
Синтез генетики и дарвинизма кратко
Обновлено: 07.07.2024
Проведенные в 1928–1930 гг. исследования (Р.А. Фишер, Н.П. Дубинин и Д.Д. Ромашов, С. Раит и др.) показали, что в эволюции большую роль играет не только появление новых мутаций, но и изменение частоты встречаемости существующих аллелей (гена) благодаря случайным процессам – колебания численности популяции, утраты генов и т.д.
Генетика позволила проанализировать основные моменты протекания эволюционного процесса от появления нового признака в популяции до возникновения нового вида. На внутривидовом (микроэволюционном) уровне при изучении эволюции оказалось возможным применить точные экспериментальные подходы, которые помогли выяснить роль отдельных эволюционных факторов, сформулировать представления об элементарной эволюционной единице (популяция), элементарном эволюционном материале и явлении. Все это привело к созданию Ф.Г. Добржанским и Н.В. Тимофеевым-Ресовским в 1937–1939 гг. учения о микроэволюции – одного из главных разделов современной теории эволюции.
Хромосомная теория наследственности не снимала противоречий между дарвинизмом и генетикой. (Собственно говоря, такая задача перед ней и не стояла.) Важнейшим шагом на пути их преодоления явилось создание синтетической теории эволюции – первого глубокого синтеза классического дарвинизма, генетики, систематики, палеонтологии, экологии. Он привел к качественному скачку в развитии биологического знания, к переходу биологии с классического на неклассический уровень познания.
Принципиальные и концептуальные положения синтетической эволюции были заложены трудами С.С. Четверикова (1926), Р.Фишера, Н.В. Тимофеева-Ресовского, С. Райта, Н.П. Дубинина, Дж. Холдейна (1929–1932) и др. Свое развитие она получила в трудах таких выдающихся биологов XX в., как Н.И. Вавилов, И.И. Шмальгаузен, Э. Майр, Дж. Симпсон, Дж. Хаксли, Ф.Г.Добжанский и др. Непосредственными предпосылками создания теории выступали: хромосомная теория наследственности; традиция разработки биометрических и математических подходов в решении проблем генетики и эволюции, в частности закон Харди–Вайнберга (1908) для идеальной популяции (гласящий, что такая популяция – совокупность особей одного вида, населяющих определенную территорию, более или менее изолированную от соседних совокупностей того же вида – стремится сохранить равновесие концентрации генов при отсутствии факторов, изменяющих его), результаты эмпирических исследований изменчивости в природных популяциях и т.д.
· элементарный эволюционный материал – это мутации (мелкие дискретные изменения наследственности), обычно случайно образующиеся. В настоящее время выделяют генные, хромосомные, геномные (изменения числа хромосом и др.), изменения внеядерных ДНК и др.;
· наследственное изменение популяции в каком-либо определенном направлении осуществляется под воздействием элементарных эволюционных факторов, таких как: мутационный процесс, поставляющий элементарный эволюционный материал; популяционные волны (колебания численности популяции в ту или иную строну от средней численности входящих в нее особей); изоляция (закрепляющая различия в наборе генотипов и способствующая делению исходной популяции на несколько новых, самостоятельных популяций); естественный отбор.

Естественный отбор – ведущий эволюционный фактор; именно он направляет эволюционный процесс. Отбор действует на всех стадиях онтогенеза особей данного вида. Существуют разные формы естественного отбора: движущий – благоприятствующий лишь одному направлению изменчивости, когда происходит дивергенция дочерних форм; дизруптивный – разрывающий, благоприятствующий двум или нескольким направлениям изменчивости; стабилизирующий – благоприятствующий сохранению в популяции оптимального фенотипа и действующий против проявлений изменчивости.
Хромосомная теория наследственности не снимала противоречий между дарвинизмом и генетикой. (Собственно говоря, такая задача перед ней и не стояла.) Важнейшим шагом на пути их преодоления явилось создание синтетической теории эволюции – первого глубокого синтеза классического дарвинизма, генетики, систематики, палеонтологии, экологии. Он привел к качественному скачку в развитии биологического знания, к переходу биологии с классического на неклассический уровень познания.
Принципиальные и концептуальные положения синтетической эволюции были заложены трудами С.С. Четверикова (1926), Р.Фишера, Н.В. Тимофеева-Ресовского, С. Райта, Н.П. Дубинина, Дж. Холдейна (1929–1932) и др. Свое развитие она получила в трудах таких выдающихся биологов XX в., как Н.И. Вавилов, И.И. Шмальгаузен, Э. Майр, Дж. Симпсон, Дж. Хаксли, Ф.Г.Добжанский и др. Непосредственными предпосылками создания теории выступали: хромосомная теория наследственности; традиция разработки биометрических и математических подходов в решении проблем генетики и эволюции, в частности закон Харди–Вайнберга (1908) для идеальной популяции (гласящий, что такая популяция – совокупность особей одного вида, населяющих определенную территорию, более или менее изолированную от соседних совокупностей того же вида – стремится сохранить равновесие концентрации генов при отсутствии факторов, изменяющих его), результаты эмпирических исследований изменчивости в природных популяциях и т.д.
· элементарный эволюционный материал – это мутации (мелкие дискретные изменения наследственности), обычно случайно образующиеся. В настоящее время выделяют генные, хромосомные, геномные (изменения числа хромосом и др.), изменения внеядерных ДНК и др.;
· наследственное изменение популяции в каком-либо определенном направлении осуществляется под воздействием элементарных эволюционных факторов, таких как: мутационный процесс, поставляющий элементарный эволюционный материал; популяционные волны (колебания численности популяции в ту или иную строну от средней численности входящих в нее особей); изоляция (закрепляющая различия в наборе генотипов и способствующая делению исходной популяции на несколько новых, самостоятельных популяций); естественный отбор.
Естественный отбор – ведущий эволюционный фактор; именно он направляет эволюционный процесс. Отбор действует на всех стадиях онтогенеза особей данного вида. Существуют разные формы естественного отбора: движущий – благоприятствующий лишь одному направлению изменчивости, когда происходит дивергенция дочерних форм; дизруптивный – разрывающий, благоприятствующий двум или нескольким направлениям изменчивости; стабилизирующий – благоприятствующий сохранению в популяции оптимального фенотипа и действующий против проявлений изменчивости.
Генетика изучает один из факторов эволюции - наследственную изменчивость, поэтому противопоставление генетики дарвинизму было искусственным. Открытие мутаций свидетельствовало о наличии материала для естественного отбора, а выявление материальных единиц наследственности - генов - доказывало возможность закрепления результатов действия отбора, сохранение и накопление полезных в данной среде мелких наследственных изменений.
Один из первых значительных шагов по объединению данных генетики и дарвинизма сделан отечественной наукой. Русский биолог С.С. Четвериков (1880-1959) впервые подвел генетическую основу под эволюционное учение Дарвина, в работе "О некоторых моментах эволюционного процесса с точки зрения современной генетики" (1926) показано, что в естественных условиях в природе внутри каждого вида существует огромное количество наследственных изменений, которые фенотипически не проявляются в силу рецессивности. Вид насыщен мутациями, составляющими неисчерпаемый материал для эволюции.
Итогом работы С.С. Четверикова и его последователей - советских ученых Н.В. Тимофеева-Ресовского, Н.К. Беляева, Б.Л. Астаурова, Н.П. Дубинина, Д.Д. Ромашова, английского исследователя Р.А. Фишера, американских ученых С. Райта и Ф. Г. Добржанского и многих других - явилось возникновение новой дисциплины - популяционной генетики.
Развитию популяционной генетики способствовал обмен многими идеями и методами между ведущими тогда генетическими школами - американской и советской. Многие работы советских генетиков печатались на европейских языках, среди них следует назвать Н.И. Вавилова, Н.В. Тимофеева- Ресовского, Г.К. Карпеченко, Н.К. Кольцова и др.

Рис. 181. Николай Константинович Кольцов.
Из: В. Полынин, 1969.

Рис. 182. Сергей Сергеевич, Четвериков.
Фото из архива автора, 1969.
В 1921 г. в Советскую Россию приехал ближайший сотрудник Моргана, будущий лауреат Нобелевской премии Герман Мёллер (1890—1967, рис. 183), рассказавший Кольцову и Четверикову о работах школы Моргана по хромосомной теории наследственности, выполненных на разводимых в пробирках американских видах дрозофил. Энтомологу Четверикову стало ясно, что с не меньшим успехом можно работать и на европейских видах этого рода. В отличие от американских генетиков, Четвериков решил изучать природные популяции дрозофил, личинки которых питаются грибами[353].

Рис. 183. Герман (Германович) Мёллер (второй слева) — сотрудник Н. И. Вавилова, член-корреспондент АН СССР — и его ассистент К. Офферман (третий слева) с советскими генетиками академиком АН УССР И. И. Аголом (крайний слева) и основателем Медико-генетического института С. Г. Левитом. И. И. Агол и С. Г. Левит были расстреляны. Мёллер в 1937 г. уехал из СССР в республиканскую Испанию, а затем в США. Фото начала 1930-х годов.
Из: Baumunk und Rie? (1994).
1) мутационный процесс происходит в природных популяциях (что тогда было совсем не очевидно);
2) большинство вновь возникших мутаций понижает жизнеспособность, хотя изредка возникают и мутации, повышающие таковую;
5) по мере старения вида в нем накапливается все большее число мутаций, при этом признаки вида расшатываются;
6) генетическая изменчивость проявляется наиболее сильно, когда многочисленный вид распадается на ряд небольших, изолированных колоний;
9) естественный отбор является антагонистом свободного скрещивания — панмиксии;
10) приспособительная эволюция без изоляции ведет к полной трансформации вида, но не может привести к его распадению на два вида [Важнейшее заключение! Четвериков представлял возможность недивергентного, анагенетического (см. ниже) видообразования без изоляции и считал, что дивергентное видообразование, или кладогенез, невозможно только на основе отбора без изоляции];
11) отбор, как и свободное скрещивание, ведет к накоплению рецессивных аллелей в гетерозиготном состоянии;
12) свободное скрещивание — панмиксия — ведет к повышению полиморфизма вида, а отбор ведет к мономорфизму;
13) отбор отбирает не только отдельный ген, контролирующий отдельный признак, но и, благодаря множественности (плейотропносги) действия генов, влияет на все генное окружение отбираемого гена, на его генотипическую среду.

Рис. 184. Николай Владимиович Тимофеев-Ресовский. Фото Esko Suomalainen (1943).
Из архива автора.

Рис. 185. Елена Александровна Тимофеева-Ресовская (урожденная Фидлер).
Из: Н. В. Тимофеев-Ресовский (1995).

Рис. 186. Николай Константинович Беляев.
Из архива Н. Б. Астауровой.

Рис. 187.Борис Львович Астауров.
Фото С. И. Раджабли.

Рис. 189. Дмитрий Дмитриевич Ромашов.
Из архива Н. Б. Астауровой.

Рис. 188. Елизавета Ивановна Балкашина.
Из архива автора.

Рис. 190. Александр Николаевич Промптов.
Из: А. Н. Промптов (1956).

Рис. 191. Петр Фомич Рокицкий.
Из: Гершензон (1992).

Рис. 192. Сергей Михайлович Гершензон.
Из: Гершензон (1992).

Из: Развитие биологии в СССР (1967).
Действительно, несмотря на то, что работа Четверикова была опубликована по-русски, а публикации его учеников в отечественных журналах вышли с большим опозданием и в неполном виде, его идеи были быстро разнесены по свету. Н. В. Тимофеев-Ресовский неустанно пропагандировал идеи своего учителя среди европейских генетиков, а Ф. Г. Добржанский — среди американских. Следует также отметить, что оба эти исследователя были по своему изначальному образованию зоологами, и потому они смогли пропагандировать идеи Четверикова не только в генетических, но и в зоологических кругах.

Рис. 194. Феодосий Григорьевич Добржанский.
Из: Dobzhansky’s Genetics of Naturals Populations (1981).
Обмену многими идеями и методами между ведущими тогда генетическими школами — американской и советской — способствовало несколько обстоятельств: многие работы советских генетиков печатались на европейских языках, в 20-е годы многие советские генетики выезжали за рубеж на длительные сроки (рис. 196—198). Среди них были Н. И. Вавилов, Н. К. Кольцов, блестящий ученик Вавилова безвременно погибший в сталинских тюрьмах цитогенетик Георгий Дмитриевич Карпеченко (1899—1942), классик цитогенетики растений Михаил Сергеевич Навашин (1896—1973), генетик и селекционер растений Антон Романович Жебрак (1901—1965), а в конце 20 — начале 30-х гг. в СССР по приглашению Вавилова долгое время работали ученики Моргана — Мёллер (1890—1967), которого советские коллеги звали Германом Германовичем, будущий Нобелевский лауреат, избранный в те годы членом-корреспондентом АН СССР; Кэльвин Бриджес (1889-1938) — один из создателей хромосомной теории наследственности, а также ведущий специалист по генетике хлопчатника С. Е. Харланд.

Из архива автора.
Крупнейший английский генетик-эволюционист Джон Бердон Сандерсон Холдейн (1892-1964, рис. 199) встречался с Четвериковым на V Международном генетическом конгрессе в Берлине в 1927 г., в его лаборатории был осуществлен перевод четвериковской статьи 1926 г., в работах его ученицы Сесиль Гордон (1936, 1939) отмечалась инициирующая роль работы Четверикова в последующих исследованиях; этим переводом пользовались сотрудники и гости лаборатории Холдейна.

Из: В.-М. Baumunk und J. Rie? (1994).

Из: В.-М. Baumunk und J. Rie? (1994).

Рис. 198. Н. И. Вавилов, Т. Г. Морган и Н. В. Тимофеев-Ресовский на Международном генетическом конгрессе в Итаке (США, 1932). Н. И. Вавилов — вице-президент конгресса — был единственным приехавшим из СССР. Н. В. Тимофеев-Ресовский прибыл из Берлина.

Рис. 199. Джон Б. С. Холдейн.
Из: Genetics (1965).

Рис. 200. Рональд Эйлмер Фишер.
Из: R. W. Clark (1984).

Рис. 201. Сьюэлл Райт.
Из: Dobzhansky’s Genetics of Naturals Populations (1981).
Дальнейшее развитие теоретическая популяционная генетика получила в трудах Д. Д. Ромашова[367], Н. П. Дубинина[368], Тимофеева-Ресовского[369], Дж. Холдейна[370], А. А. Малиновского, С. Ли[371] . В 1935 г. в СССР Андрей Николаевич Колмогоров (1903—1987, рис. 202) математически показал, что наибольшие темпы изменения генотипического состава популяции характерны не для больших и не для малых популяции, а для серии полуизолятов с периодически возникающими и исчезающими потоками генов между ними [372].

Рис. 202. Андрей Николаевич Колмогоров.
Фото 1930-х гг. из архива автора.
Ленинградский генетик Юрий Александрович Филипченко (1882—1930) в 1927 г. предложил разделить эволюцию на два процесса — микроэволюцию, то есть дивергенцию от популяционного до видового уровня, и макроэволюцию, то есть дивергенцию на уровне выше вида[375]. По мнению Филипченко (в этом отношении он резко расходился как с рядом современных ему, так и с большинством последующих эволюционистов), роды произошли иным путем, чем виды. Он считал, что мутации, их комбинации и отбор могут объяснить лишь микроэволюцию[376]. Любопытно, что сейчас многие критики синтетической теории эволюции возвращаются к подобной позиции.

Рис. 203. Схема получения кариотипа нового синтезированного вида Crepis nova:
а — хромосомы Cr. tectorum; б, г — исходные гетерозиготные транслоканты; в, д — те же транслокацни, но переведенные в гомозиготное состояние; г — гибрид между в и д; ж-п — девять возможных комбинаций хромосом в потомстве е; п — Cr. nova.
Е. Н. Герасимовой из: Н. Н. Воронцов.
Идеи Четверикова роли явления рецессивности в эволюции с помощью Н. В. Тимофеева-Ресовского и Ф. Г. Добржанского распространились по мировому сообществу генетиков и эволюционистов и стали восприниматься как самоочевидные. В нашей стране на эти идеи особое внимание обратил Иван Иванович Шмальгаузен (1884—1963). В самом деле, ведь отбор идет по фенотипам, и, таким образом, особи с генотипами АА и Аа, имея одинаковый фенотип благодаря доминантности аллеля А над рецессивным аллелем а, обладают равной селективной ценностью перед лицом естественного отбора. Таким образом, под покровом доминантного аллеля А рецессивный аллель а может очень долго существовать в популяции даже в том случае, если гомозиготы по рецессивному аллелю аа имеют пониженную жизнеспособность (табл. 3).
Таблица 3. Темпы возрастания частоты доминантного аллеля А в популяции, испытывающей слабое действие отбора (особи с генотипами АА и Аа имеют 1%-ное селективное преимущество по сравнению с генотипом аа). По К. Пэтау, 1939


Рис. 204. Николай Иванович Вавилов.
Из: С. Резник (1968).

Рис. 205. Георгий Андреевич Левитский. Классик цитогенетики. Член-корреспондент АН СССР. Арестован в 1940 г. вместе с Н. И. Вавиловым. Погиб в лагерях.
Из: С. М. Гершензон (1992).

Рис. 206. Георгий Дмитриевич Карпеченко. Классик цитогенетики. Арестован в 1940 г. вместе с Н. И, Вавиловым. Погиб.
Из: С. М. Гершензон.

Рис. 208. Антон Романович Жебрак. Президент АН Белоруссии. В 1948 г. был снят со всех постов за критику Т. Д. Лысенко. В середине 50-х годов — заведующий кафедрой ботаники Московского фармацевтического института, где вместе с В. В. Сахаровым восстановил подготовку генетиков.
Из. Выдающиеся советские генетики (1980).

Рис. 207. Петр Михайлович Жуковский. Ботаник, генетик, эволюционист. Академик ВАСХНИЛ. Был директором вавиловского Всесоюзного института растениеводства. с 1948 г. был вынужден прекратить работы по генетике.
Из: Соратники Николая Ивановича Вавилова (1994).

Рис. 209. Сирилл Дарлингтон (1903-1981) — в центре, слева — Г. Г. Мёллер, справа — Н. В. Тимофеев-Ресовский. 1930-е гг.

Современная синтетическая теория эволюции, или СТЭ, — это объединение дарвинизма, генетики, молекулярной биологии и других наук. Синтетическая теория эволюции кратко и понятно описывает, как происходят эволюционные изменения живых организмов, какие признаки подвергаются отбору, какие факторы действуют на них и как это ведет к появлению новых видов.
В статье мы разбираем основные положения СТЭ, выясняем, почему Дарвин был прав (хотя учитывал не все) и как генетика и другие науки связаны с эволюцией.
История теорий эволюции
Эволюция – это естественный непрерывный процесс развития популяций живых организмов. Они все лучше и лучше приспосабливаются к окружающим условиям либо вымирают из-за более приспособленных конкурентов или резких изменений среды. Так появляются и исчезают виды.
- Разнообразие живых организмов: почему на Земле обитает столько разных живых существ, от бактерий до человека;
- Почему живые организмы обладают приспособлениями к условиям жизни и как они появились: зачем хищным животным клыки, а птицам – воздушные мешки;
- Почему организмы, обитающие в похожих условиях, сходны между собой: у рыб, морских котиков и жуков-плавунцов обтекаемая форма тела;
- Как живые организмы приспосабливаются к изменениям условий: почему рыба илистый прыгун умеет прыгать, у ящериц кожа покрыта чешуей и зачем кошке хвост.
На эти вопросы пытались отвечать разными способами. До появления теории эволюции Дарвина о развитии живых организмов задумывались еще в античности.
В средневековье основной теорией происхождения был креационизм. Однако уже в эпоху Возрождения с ним начали спорить ученые: наблюдения и расчеты слишком сильно противоречили богословию.
В Новое Время все больше ученых предполагало, что живые организмы меняются – эволюционируют. Однако ни причины, ни механизмы этого не были ясны.
Попытку объяснить эволюцию живых организмов сделал Жан Ламарк. Основные положения ламаркизма:
- Живые организмы развиваются от простого к сложному;
- Видов живых организмов не существует, важны только отдельные особи;
- Особи приспосабливаются к изменениям среды и конкурируют друг с другом;
- Механизм приспособления – тренировка наиболее используемых органов. Если органы не тренировать, они исчезают, как глаза крота.
Некоторые идеи Ламарка были верны. Однако его теория не выдержала экспериментальной проверки.
Август Вейсман отрезал хвосты нескольким поколениям мышей. Согласно ламаркизму, у последующих поколений хвосты должны были исчезнуть из-за нетренированности. Однако этого не произошло. Страдания мышей доказали, что приобретенные признаки не наследуются.
В действительности это верно для признаков, приобретенных не в результате изменения наследственной информации. Однако во времена Ламарка генетику еще не знали.
Ламаркизм критиковал и Жорж Кювье, основатель сравнительной анатомии, палеонтологии и теории катастрофизма. Согласно ей, каждый геологический период Земли заканчивался катастрофой, в результате чего изменялись флора и фауна. Теория Кювье не вошла в синтетическую теорию эволюции, поскольку она была тесно связана с религией. Однако она стала основой неокатастрофизма – гипотезы об одновременных изменениях рельефа Земли в результате тектонических катастроф.
С Кювье спорили сторонники трансформизма – идеи о том, что живые организмы происходят друг от друга. Подобные предположения высказывали в разные времена в разных странах. В XIX веке теорию трансформизма в разных вариантах защищали разные ученые. В частности, ее придерживался Ламарк. Частично идеи трансформизма присутствуют и в теории Дарвина.
Окаменевшие останки древнего гигантского животного среди раковин моллюсков современных видов навели его на идею, что вымирание не всегда – следствие катастроф. [1] Другие наблюдения – в частности, за галапагосскими вьюрками и черепахами – позволили Дарвину предположить, что разные виды могут происходить от общего предка. [1]
Вернувшись в Англию, Дарвин начал вести наблюдения за сельскохозяйственными растениями и животными. Они показывали необходимость приспосабливаться к определенным условиям. Дарвин назвал ее естественным отбором.
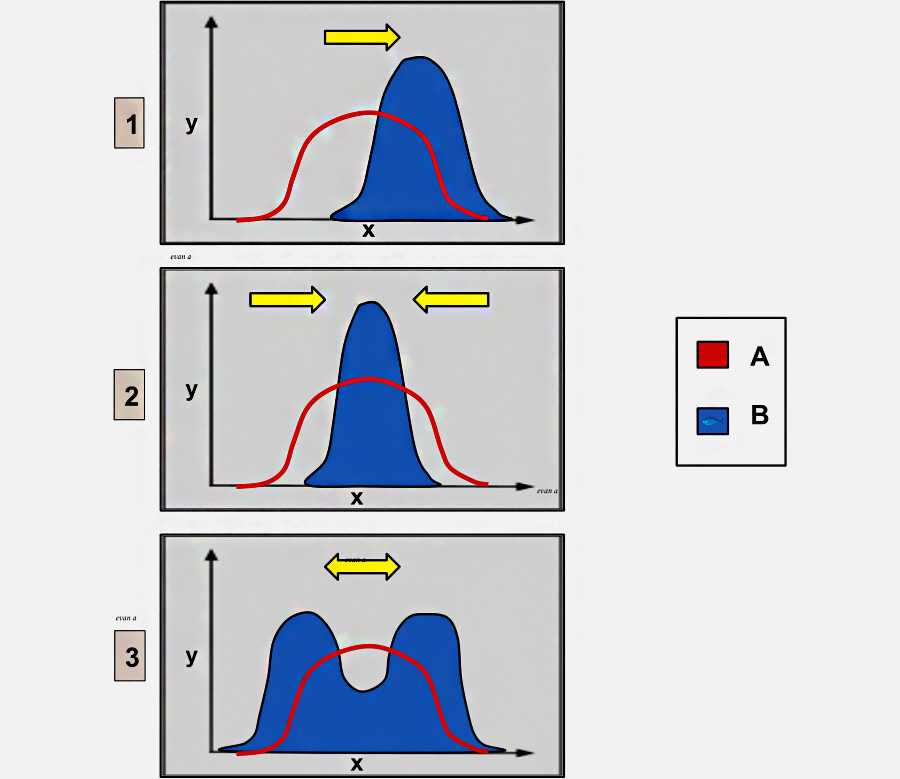
1 — направленный отбор: предпочтение отдается одному экстремальному фенотипу;
2 — стабилизирующий отбор: промежуточный фенотип предпочтительнее крайнего; 3 — разрушающий отбор: экстремальный фенотип предпочтительнее промежуточного.
Ось X: фенотипический признак
Ось Y: количество организмов
Группа А: исходная популяция
Группа B: после отбора

Слева — Чарльз Дарвин; справа — Альфред Уоллес. Интересный факт: Чарльз Дарвин, получив рукопись Альфреда Уоллеса о естественном отборе, пообещал использовать термины и наработки своего коллеги. 1 июля 1858 г. выдержки из трудов Дарвина и Уоллеса относительно естественного отбора были впервые представлены широкой публике — на чтениях в Линнеевском обществе.
Дарвинизм считают основой синтетической теории эволюции. Согласно ему, главные эволюционные факторы:
- Естественный отбор – выживают и оставляют потомство наиболее приспособленные особи;
- Борьба за существование – живые организмы конкурируют друг с другом за ресурсы;
- Наследственная изменчивость – живые организмы могут приобретать новые признаки, и, если они полезны, их передадут потомкам;
- Изоляция – невозможность межвидового скрещивания.
Теорию Дарвина подтверждают эмбриологические данные: эмбрионы позвоночных на ранних стадиях развития очень похожи. Различия проявляются потом. Это указывает, что позвоночные животные имеют общее происхождение. Однако ее критиковали современные ученому палеонтологи и некоторые натуралисты. Палеонтологи еще не успели обнаружить переходные формы живых организмов, существование которых предполагает дарвинизм.
Кроме того, теория Дарвина не объясняла, что именно является носителем наследственности и как происходит наследование. C этим связано одно из самых серьезных возражений против дарвинизма.
Почему кошмар Дженкина не проявляется в реальности, объясняет генетика. Причина – в дискретности наследования: наследуется не весь геном целиком, а его отдельные участки – гены. Поэтому даже если признак размыт во втором поколении, он проявится позднее. А для проявления доминантных признаков достаточно одной копии гена.
В трудах Чарльза Дарвина генетика не упоминается. Эту науку, основанную Грегором Менделем, оценили позднее. Даже в 20-30-х годах XX века она казалась многим не вполне научной.
Законы генетики, которые Мендель открыл, собирая и статистически анализируя данные, подтвердились после 1944 года, когда было доказано, что носителем наследственной информации является ДНК. Генетика и особенности передачи и изменения генов прояснили вопросы, на которые не мог ответить дарвинизм.
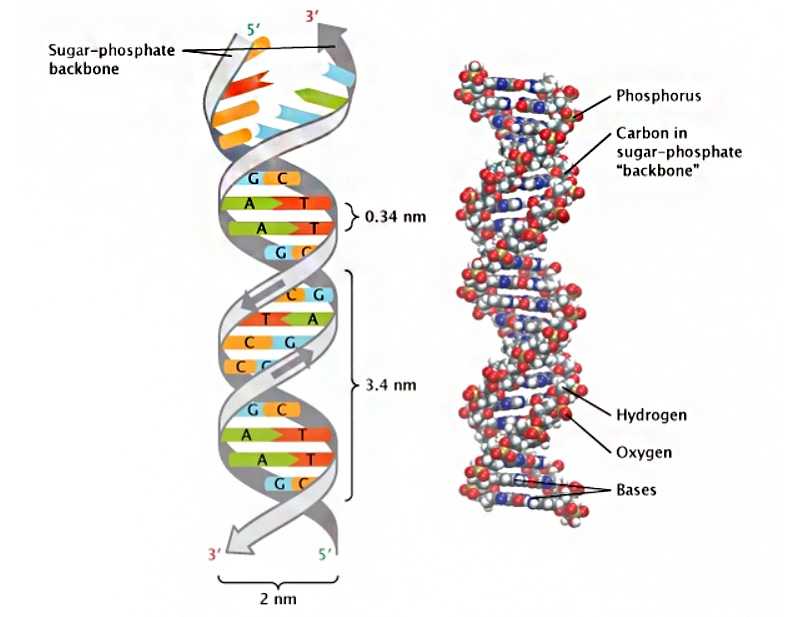
Трехмерная структура двойной спирали ДНК, открытая Джеймсом Уотсоном и Фрэнсисом Криком. Комплементарные основания удерживаются вместе с помощью водородных связей.
Синтетическая теория эволюции
Чтобы кратко и понятно объяснить процесс эволюции, потребовался синтез дарвинизма, генетики, палеонтологии и эмбриологии. Современная СТЭ кроме этих дисциплин включает данные молекулярной биологии и других наук. Естественно, такое преобразование теории Дарвина произошло не сразу.
В начале XX века в научном сообществе признавали идею эволюции, но отвергали естественный отбор как ее основную движущую силу. Во многом к этому привел кошмар Дженкина и недостаточные знания о наследственности.
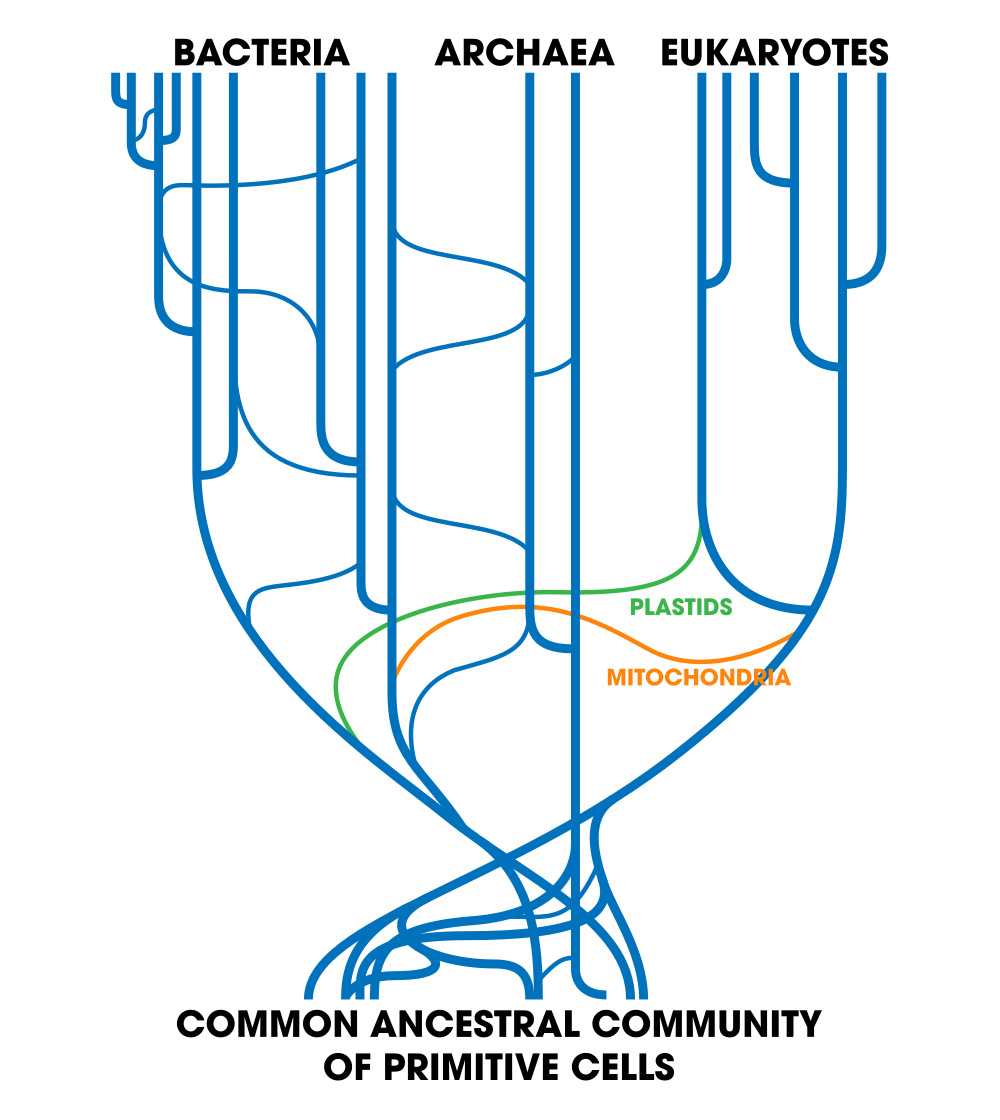
Древо жизни 21-го века, показывающее горизонтальный перенос генов среди прокариот и события сальтационного эндосимбиоза, породившего эукариот, не вписывается в теории 20-го века.
Только в 1926 году Сергей Сергеевич Четвериков опубликовал статью, в которой описал эволюцию с точки зрения современной ему генетики. Эта статья не была опубликована в англоязычных изданиях, однако ее цитировали Николай Тимофеев-Ресовский, Феодосий Добржанский и Джон Холдейн.
Идеи Четверикова хорошо соотнеслись с работами Рональда Фишера. Он создал математическую модель эволюции как естественного отбора генов. Единицей эволюции при этом оказалась популяция, причем эволюция шла тем быстрее, чем больше генетическое разнообразие в популяции. Модель Рональда Фишера стала одной из основ синтетической теории эволюции.
- Генные и хромосомные мутации;
- Изменения в популяции, вызванные изменениями частот и комбинаций мутаций;
- Закрепление изменений путем репродуктивной изоляции – невозможности скрещиваться с другими видами.
Считается, что синтетическая теория эволюции возникла именно в 1937 году. Сам термин СТЭ появился позднее – в 1942 году [3]. Его впервые использовал эволюционист Джулиан Хаксли, родной брат писателя Олдоса Хаксли.
Современная синтетическая теория эволюции учитывает особенности наследования, мутации и то, как под их действием меняется приспособленность живых организмов к окружающей среде. Наконец, СТЭ учитывает влияние различных факторов на эволюцию:
- Естественный отбор – важнейший, но не единственный фактор эволюции согласно СТЭ. Отбору подвергаются генотипы в целом либо аллели генов – то, в каких вариантах существует ген.
- Борьба за существование – необходимость выживать в окружающей среде и конкурировать за выживание с другими организмами.
- Возникновение мутаций приводит к появлению новых аллелей или новых генов. Мутации поставляют материал для естественного отбора.
- Изменения частот генов в популяции как результат миграций (поток генов), случайностей (дрейф генов) или эффекта бутылочного горлышка – ситуации, когда условия меняются так сильно и резко, что выживает лишь небольшое количество организмов из популяции. Зачастую у них есть признаки, позволившие им выжить именно в условиях таких изменений.
- Стабилизирующий отбор поддерживает соотношение частот генов в популяции неизменным, если окружающие условия не меняются [5];
- Изоляция – географические, биохимические или поведенческие препятствия для скрещивания между популяциями. Именно изоляция приводит к образованию новых видов живых организмов.
Синтетическая теория эволюции – не единственная современная теория эволюции. Остальные научные теории предполагают действие других факторов эволюции либо иную оценку их влияния. Однако СТЭ достаточно хорошо объясняет, как появляются новые виды и что именно подвергается отбору. Уточнения теории необходимы в основном для организмов, не имеющих полового процесса.
Основные положения синтетической теории эволюции
Синтетическую теорию эволюции проще всего объяснить, используя ее основные положения. Это закономерности, из которых следует, как и почему происходит эволюция живых организмов и на что она направлена.
- Единица эволюции – популяция, то есть, группа живых организмов одного вида, живущая на одной ограниченной территории и способная свободно скрещиваться друг с другом. Именно из популяций образуются новые виды.
- Материал эволюции – гены и их сочетания (генотипы). Новые гены и их сочетания возникают в результате случайных процессов – мутаций и рекомбинаций.
- Главный движущий фактор эволюции – естественный отбор. Другие процессы, которые приводят к изменению частоты встречаемости генов в популяции, также являются движущими факторами эволюции. Скорость и направление эволюции определяется сочетанием движущих факторов.
- Эволюция не имеет конечной цели. Это приспособление именно к конкретным условиям обитания. Если условия изменятся, к ним придется приспосабливаться заново.
- Изменения генов накапливаются в поколениях. Постепенно они приводят к образованию новых видов, родов и т.д.
Разные источники по-разному формулируют основные положения синтетической теории эволюции. Но все они сходятся в главном: существующие виды дают начало новым в результате приспособления к окружающей среде и отбора новых вариантов генов.
Мифы о синтетической теории эволюции
Суть синтетической теории эволюции, как и ее основные положения, не всегда понимают верно. Это приводит к различным заблуждениям: от смешивания СТЭ с дарвинизмом или ламаркизмом до попыток объяснять с ее помощью небиологические явления. Разберем несколько самых распространенных мифов об СТЭ.
Миф: Теория эволюции описывает появление жизни
Факт: происхождение жизни описывает абиогенез. Теория эволюции описывает, как живые организмы приспосабливаются к условиям обитания.
Миф: Естественный отбор предполагает, что выживает всегда сильнейший
Факт: Естественный отбор – это сохранение генотипов или аллелей генов, которые обеспечивают наилучшее приспособление к конкретным условиям. Чтобы сохранить их, желательно выжить самому и оставить потомство, которое также успешно размножится. Другой вариант – не размножаться, а способствовать выживанию носителей тех же генов. [6]
Миф: Синтетическая теория эволюции – это теория, а значит, она не доказана.
Факт: В науке теорией называют непротиворечивую концепцию, которая описывает и объединяет закономерности, касающиеся определенного явления. Научные теории позволяют предсказывать результаты тех или иных событий, которые подчиняются описанным закономерностям. СТЭ соответствует этому определению.
Миф: Концепция эволюции лженаучна, ее невозможно подтвердить экспериментально.
Факт: Эволюцию подтверждают палеонтологические и эмбриологические данные, а также результаты исследования ДНК и РНК различных организмов, полученные методами молекулярной биологии. Кроме того, примером эволюции является развитие у бактерий устойчивости к антибиотикам – этот процесс достаточно быстрый, и его можно наблюдать.
Миф: Без направленной осознанной воли сложные организмы не могли возникнуть.
Факт: Данное утверждение не является научным и не предполагает научного подхода к ответу. Результаты исследований в области молекулярной биологии, палеонтологии, эмбриологии показывают, что сложные организмы возникли в результате действия эволюционных закономерностей на случайно возникающие новые варианты генов.
Миф: Эволюция не объясняет появление разума.
Факт: Разум – один из способов приспосабливаться к окружающей среде. Как и другие эволюционные приспособления, он отчасти развился под действием закономерностей, а отчасти – благодаря случайностям.
Миф: Теория эволюции – основа социального дарвинизма
Факт: Распространять биологические закономерности на человеческое общество неверно с точки зрения науки. Общества развиваются по иным законам, и взаимная помощь и поддержка необходимы для существования цивилизации.
Большинство противников синтетической теории эволюции не имеют отношения к науке и плохо разбираются в ней.
Но и сторонники СТЭ не всегда хорошо знают ее. Разобраться в процессах эволюции помогут научно-популярные книги, фильмы, сериалы и лекции.
Интересные доклады и презентации о СТЭ
Тем, кто интересуется биологией вообще и синтетической теорией эволюции в частности рекомендуем лекции Станислава Дробышевского. Помимо антропогенеза он рассказывает и о теории эволюции. Юмор Дробышевского известен его постоянным слушателям, которые регулярно цитируют самые удачные высказывания в комментариях к видео.
Кроме научно-популярных лекций рекомендуем фильмы и книги о синтетической теории эволюции и сопутствующих дисциплинах.
Заключение
Фильмы и сериалы, которые стоит посмотреть
Книги, которые стоит прочитать
Синтетическая теория эволюции — ответы на популярные вопросы
Основной движущей силой эволюции СТЭ называет естественный отбор. Он действует на генотипы либо на аллели генов. Кроме того, движущими силами эволюции являются борьба за существование, возникновение мутаций, поток генов (обмен между разными популяциями), дрейф генов (случайные изменения частот аллелей) и изоляция (препятствие потоку генов).
В целом – нет. Чарльз Дарвин не мог знать о биологических открытиях XX и XIX веков. Поэтому в его работах есть устаревшие предположения. Однако основные принципы эволюции Дарвин вывел верно. Синтетическая теория эволюции основана на теории Дарвина и дополняет ее, а не опровергает.
Наличием общего предка с такими же чертами (параллелизм) либо сходством условий обитания (конвергенция). Если же организмы не близкородственные и развивались в разных условиях, их сходство объясняется похожими мутациями. Совпадение мутаций – достаточно вероятное событие: некоторые участки ДНК мутируют активнее других, а набор мутаций хотя велик, но все-таки конечен.
Если Ламарк был неправ, почему у более высоких и мускулистых людей обычно и дети более высокие и мускулистые?
Нет. Второе начало термодинамики гласит, что энтропия не убывает при усложнении замкнутых систем. Живые организмы – это открытые системы, поэтому они могут усложняться, и их энтропия при этом не увеличивается.
Потому что они не влияют на приспособленность организмов. У таких признаков может измениться функция. Это произошло, например, с аппендиксом человека: он не участвует в пищеварении, но необходим для нормальной работы иммунной системы и поддержания микрофлоры кишечника. Иногда признак может быть утрачен – как глаза у некоторых кротов. Зрение не нужно им для выживания и никак не влияет на приспособленность к среде. Поэтому мутации, затрагивающие зрение, кротам не вредят. Они накапливаются в популяции и постепенно приводят к утрате глаз.
Источники
Читайте также: