
Строение проводящих пучков жилок кратко
Обновлено: 05.07.2024
Элементы флоэмы и ксилемы в органах растений располагаются рядом, образуя часто проводящие пучки. В начале формирования структуры органов проводящие пучки образуются из прокамбия. У однодольных растений после израсходования этой первичной образовательной ткани дальнейшее разрастание пучка прекращается. Такие пучки, не содержащие уже образовательной ткани, называются закрытыми.
У двудольных растений и голосеменных из прокамбия возникает камбий, за счёт которого происходит разрастание пучка. Проводящие пучки, содержащие камбий, называются открытыми. Из прокамбия и камбия образуются проводящие и механические элементы ксилемы и флоэмы. Элементы ксилемы и флоэмы, сформированные деятельностью прокамбия, называются первичными, а деятельностью камбия — вторичными. За счёт деятельности камбия в открытых проводящих пучках ежегодно образуются новые слои ксилемы и флоэмы, что обеспечивает утолщение органов растений.
В зависимости от взаимного расположения ксилемы и флоэмы различают четыре типа проводящих пучков: коллатеральные, биколлатеральные, концентрические и радиальные (рис. 15).
-
В коллатеральных пучках флоэма и ксилема располагаются рядом. В стебле и корне ксилема обычно расположена ближе к центру, а флоэма — к периферии.
Фотосинтез — процесс, при котором зелёные растения, используя солнечный свет, из неорганических веществ производят органические вещества.
Изучение микроскопического строения листовой пластинки позволит понять механизм фотосинтеза и функции зелёных листьев.

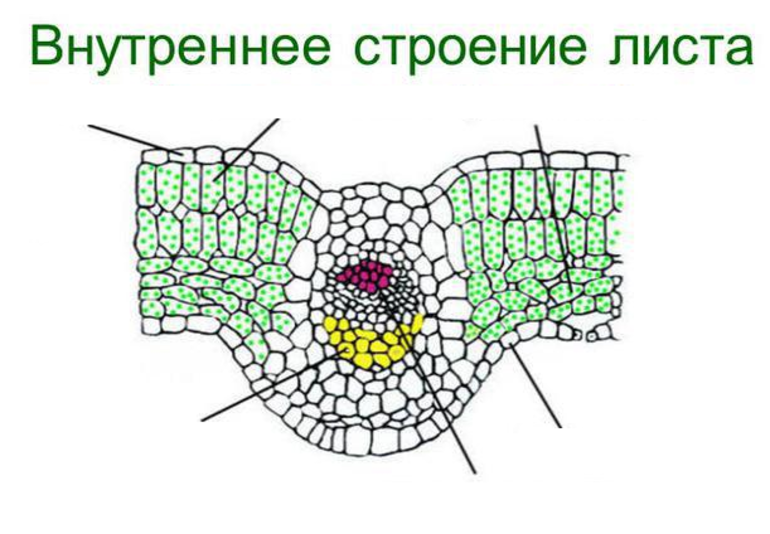
Если рассмотреть поперечный срез листовой пластинки под микроскопом, то можно заметить, что сверху и снизу он покрыт тонкой бесцветной кожицей, а внутри находятся зелёные клетки. Хорошо видна жилка, или проводящий пучок.
На верхней и нижней поверхностях листа находится кожица ( эпидермис , эпидерма ). Это разновидность покровной ткани, которая защищает клетки от механических повреждений и от высыхания, а также обеспечивает газообмен и испарение воды.
На поверхности кожицы находится кутикула — восковой слой (восковой налёт), который предотвращает потерю воды. Растения с толстой кутикулой испаряют меньше воды, чем растения с тонкой кутикулой.
В клетках мякоти имеются хлоропласты , в которых происходит фотосинтез . В мякоти присутствуют две разновидности основной ткани: столбчатая и губчатая. Столбчатая ткань находится под верхним эпидермисом. Она состоит из нескольких слоёв продолговатых клеток, содержащих большое количество хлоропластов.
Под столбчатой тканью расположены клетки губчатой ткани. Эти клетки округлые и расположены рыхло. Между ними много межклетников , заполненных воздухом. В клетках губчатой ткани меньше хлоропластов по сравнению со столбчатой тканью.
На поперечном срезе листовой пластинки под микроскопом хорошо видны жилки (проводящие пучки). Они образованы сосудами, ситовидными трубками и волокнами.

Волокна имеют толстые стенки. Они выполняют опорную функцию. Сосуды обеспечивают поступление к клеткам листа воды и минеральных солей, а ситовидные трубки — отток образовавшихся в ходе фотосинтеза органических веществ к другим органам растения.
Жилки — это проводящие пучки, соединяющие стебли и листья. Главная функция этой части растения — передача питательных веществ. Различают несколько типов жилкования листа — параллельное, дуговое, сетчатое и пальчатое, а также несколько нестандартных вариаций. Как правило, система жил у однодольных проще, чем у двудольных.
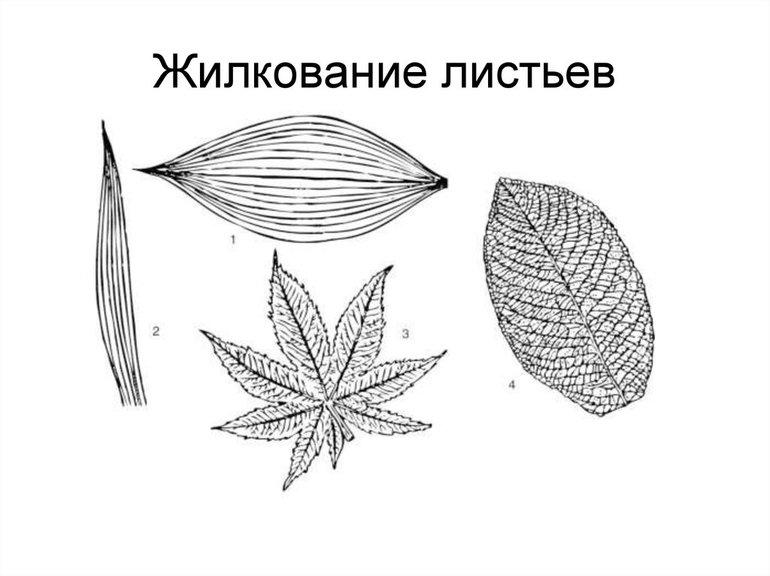
Типы и виды
Начальная точка у всех жилок находится у черешка, который соединяется с листовой пластинкой. Из неё выходит один или несколько главных сосудов, от которых ответвляются более мелкие и тонкие отростки, почти или полностью доходящие до краёв листа. Самые маленькие из них находятся на периферии, а также в центре. Рисунок жил нередко повторяет форму разветвления ветвей растения.
Типы жилкования зависят от многих факторов и являются важным систематическим признаком, по которому представителей флоры относят к тому или иному классу. Определить их довольно легко — достаточно посмотреть на растение.
Основные из разновидностей:
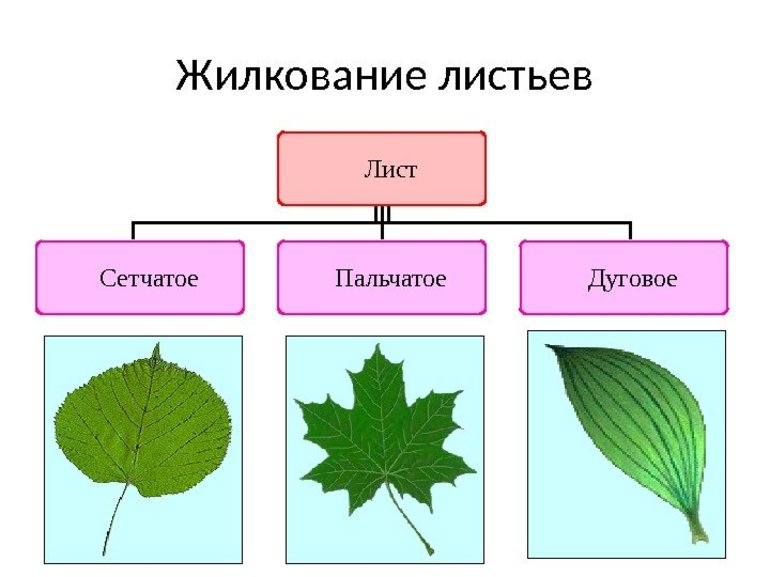

- Дуговое. Несколько одинаковых по размерам жил выходят из черенка и опоясывают лист, изгибаясь дугами, сходящимися у основания и верхушки. Центральная ось прямая. Такое жилкование характерно для ландыша, тюльпана и других простых однодольных растений. Некоторые двудольные, такие как подорожник, тоже используют эту разновидность, но у них прожилки соединяются наверху.
- Параллельное. При таком типе несколько крупных жил проходят от основания до конца листа по оси параллельно друг другу. Жилки имеют одинаковый размер и толщину. Примеры жилкования листьев подобного вида включают большинство сельскохозяйственных культур — кукурузу, пшеницу, просо и другие однодольные злаковые.
- Сетчатое или перистое. По центру листа проходит одна крупная жила, от которой отходит множество мелких. Они постепенно уменьшаются в длине и толщине по мере удаления от центральной оси, покрывая практически всю пластину сеткой. Такой тип жилкования является самым распространённым среди высокоразвитых двудольных растений, поскольку он является наиболее совершенным и эффективным. Он характерен почти для всех представителей розоцветных, капустных, бобовых, паслёновых, астровых. Например, для яблони, липы, осины, клёна, черёмухи.
- Пальчатое. Состоит из нескольких одинаковых главных жилок, от которых отходят уменьшающиеся отростки. В таблице жилкования листьев сюда принадлежат клён, люпин, лютик.
- Дихотомическое. Главная жилка ветвится вильчато, а её отростки доходят до краёв листа. Под такое описание подходят большинство папоротниковых и гинкго.
- Одна неветвящаяся жила. Такое жилкование бывает у простых растений — плаунов и хвощей, а также у большинства хвойных.
Существуют также нестандартные виды жилкования — радиальное, продольное, поперечное, веерное, перестопетлевое, перистокраебежное. Между ними есть смешанные и переходные формы, усложняющие классификацию.
Функции жилок
Жилки листа выполняют функции, от которых зависит нормальная работа и жизнь всего организма растения. Их основная роль:
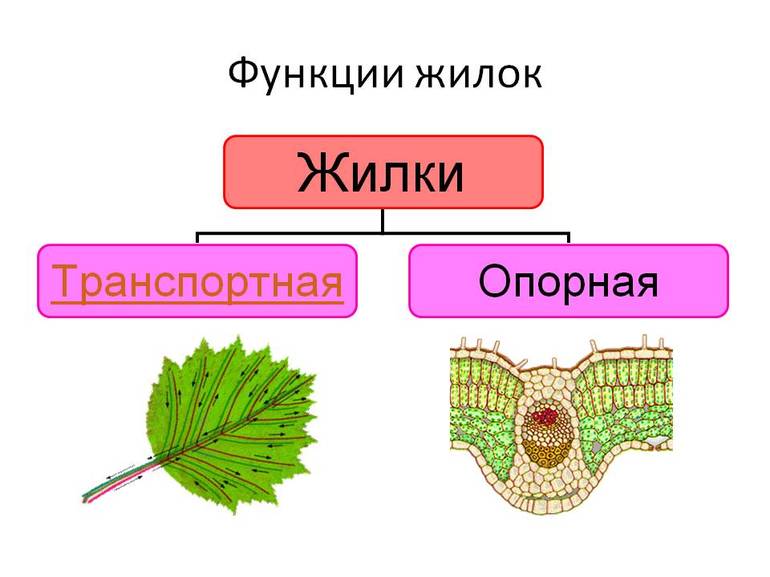
- Транспортная. Жилки снабжают листья водой и минеральными веществами, а также передают продукты фотосинтеза корням, плодам, почками и другим частям растения. Они являются частью сложной системы сосудов и сетчатых трубок, ответственных за транспортировку минералов и питательных веществ по всему растению. Поскольку большая их часть впервые появляется именно в листьях, жилки необходимы для поддержания нормальной жизнедеятельности.
- Механическая. Жилки — опора для листовой паренхимной ткани. Они защищают лист от разрыва, увеличивая его прочность, а также соединяют основание с листовой пластиной, обеспечивая сбор питательных веществ со всей площади, и объединяют листок со стеблем через черешок (у черешковых растений) или напрямую (у сидячих).
Система жилок служит связью листа с основной частью растения на внешнем и внутреннем уровне. Благодаря ей, возможны все обменные процессы с участием хлорофилла и продуктов его переработки.
Внутреннее строение
Жилки покрывают проводящую основу листовой пластинки, перенося из неё различные вещества к другим частям растения. Они представляют собой проводящие пучки, состоящие из специфических тканей — луба и древесины. Основу составляет первый тип.
Лубовая ткань образована из вытянутых в длину живых безъядерных клеток. В месте их соприкосновения друг с другом образуются небольшие отверстия, через которые передается вода, раствор сахара и других минеральных веществ. Прилегая друг другу, такие структуры образуют трубки. Они подходят для передачи органических составов на большие расстояния.
По краям трубок расположены клетки-спутницы. Они отличаются небольшими размерами, но крупными полиплоидными ядрами и митохондриями. Их функция — управление деятельностью трубок путём передачи им нуклеинов и энергии в форме АТФ. Таким образом, спутницы регулируют нисходящий (проходящий от листьев к корням) ток питательных веществ.
Кроме луба, в состав жилок входит древесина, которую в биологии называют ксилемой. Она состоит из 3 типов функциональных элементов:
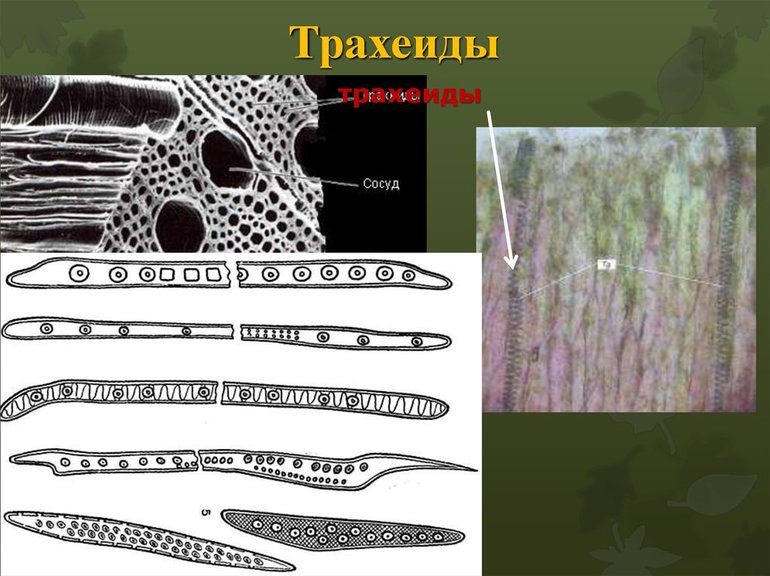
- Трахеиды. Это длинные и тонкие мёртвые клетки с толстой одеревеневшей оболочкой. Их поверхность покрыта порами, через которые вещества фильтруются их одной трахеиды в другую. Их функции — передача воды и растворов горизонтальной и вертикально.
- Паренхимные неполяризованные клетки. Примерно одинаковы по размерам. Образуют опорную ткань — паренхиму.
- Членики. Небольшие клетки, расположенные друг над другом и образующие длинную полую трубку — сосуд растения. Образуются из паренхимных клеток с больши́м ядром и тонкими стенками.
Древесина в жилках листа выполняет обратную лубу функцию. Она участвует в восходящем (проходящем от корней к листьям) токе, передавая зелёным тканям недостающие питательные вещества.
Кроме этик двух тканей, в состав жилок входят волокна. Они представляют собой вытянутые клетки с заострёнными кончиками и толстыми одревесневшими оболочками. Волокна окружают крупные жилки, выполняя защитную функцию.
Ситовидные трубки и сосуды
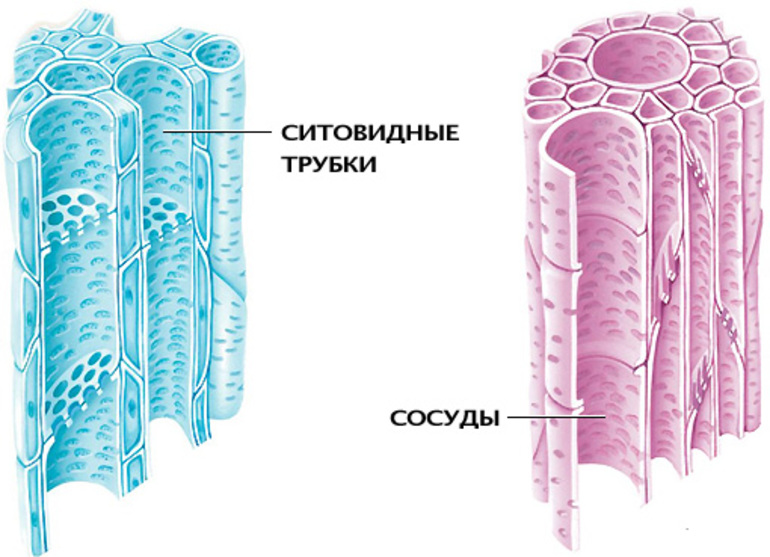
Древесина в составе жилок образует сосуды — важный элемент, необходимый для жизнедеятельности растения. Внешне они похожи на трубочки с плотными оболочками и внутренними утолщениями в форме колец или спиралей. Состоят сосуды из мёртвых клеток, но первоначально образовываются их живых паренхим.
Особенности строения сосудов обеспечивают высокое напряжение при заполнении их водой и минеральными растворами. Это обеспечивает возможность выполнения их основной функции — сила давления становится настолько высокой, что жидкость транспортируется снизу вверх от корня к листьям.
Из лубовой ткани образуются ситовидные трубки. Они получили своё название из-за внешнего строения, напоминающего решётку. Поперечные и внешние стенки структур содержат многочисленные отверстия, через которые транспортируется вода и полученные через фотосинтез питательные вещества. Давление внутри довольно низкое, потому движение происходит от листьев к корню.
В отличие от сосудов, ситовидные трубки жилок состоят из живых цитоплазменных клеток. Продолжительность их функционирования варьируется — у большинства растений она составляет не более года, но у некоторых паль может доходить до 10−20 лет. Отмирающие клетки превращаются в мозолистые тела, после чего деформируются и поглощаются организмом.
Жилки листа — важные элементы, от которых зависит нормальная жизнедеятельность всего растения. Они отвечают за транспортировку воды, минералов, продуктов фотосинтеза и других вещей к корням, листьям, плодам и прочему. В зависимости от формы листьев, вида крепления, способа членения, листорасположения и других характеристик растения меняется схема жилкования и строение отдельных жилок.
Важной частью листа являются проводящие пучки (жилки). В состав листа входят коллатеральные пучки закрытого типа, причём ксилема обращена к верхней эпидерме, а флоэма к нижней эпидерме. Очень часто в состав жилок входит механическая ткань, причём в состав крупных жилок входит склеренхима, а в состав мелких – колленхима. Склеренхима крупных жилок может располагаться под жилкой и над жилкой. Иногда механические ткани полностью окружают пучок.
У многих растений вокруг жилки располагаются обкладочные клетки проводящего пучка. Этот клетки мезофилла, которые в виде радиального кольца примыкают к жилке. Эти клетки могут иметь небольшое количество хлоропластов (и тогда они принимают незначительное участие в фотосинтезе) или не иметь их. Обкладочные клетки проводящего пучка обеспечивают транспорт водных растворов из ксилемы в мезофилл, а также переносят органические вещества из мезофилла во флоэму.
Типы жилкования: сетчатое, дуговое, параллельное. У некоторых папоротников и плаунов встречается дихотомическое жилкование. Достаточно часто в состав мезофилла входят клетки идиобласты. Это выделительные клетки, способные накапливать и выделять вещества вторичного метаболизма, при этом длительное хранение этих веществ происходит в мёртвых идиобластах.
Часто у растений с крупными листьями в мезофилле встречаются склереиды – одиночные мёртвые механические клетки звёздчатой формы с сильно утолщёнными клеточными стенками. Они выполняют механическую функцию.
Жилкование листа
У многих папоротников и архаичных семенных растений (гинкго) оно дихотомическое, или вильчатое.
У большинства хвойных в листе проходит одна или несколько продольных, не связанных между собой жилок.
У однодольных растений жилки листа соединяются с проводящей системой стебля через основание листа, не сливаясь друг с другом или сливаясь лишь близ верхушки. Между собой они обычно соединяются сетью мелких поперечных жилок – перемычек. В зависимости от особенностей прохождения продольных жилок в пластинке выделяют параллельное и дуговидное жилкование. О листе при этом говорят, что он параллельнонервный или дугонервный.
У двудольных известны два основных типа жилкования: перистое и пальчатое.
У листьев с перистым жилкованием (перистонервный лист) имеется одна главная жилка, являющаяся продолжением черешка. От главной жилки под углом отходят боковые жилки первого порядка, о них жилки второго порядка и т. д.
Иногда боковые жилки первого порядка доходят до края пластинки и оканчиваются здесь в лопастях, краях зубцов и т. д. Это так называемое перистокраебежное жилкование (ольха, берёза, вяз).
Если боковые жилки направляются к краю пластинки, но, не достигнув его, заворачивают дугой вперёд, соединяются со следующей боковой жилкой и образуют петлю, – это перистопетлевидное жилкование (многие двудольные).
Если боковые жилки последовательно ветвятся, в результате чего образуется всё более густая сеть без ясно выраженных петель, – это перистосетчатое жилкование (ива, груша, яблоня).
У листьев с пальчатым жилкованием (пальчатонервный лист) не имеется главной жилки. От места сочленения черешка с листовой пластинкой отходят несколько пальцевидно расходящихся крупных жилок первого порядка. Они могут доходить до края листовой пластинки (пальчатокраебежное – клён), или образовывать вдоль края петли (пальчатопетлевидное), или при свободном ветвлении жилки второго и последующего порядков образуют сетку (пальчатосетчатое).
Основные функции листа: фотосинтез, дыхание и газообмен (в том числе и транспирация). На свету фотосинтез резко преобладает над дыханием.
- колючки (кактус), в которые видоизменяется вся листовая пластинка. Они не участвуют ни в фотосинтезе, ни в транспирации, но защищают растение от поедания
- усики (бобовые), в которые видоизменяется верхняя часть листа. Они способствуют прикреплению растения к опоре.
Фотосинтез – это сложный многоступенчатый процесс превращения световой энергии в энергию химических связей органических молекул. При этом из окружающей среды для построения органических веществ поступают СО2, Н2О и минеральные соли, а выделяется О2
Если вам нужна помощь в написании работы, то рекомендуем обратиться к профессионалам. Более 70 000 авторов готовы помочь вам прямо сейчас. Бесплатные корректировки и доработки. Узнайте стоимость своей работы.
6СО2 + 12 Н2О → квант света, хлорофилл → С6Н12О6 + 6О2 + 6 Н2О
Для растений, в отличие от животных, свойственно два способа питания: воздушное питание и почвенное питание.
Кроме углеводов в процессе фотосинтеза образуются белки и сложные липиды, для этого необходимо наличие минеральных анионов (нитрат, нитрит, аммоний, сульфат, сульфит, фосфат), катионов металлов и аммиака. Процессы фотосинтеза и минерального питания тесно связаны между собой и обеспечивают растению способность синтезировать органические вещества из простых веществ с использованием солнечного света в качестве источника энергии.
Каждая фотосистема содержит 250-400 молекул пигментов. Все пигменты фотосистемы могут поглощать кванты света, но только одна молекула хлорофилла данной фотосистемы может использовать поглощённую энергию в фотохимических реакциях. Эта молекула называется пигментом – ловушкой, или реакционным центром фотосистемы. Другие молекулы пигментов называются антенными, поскольку они улавливают энергию света, подобно антеннам, для дальнейшей передачи её реакционному центру.
Выделяют два вида фотосистем: фотосистема – 1 и фотосистема – 2.
Фотосистемы состоят из следующих компонентов:
Фотосистемы 1 и 2 обычно работают синхронно и непрерывно, но фотосистема– 1 может функционировать отдельно.
3. совокупность белков – ферментов, которые активизируют окислительно-восстановительные превращения.
Фотофизический этап начинается с поглощения кванта света молекулами фотосинтеза. Хлорофиллы поглощают красные и сине-фиолетовые лучи, каротиноиды – сине-фиолетовые лучи, а фикобелины – жёлто-зелёные.
Молекулы пигментов, поглощая квант света, переходят в возбуждённое состояние, при этом энергия света переходит в энергию возбуждённых молекул хлорофилла. Молекулы хлорофилла передают энергия возбуждения пигмент-ловушкам реакционного центра.
Молекулы РЦ участвуют не только в фотофизических, но и в фотохимических превращениях.
Фотохимические превращения заключаются в том, что возбуждённые пигмент- ловушки реакционных центров отдают электроны в цепь белков – переносчиков (электронотранспортная цепь, или ЭТЦ). В результате движения электронов по ЭТЦ происходят реакции окисления и восстановления. Таким образом, в ходе фотохимических превращений энергия возбуждения молекул пигмент-ловушки превращается в энергию окисления–восстановления. Энергия окисления–восстановления (падающая энергия) превращается в энергию макроэргических связей АТФ, благодаря сопряжению реакций окисления, восстановления и фотофосфорилирования. Фотофосфорилирование на мембранах тилакоидов хлоропластов осуществляется двумя путями: циклическим и нециклическим.
Циклическое фосфорилирование происходит с участием только фотосистемы – 1. При этом донором и акцептором пары электронов является пигмент ловушка Р700. Энергия, выделяемая в процессе движения электронов, используется для синтеза АТФ. АТФ – это единственный продукт циклического транспорта электронов.
Нециклическое фосфорилирование происходит при согласованном взаимодействие двух фотосистем. Конечным акцептором электронов в фотосистеме – 1 является окисленная форма НАДФ → НАДФ+. В результате работы фотосистемы НАДФ+ переходит в восстановленную форму НАДФ.Н+.
НАДФ+ + 2е– + 2Н+ → НАДФ.Н+
При работе фотосистемы – 2 осуществляется синтез АТФ и конечным акцептором электронов является Р700. Работа фотосистемы – 2 активирует такой процесс как фотолиз воды. В результате выделяется побочный продукт – кислород.
Н2О ® ОН- + Н+ , 2ОН- ® Н2О2 , Н2О2 ® Н2О + О2
Электроны поступают на Р680, а протоны идут на восстановление НАДФ.
Итак, в световую фазу образуются два энергоёмких вещества: АТФ и НАДФН+. Накопленная в них энергия будет использоваться в темновую фазу фотосинтеза.
Темновая фаза фотосинтеза протекает в строме хлоропласта и представляет собой ряд биохимических превращений, в ходе которых СО2 восстанавливается до углеводов, белков и жиров. Темновая фаза может осуществляться как на свету, так и в темноте, но в темноте до тех пор, пока есть продукты световой фазы.
Темновые реакции могут осуществляться путём цикла Кальвина (С3 путь темновой фиксации углерода), путём Хетча-Слэка (С4 путь темновой фиксации углерода), САМ-метаболизм (темновая фаза по типу толстянковых).
Цикл Кальвина встречается у всех высших растений и включает в себя три последовательных этапа, объединённых в циклический процесс.
1. Карбоксилирование. На этом этапе взаимодействуют рибулозодифосфат, СО2 и вода с образованием фосфоглицериновой кислоты (ФГК). Т. к. молекула ФГК содержит три атома углерода, цикл Кальвина получил название С3 пути, а растения с таким типом фотосинтеза называются С3 растения.
2. Восстановление. Этот этап включает две реакции. В течение первой реакции происходит перенос остатка фосфорной кислоты с АТФ на ФГК (реакция фосфорилирования). В течение второй реакции происходит собственно восстановление с участием НАДФ.Н+. В результате образуется фосфоглицериновый альдегид (ФГА). Уровень восстановления ФГА соответствует уровню восстановления углеводов.
3. Регенерация. Этот этап представляет собой разветвлённую цепь биохимических реакций с образованием ряда промежуточных продуктов. Эти промежуточные продукты имеют большое значение для пластического обмена растений и частично могут выводиться из цикла Кальвина. На этом этапе возобновляются молекулы первичного акцептора (РДФ), которые вновь используются на первом этапе (карбоксилирования).
Первый продукт фотосинтеза фруктоза. Затем она может изомеризоваться в глюкозу, а она полимеризуется с образованием первичного крахмала.
Регенерация – это многоступенчатый процесс, протекающий с большим количеством промежуточных продуктов. В ходе реакции регенерации судьба шести молекул ФГА не одинакова. Пять молекул ФГА используется на регенерацию трёх молекул РДФ, а одна молекула ФГА уходит на образование фруктозы. Для образования одной молекулы фруктозы надо пройти два цикла Кальвина.
У некоторых растений на базе цикла Кальвина сформировались другие типы фиксации СО2. С4 –путь происходит у многих растений засушливых субтропиков и тропиков. У культурных растений С4 –путь наблюдается у кукурузы, сорго, сахарного тростника. Этот путь фотосинтеза является адаптивным и обеспечивает быстрое образование органических веществ в условиях засушливого климата. У многих растений из семейства толстянковые (кактус, алоэ, каланхоэ) темновые реакции осуществляются по пути САМ-метаболизма. Для них характерно невысокая производительность фотосинтеза в сочетании с высокой засухо – и жароустойчивостью.
Лист является важным органом любого растения. Основные функции листа — фотосинтез и транспирация. Строение листа характеризуется наличием черешка и листовой пластинки. Внешне черешок похож на стебель, однако по происхождению он все же является частью листа.
Лист по строению предполагает наличие кожицы, которой покрыта поверхность любого листа. Кожица является защитой от различных повреждений, высыхания и попадания внутрь болезнетворных бактерий.
Строение кожицы листа характеризуется тем, что ее клетки плотно примыкают друг к другу: это объясняется тем, что они являются покрывной тканью. Почти все клетки в листах не имеют цвета и прозрачные, поэтому свет без проблем проникает через поверхность листка в клетку. Как видим, строение листьев и строение клетки листа напрямую связаны с функциями листьев и формируют их особенности.
Начинают изучать клеточное строение листа в 6 классе школы.Контент.
Характеристика эпидермиса
Эпидермис — это то, чем лист покрыт снаружи.
Эпидермис является живой тканью листа и может состоять из одного или нескольких слоев клеток.
Такие клетки листа обычно не отличаются хорошо дифференцированными хлоропластами. Клетки соединены между собой достаточно плотно, благодаря чему эпидермис защищает ткани листа от чрезмерной потери воды и играет важную роль в осуществлении листом функции механической опоры.
Эпидермис имеет особенность в виде различных выростов на внешней поверхности клеток: волосков, кутикул, шипиков.
Также стоит упомянуть устьица листа, которые находятся между клетками эпидермиса. Основная функция устьиц — осуществление водо- и газообмена растения с окружающей средой. Эта функция выполняется, в том числе, за счет особенностей строения устьица листа.
Характеристика мезофилла
Мезофилл — основная ткань, которая размещается между верхним и нижним эпидермисом.
Она представляет собой фотосинтезирующую ткань: в нее входят живые клетки с большим количеством хлоропластов.
Мезофилл делится на губчатую и палисадную паренхиму. Последняя включает клетки, расположенные перпендикулярно к поверхности эпидермиса — они напоминают ряд столбиков (столбчатая паренхима). У клеток палисадной паренхимы призматическая форма, эти клетки удлинены. Расположение палисадной паренхимы — под эпидермисом. При этом у одних растений она располагается только в верхней стороне листа, а у других — с обеих сторон.
Разделение или дифференциация мезофилла основана на виде растения и специфике его выращивания. При ярком освещении хорошее развитие получает палисадная паренхима.
Злаковые умеренной зоны не имеют деления на палисадную и губчатую паренхимы.
Эти две ткани устроены по-разному, так как они отвечают за разные функции. И здесь мы найдем ответ на вопрос, как строение листа обеспечивает его фотосинтезирующие функции.
Палисадная паренхима является высокоспециализированной тканью и выполняет функцию фотосинтеза. Это логично, ведь большинство хлоропластов располагаются именно в этой ткани и концентрируются около стенок клетки — так они лучше освещаются и снабжаются углекислым газом.
Губчатая паренхима помимо функции фотосинтеза (хоть и в меньшей степени) выполняет запасающую функцию: в клетках листа скапливается запасной крахмал.
Характеристика проводящей ткани
Проводящая ткань листа включает сосудисто-волокнистые пучки: они сконцентрированы в жилках. По этим пучкам в лист попадает вода, насыщенная питательными веществами, и отводятся продукты фотосинтеза.
Проводящая ткань пластинки и черешка листа и проводящая система стебля образуют единое целое. Строение жилки листа может характеризоваться как одним пучком, так и целой группой пучков, тесно между собою сомкнутых.
Сосудисто-волокнистые пучки основных жилок листа отличаются типичным строением. По мере раздробления пучков сосуды и ситовидные трубки уменьшаются. В едва заметных разветвлениях жилок нет флоэмы. Ксилема также упрощается: в ней отсутствует трахея, сокращается количество трахеид. На концах жилок — одиночные трахеиды.
То, насколько крепкая листовая пластинка, зависит от развития системы механических тканей. В нее входят:
- склеренхимные обкладки пучков;
- тяжи механической ткани. Они размещаются против проводящих пучков и смыкаются позади склеренхимных обкладок;
- каменистые клетки;
- опорные клетки и др.
Функции устьица и его строение
Устьице по форме напоминает щель, которая располагается между двумя клетками со специфическим строением.
Эти клетки серповидные, между собой они смыкаются противоположными концами (замыкающие клетки). Они существенно отличаются от других клеток эпидермиса: по форме и наличию хлоропластов.
Устьица располагаются с нижней части листовой пластинки. Однако есть растения, у которых оно расположено в верхней части (злаки, капуста).
Устьица водных растений располагаются только в верхней стороне пластинки.
Число устьиц на листьях растений варьируется от 40 до 600 (на один квадратный миллиметр).
Листья с параллельным жилкованием (такие есть у хвойных растений) размещаются параллельными рядами. У других растений какого-либо конкретного порядка нет.
Устьица открываются по разным причинам:
- для осуществления газообмена;
- для фотосинтеза и дыхания листа;
- для контроля над водным балансом.
То, как осуществляется устьичное движение, определяется особенностями структуры замыкающих клеток, а также изменениями их тургорного давления. Неравномерное утолщение оболочек — отличительная характеристика строения замыкающих клеток устьиц. Это приводит к тому, что задняя стенка замыкающей клетки с увеличением тургора выпячивается в сторону щели, поскольку эта стенка отличается большей эластичностью и небольшой толщиной. При этом передняя стенка выпрямляется и становится вогнутой, а вся клетка изгибается в противоположную от щели сторону. Происходит открытие устьица.
Тургорное давление замыкающих клеток меняется в связи с большими затратами энергии. Регуляция осмотического давления замыкающих клеток осуществляется при помощи органических кислот, одновалентных катионов, в частности — калия.
Когда одновалентные катионы поступают в вакуоль замыкающих клеток, то осмотический потенциал последних увеличивается. В эти клетки поступает вода, и устьице открывается. Снижение осмотического давления происходит в результате выхода осмотических активных веществ из вакуолей в цитоплазму замыкающих клеток или из вообще из клетки. Устьице закрывается.
Поддержание электронейтральности замыкающих клеток при открытых устьицах обеспечивается образованием органических анионов.
Процесс поступления воды в клетку
Поступление воды в клетку — непростой процесс, который обусловлен множеством факторов.
Вся система коллоидов цитоплазмы принимает активное участие в поглощении воды.
Сосущая сила — сила насасывания клеткой воды.
Есть опыт, который помогает понять, как происходит поступление воды в живую клетку, а также показывает полупроницаемость и эластичность цитоплазмы.
К оборотной стороне покровного стекла, вплотную к нему, подносят фильтровальную бумагу: она оттягивает воду до того момента, пока раствор селитры полностью ее не заменит, входя под покровное стекло.
Спустя определенное время даже при небольшом увеличении микроскопа можно обнаружить отхождение протопласта от оболочки клетки. Такой процесс называется плазмолизом.
Далее протопласт округляется и размещается в середине клетки или возле одной из ее стенок. Происходит это после его отделения от всей внутренней поверхности оболочки. В результате происходит заполнение пространства между протопластом и оболочками клетки раствором плазмолитика.
Как клетка листа испаряет воду
Транспирация — испарение воды растениями.
Воду испаряет вся поверхность растения, но особенно интенсивно — лист.
Есть два вида транспирации:
- Кутикулярная. В этом случае воду испаряет вся поверхность листа.
- Устьичная. Испарение осуществляется через устьице листа.
Транспирация важна тем, что благодаря ей внутрь листа поступает углекислый газ, а это — основа углеродного питания растения. Кроме того, благодаря транспирации лист не перегревается.
Читайте также: