы с различной скоростью: на одну клетку флоэмы камбий отделяет несколько клеток ксилемы. В результате этого ксилема (или древесина) нарастает намного быстрее, чем флоэма (луб), соответственно, почти вся масса ствола и ветвей дерева приходится на долю ксилемы. Флоэма, напротив, составляет сравнительно тонкий слой вторичной коры.

Строение стебля у древесных растений
Сплошное камбиальное кольцо у большинства древесных растений (в качестве примера можно взять липу) образуется в самом начале формирования стебля.
Деление клеток камбия происходит тангентально, что определяет расположение клеток правильными рядами по радиусу. При нарастании ксилемы камбий сдвигается ближе к периферии и его окружность увеличивается за счёт радиального деления его клеток.
При вторичном утолщении вторичную ксилему или древесину с паренхимными лучами составляет все, что откладывается вовнутрь ствола, а все то, что откладывается наружу, т.е. к периферии ствола, составляет вторичную флоэму или луб с сердцевинными лучами.
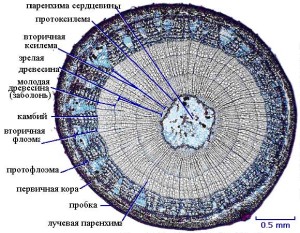
Строение стебля у древесных растений
Проводящая система ксилемы состоит из трахеид и сосудов. У хвойных растений только трахеиды выполняют функцию проведения, а у древесных лиственных растений — и трахеиды, и сосуды.
Появление годичных колец в ксилеме (древесине) происходит в результате периодической деятельности камбия. В весенний период, как правило, много воды и питательных веществ, поэтому камбий образует крупные элементы древесины с большим просветом и тонкими стенками. Ближе к концу вегетационного периода происходит затухание деятельности камбия и в древесине начинают преобладать механические элементы и узкие сосуды.
Сердцевинные лучи состоят из крупных паренхимных клеток, которые имеют прямоугольную форму. По своему происхождению эти лучи могут быть первичными и вторичными. Первичные лучи тянутся от первичной коры до сердцевины, и они длиннее вторичных. Основная функция лучей — проведения воды и органических веществ в горизонтальном направлении.
Механическая ткань в ксилеме (древесине) состоит из толстостенных узких и уже одревесневших клеток.
В состав вторичной флоэмы тоже входят 3 типа тканей: основная, механическая и проводящая. Флоэмные сердцевинные лучи проходят от камбия наружу, и стенки их клеток не одревесневают. В своей периферийной части лучи сильно расширяются. Механическую ткань вторичной флоэмы называют вторичной склеренхимой, она представлена лубяными волокнами (твёрдый луб). Проводящая ткань представляет собой ситовидные трубки с клетками-спутницами (мягкий луб).
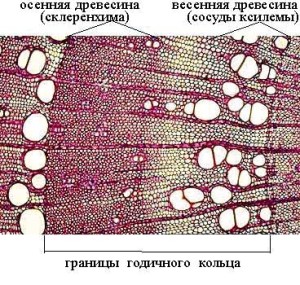
Годичное кольцо у деревьев
Утолщение стебля является результатом активной деятельности камбия. Эпидерма из-за внутреннего давления разрывается и в местах разрыва под эпидермой образуется пробковый камбий (или феллоген). Клетки феллогена делятся, при этом наружу откладываются клетки пробки (или феллемы), а вовнутрь — клетки феллодермы. Эти 3 слоя: феллема, феллоген и феллодерма, и образуют вторичную покровную ткань — перидерму (пробку).
В следующем разделе мы рассмотрим метаморфозы побега.
Информация о статье:
В статье рассказывается об анатомии стебля. Описываются различия в строении стеблей однодольных и двудольных растений.
Анатомическое строение стебля двудольных и однодольных растений резко различно.
У однодольных растений разделения стебля на кору, проводящую ткань и сердцевину не наблюдается благодаря разбросанному расположению сосудистых пучков по всей поверхности среза (рис. 107). Это характерно для стебля лилейных, пальм и некоторых злаков, имеющих сплошной стебель (например, у кукурузы, сорго, сахарного тростника). У большинства же злаков срединная часть стебля, соответствующая сердцевине, атрофируется, и стебель становится полым внутри (соломина). В этом случае пучки расположены в ткани стебля более правильно, в периферических слоях стебля, по кругу. Кора у однодольных имеется, но не бывает так резко выражена, как у двудольных растений.
У однодольных, имеющих длинные междоузлия, пучки, выйдя из листа (общие пучки), сначала приближаются к центру стебля, затем идут вдоль стебля отвесно, а ниже приближаются к периферии и там уже присоединяются своими нижними концами к другим пучкам. При прохождении в центральной части стебля пучки утолщаются, и поэтому в центре поперечного разреза расположены более крупные пучки. Такое расположение пучков имеет место, например, у пальмы, аспидистры (пальмовый тип). У однодольных с меньшим развитием междоузлий, например у наших злаков, пучки тянутся отвесно.
На поперечном разрезе стебля драцены от периферии к центру сменяют друг друга следующие элементы:
Стебли однодольных растений, так же как стебли Двудольных, имеют эпидермис, первичную кору и центральный цилиндр. Закрытые проводящие пучки расположены по всей толще центрального цилиндра.
Развитию проводящей системы стебля предшествует заложение в основании листовых зачатков, находящихся под конусом нарастания побега, многочисленных тяжей прокамбия, из которых развиваются пучки листовых следов. В стебле однодольного растения образовательное кольцо не выражено. Прокамбиальные тяжи закладываются и дифференцируются в проводящие пучки неодновременно. Войдя в стебель, наиболее крупные пучки, которые развиваются в первую очередь, дуговидно изгибаются, доходят почти до середины стебля и на протяжении нескольких междоузлий идут более или менее вертикально. Затем они изгибаются к периферии и соединяются с пучками листовых следов ниже расположенных листьев. Более мелкие, позднее дифференцирующиеся пучки тоже дуговидно изгибаются, но они располагаются ближе к периферии центрального цилиндра, а некоторые из них проходят даже вдоль первичной коры стебля. Таким образом, проводящая система стебля представлена многочисленными пучками. Расположением проводящих пучков объясняются некоторые особенности анатомической топографии стебля однодольных: сердцевины у большинства представителей нет, первичная кора не всегда резко отграничена от центрального цилиндра. Эндодерма хорошо выражена лишь в подземных органах — корневищах.
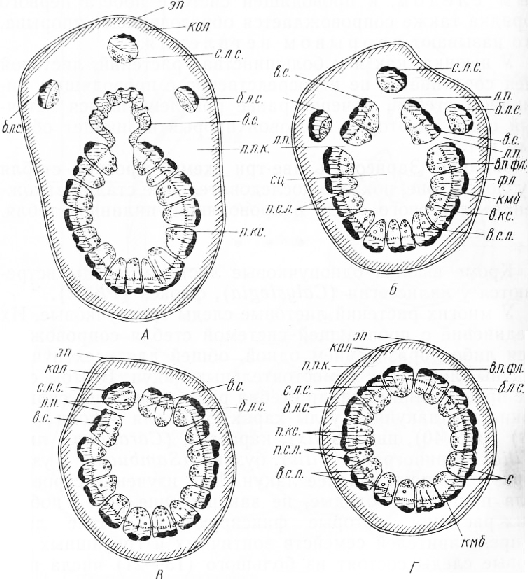
Рис. 1. Строение трехлакунного узла яблони. А — Г —последовательные стадии соединения веточного и листовых следов с проводящей системой материнского стебля: с. л. с. — срединный и б. л. с. — боковой пучки листовых следов, в. с. — веточный след, л. п. — листовые прорывы, эп — эпидермис, кол — колленхима, п. п. к. — паренхима первичной коры, в. п. фл. — волокна первичной флоэмы, фл — флоэма, в. кс. — вторичная ксилема, п. кс. — первичная ксилема, с — сосуды, п. с. л. — первичные и в. с. л. — вторичные сердцевинные лучи, кмб — камбий, сц — сердцевина
Проводящие пучки в стеблях однодольных растений коллатеральные, реже концентрические, закрытые (без камбия). Все проводящие элементы первичного (прокамбиального) происхождения. Вторичное утолщение встречается лишь у некоторых древовидных растений, но у них оно осуществляется иначе, чем у двудольных.
С особенностями анатомической структуры стеблей однодольных можно ознакомиться на примере растений из семейств лилейных, касатиковых, злаков.
Поперечные срезы обрабатывают флороглюцином и соляной кислотой и заключают в глицерин. До проведения реакции на одревеснение на срезы можно подействовать раствором иода в водном растворе иодистого калия, чтобы выявить клетки, содержащие запасной крахмал.
Стебель купены лекарственной (Polygonatum officinale All.)
На поперечном срезе стебля купены можно выделить три зоны:
1) покровную, представленную эпидермисом, 2) первичную кору, 3) центральный цилиндр.
Эпидермис имеет обычное строение. Однако в более старых участках стеблей стенки его клеток нередко одревесневают. Так как у однодольных растений нет вторичного утолщения, то утрата эластичности оболочками клеток эпидермиса существенного значения не имеет.
Первичная кора состоит из тонкостенных паренхимных клеток. Колленхимы, как и у большинства однодольных, нет. Эндодерма не выражена.
Вся остальная часть центрального цилиндра занята тонкостенными клетками основной паренхимы с большим числом закрытых коллатеральных проводящих пучков.
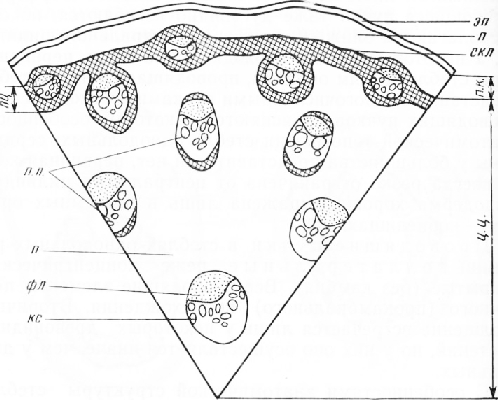
Рис. 2. Схема строения стебля купены (рисунок Г. Б. Кедрова) : эп — эпидермис, п. к. — первичная кора, ц. ц. — центральный цилиндр, пц — перицикл, л —паренхима, скл — склеренхима, фл — флоэма, кс— ксилема, п. п. — проводящие пучки
Ксилемная часть пучка, состоящая преимущественно из пористых трахеид, на поперечных срезах нередко имеет очертания широкой латинской буквы V, в углублении которой находятся группы ситовидных трубок с сопровождающими клетками.
Пучки располагаются по всей центральной части стебля, некоторые из них находятся в склеренхиме перицикла и даже в первичной коре. Пучки, примыкающие к склеренхиме перицикла, окружены механическими волокнами. В связи с этим наружная и внутренняя границы перицикла имеют извилистые очертания.
Задание. Зарисовать схему строения стебля, отметив эпидермис, паренхимную первичную кору, центральный цилиндр с перициклом и проводящими пучками.
Вместо купены можно использовать стебли тюльпана (Tulipa), спаржи (Asparagus), эремуруса (Eremurus), у которых также хорошо выражен склеренхимный пери-цикл.
Стебель кукурузы обыкновенной (Zea mays L.)
Строение стебля кукурузы имеет много общего со строением стебля купены. Различия заключаются в том, что в стебле кукурузы, как правило, отсутствует первичная кора, которая заметна лишь в самых нижних междоузлиях в виде немногих слоев тонкостенных паренхимных клеток. Склеренхима перицикла придвинута вплотную к эпидермису.
Закрытые коллатеральные проводящие пучки имеют своеобразное строение. В состав ксилемы входит от 3 до 5 сосудов, из них 1—3— узкопросветные, составляющие короткую радиальную цепочку. Это сосуды протоксилемы с кольчатыми и спиральными утолщениями стенок. Они расположены на границе с межклетником (так называемой лакуной протоксилемы) , а иногда вдаются в него. Метаксилема представлена двумя крупными пористыми сосудами и небольшим числом расположенных между ними волокнистых трахеид или волокон с одревесневшими стенками.
Флоэма примыкает к ксилеме с наружной стороны. На поперечных срезах она имеет вид сеточки, у которой шестиугольные ячеи образованы перерезанными ситовидными трубками, а узлы — четырехугольными сопровождающими клетками с зернистой цитоплазмой.
Проводящие элементы пучка окружены склерен-химным влагалищем или обкладкой, состоягцей из толстостенных волокон с одревесневшими стенками. Склеренхимное влагалище хорошо выражено над флоэмой и с внутренней стороны ксилемы, а также по сторонам двух крупных сосудов метаксилемы. Такое строение проводящих пучков характерно для злаков и представителей близких к ним семейств, например осоковых.
Очертания проводящих пучков и степень развития окружающей их механической ткани различны. Пучки, лежащие в центральной части стебля, несколько вытянуты в радиальном направлении. Вытянута также водоносная полость у основания протоксилемы (лакуна про-токсилемы). Склеренхимная обкладка выражена слабо и состоит из сравнительно небольших групп толстостенных волокон, располагающихся над флоэмой, с внутренней стороны ксилемы и по бокам пучка у крупных пористых сосудов метаксилемы.
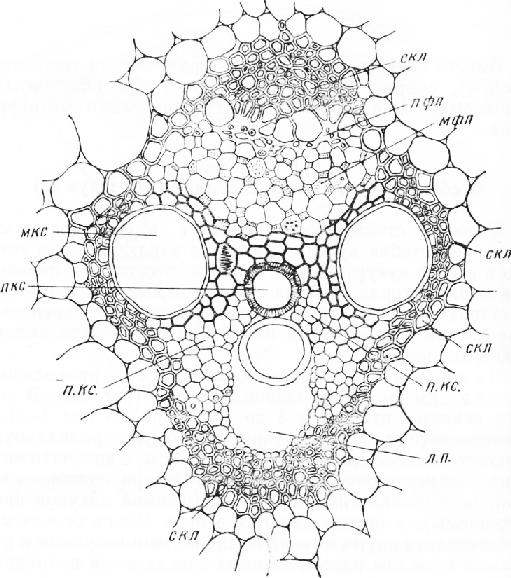
Рис. 3. Проводящий пучок кукурузы (из Ростовцева, 1948): скл — склеренхима, пфл — протофлоэма, мфл — метафлоэма, мкс — метаксилема, п. кс. — паренхима ксилемы, л. п. — лакуна протоксиле-мы, пкс — протоксилема
Ближе к периферии стебля пучки более или менее округлые, часто с одним сосудом протоксилемы. Лакуна небольшая, у самых периферических пучков ее может не быть. Механическая обкладка шире, чем у внутренних пучков. Наибольшей мощности она достигает у пучков, расположенных близ эпидермиса. Оболочки паренхимных клеток, окружающих периферические пучки, одревесневают. Таким образом, под эпидермисом возникает сплошное кольцо одревесневших тканей: перицикличе-ской склеренхимы, паренхимы и обкладки проводящих пучков.
Такое расположение механических тканей увеличивает прочность периферической части стебля на сжатие и растяжение, которые возникают при изгибе органа. Центральная часть стебля при изгибе испытывает наименьшие напряжения.
Задание.
1. При малом увеличении зарисовать схему строения стебля.
2. При большом увеличении сделать детальный рисунок проводящего пучка.
Стебель ржи посевной (Secaie cereale L.)
В отличие от кукурузы центральная часть междоузлия стебля (соломины) ржи занята крупной воздушной полостью.
Снаружи стебель покрыт эпидермисом с устьицами. Стенки эпидермальных клеток обычно одревесневают. Под эпидермисом находится склеренхима, состоящая из плотно сомкнутых волокон с сильно одревесневшими стенками, в которых хорошо заметны поровые каналы. В склеренхиму погружены островки ассимиляционной ткани, состоящей из тонкостенных паренхимных клеток с хлоропластами.
Мощность ассимиляционной ткани и склеренхимы в разных частях междоузлия неодинакова. В верхней, хорошо освещаемой части междоузлия островки ассимиляционной ткани крупнее, чем в нижней части, окруженной влагалищем листа и поэтому находящейся в худших условиях освещения. В основании междоузлия ассимиляционная ткань часто не развивается.
Между участками ассимиляционной ткани, в глубже лежащих слоях склеренхимы расположены мелкие проводящие пучки типичного для злаков строения. К центру от них, в широком слое паренхимы с одревесневающими к концу вегетации оболочками клеток находятся более крупные пучки. Они лежат не строго по кругу, а несколько смещены один относительно другого. Наружные из них почти вплотную придвинуты к склеренхиме, и склеренхимный тяж, сопровождающий их флоэму, соединяется с субэпидермальной склеренхимой, внутренняя граница которой приобретает неровные очертания. Расположение мощно развитой склеренхимы на периферии стебля обусловливает создание чрезвычайно экономной и совершенной в механическом отношении конструкции, устойчивой на изгиб и на излом. Внутренние пучки более крупные. Со стороны флоэмы они также снабжены тяжом склеренхимы, который, однако, не соединен с наружной склеренхимой. Эти пучки окружены паренхимными клетками, стенки которых к концу вегетационного периода обычно одревесневают.
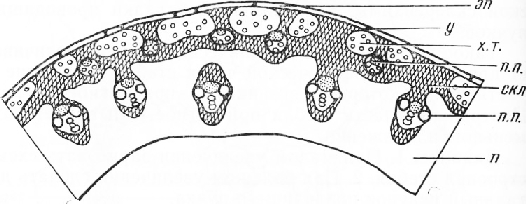
Рис. 4. Схема строения соломины ржи: эп — эпидермис, у — устьица, х. т. — хлорофиллоносная ткань, скл — склеренхима, п. п. — проводящие пучки, п — паренхима
Задание. Зарисовать схему строения стебля при малом увеличении микроскопа.
Строение, сходное с описанным, имеют стебли овса, пшеницы, ячменя и других злаков.
Читайте также: