
Симбиотическая теория происхождения эукариотических клеток кратко
Обновлено: 05.07.2024
Симбиотическая гипотеза в последнее время находит большое количество сторонников и часто называется уже не гипотезой, требующей доказательств, а теорией.
Симбиотическая, или эндосимбиотическая, гипотеза исходит из того, что эукариоты представляют собой результат симбиоза между различными прокариотами. Допускается, что ядро развилось вследствие обособления ДНК из цитоплазмы путём впячивания плазматической мембраны и обволакивания ядерного вещества. Сходным образом могли возникнуть и эндоплазматическая сеть, и аппарат Гольджи.
Одновременно с этими процессами или несколько позднее таким же путём мог произойти и эндоцитозный захват автотрофных прокариот, например сине-зелёных водорослей (цианобактерий), обладающих полным набором пигментов фотосинтеза. Из этих симбионтов развились хлоропласты. Допускается также, что некоторые сложные по строению клетки, например простейших, произошли в результате симбиотических связей не только с прокариотами, но и с другими эукариотами.
Симбиотическая гипотеза в настоящее время находит подтверждение рядом эволюционных реликтов в существующих и сейчас симбиотических связях между прокариотами и эукариотами. Такие связи наблюдаются среди растительных и животных организмов. Например, свободноживущая амёба Pelomyxa palustris (пеломикса болотная) не имеет митохондрий, но содержит в себе симбиотические бактерии, которые обеспечивают ей дыхание и АТФ.
Расцвет эукариот на Земле начался около 1 млрд лет назад, хотя первые из них появились намного раньше (возможно 2,5 млрд лет назад). Происхождение эукариот могло быть связано с вынужденной эволюцией прокариотических организмов в атмосфере, которая стала содержать кислород.
Симбиогенез — основная гипотеза происхождения эукариот
Существует несколько гипотез о путях возникновения эукариотических клеток. Наиболее популярная — симбиотическая гипотеза (симбиогенез). Согласно ей, эукариоты произошли в результате объединения в одной клетке разных прокариот, которые сначала вступили в симбиоз, а затем, все более специализируясь, стали органоидами единого организма-клетки. Как минимум симбиотическое происхождение имеют митохондрии и хлоропласты (пластиды вообще). Произошли они от бактериальных симбионтов.
Клеткой-хозяином мог быть относительно крупный анаэробный гетеротрофный прокариот, похожий на амебу. В отличие от других, он мог приобрести способность питаться путем фаго- и пиноцитоза, что позволяло ему захватывать других прокариот. Они не все переваривались, а снабжали хозяина продуктами своей жизнедеятельности). В свою очередь, получали от него питательные вещества.
Митохондрии произошли от аэробных бактерий и позволили клетке-хозяину перейти к аэробному дыханию, которое не только намного эффективней, но и облегчает существование в атмосфере, содержащей достаточно большое количество кислорода. В такой среде аэробные организмы получают преимущество над анаэробными.
Позже в некоторых клетках поселились похожие на ныне живущих синезеленых водорослей (цианобактерий) древние прокариоты. Они стали хлоропластами, дав начало эволюционной ветви растений.
Кроме митохондрий и пластид симбиотическое происхождение могут иметь жгутики эукариот. В них превратились симбионты-бактерии наподобие современных спирохет, имеющих жгутик. Считается, что в последствии из базальных тел жгутиков произошли центриоли, столь важные структуры для механизма клеточного деления эукариот.
Эндоплазматическая сеть, комплекс Гольджи, пузырьки и вакуоли могли произойти от наружной мембраны ядерной оболочки. С другой точки зрения, некоторые из перечисленных органелл могли возникнуть путем упрощения митохондрий или пластид.
Во многом неясным остается вопрос происхождения ядра. Могло ли оно также образоваться из прокариота-симбионта? Количество ДНК в ядре современных эукариот во много раз превышает его количество в митохондриях и хлоропластах. Возможно часть генетической информации последних со временем переместилась в ядро. Также в процессе эволюции происходило дальнейшее увеличение размера ядерного генома.
Кроме того в симбиотической гипотезе происхождения эукариот не все так однозначно с клеткой-хозяином. Им мог и не быть один вид прокариот. Используя методы сравнения геномов, ученые делают вывод, что клетка-хозяин близок к археям, при этом сочетает в себе признаки архей и ряда неродственных групп бактерий. Отсюда можно сделать вывод, что появление эукариот происходило в сложном сообществе прокариот. При этом процесс скорее всего начался с метаногенной археи, вступавшей в симбиоз с другими прокариотами, что было вызвано необходимостью обитания в кислородной среде. Появление фагоцитоза способствовало притоку чужих генов, а ядро образовалось для защиты генетического материала.
Молекулярный анализ показал, что различные белки эукариот происходят от разных групп прокариот.
Доказательства симбиогенеза
В пользу симбиотического происхождения эукариот говорит то, что митохондрии и хлоропласты имеют собственную ДНК, причем кольцевую и не связанную с белками (также обстоит дело у прокариот). Однако в генах митохондрий и пластид есть интроны, чего нет у прокариот.
Пластиды и митохондрии не воспроизводятся клеткой с нуля. Они образуются из ранее существующих таких же органелл путем их деления и последующего роста.
В настоящее время существуют амебы, у которых нет митохондрий, а вместо них есть бактерии симбионты. Также есть простейшие, сожительствующие с одноклеточными водорослями, выполняющими в клетке-хозяине роль хлоропластов.
Инвагинационная гипотеза происхождения эукариот
Кроме симбиогенеза существуют и другие взгляды на происхождение эукариот. Например, инвагинационная гипотеза. Согласно ей, предком эукариотической клетки был не анаэробный, а аэробный прокариот. К такой клетке могли прикрепляться другие прокариоты. Потом их геномы объединялись.
Ядро, митохондрии и пластиды возникли путем впячивания и отшнуровывания участков клеточной мембраны. В эти структуры попадала чужеродная ДНК.
Усложнение генома происходило в процессе дальнейшей эволюции.
Инвагинационная гипотеза происхождения эукариот хорошо объясняет наличие двойной мембраны у органелл. Однако она не объясняет, почему система биосинтеза белка в хлоропластах и митохондриях сходна с прокариотической, в то время как таковая в ядерно-цитоплазматическом комплексе имеет ключевые отличия.
Причины эволюции эукариот
Все разнообразие жизни на Земле (от простейших до покрытосеменных и млекопитающих) дали клетки эукариотического, а не прокариотического типа. Возникает вопрос, почему? Очевидно, ряд особенностей, возникших у эукариот, существенно повысили их эволюционные возможности.
Во-первых, у эукариот есть ядерный геном, который во много раз превосходит количество ДНК у прокариот. При этом эукариотические клетки диплоидны, кроме этого в каждом гаплоидном наборе определенные гены многократно повторяются. Все это обеспечивает, с одной стороны, большие масштабы для мутационной изменчивости, а с другой — уменьшает угрозу резкого снижения жизнеспособности в результате вредной мутации. Таким образом, эукариоты, в отличие от прокариот, обладают резервом наследственной изменчивости.
Эукариотические клетки имеют более сложный механизм регуляции жизнедеятельности, у них существенно больше различных регуляторных генов. Кроме того, молекулы ДНК образовали комплексы с белками, что позволило наследственному материалу упаковываться и распаковываться. Все вместе это дало возможность считывать информацию частями, в разных сочетаниях и количестве, в разное время. (Если в клетках прокариот транскрибируется почти вся информация генома, то в эукариотических клетках обычно менее половины.) Благодаря этому эукариоты могли специализироваться, лучше приспосабливаться.
У эукариот появились митоз, а затем и мейоз. Митоз позволяет воспроизводить генетически сходные клетки, а мейоз сильно увеличивает комбинативную изменчивость, что ускоряет эволюцию.
Большую роль в процветании эукариот сыграло приобретенное их предком аэробное дыхание (хотя оно есть и у многих прокариот).
На заре своей эволюции эукариоты обзавелись эластичной оболочкой, обеспечивавшей возможность фагоцитоза, и жгутиками, позволившими им двигаться. Это дало возможность эффективней питаться.

Биология
Симбиогенез
Теория симбиогенеза (симбиотическая теория, эндосимбиотическая теория, теория эндосимбиоза) объясняет механизм возникновения некоторых органоидов эукариотической клетки — митохондрий, гидрогеносом и пластид.
Схема эволюции эукариотических клеток. 1 — образование двойной мембраны ядра, 2 — приобретение митохондрий, 3 — приобретение пластид, 4 — внедрение получившейся фотосинтезирующей эукариотической клетки в нефотосинтезирующую (например, в ходе эволюции криптофитовых водорослей), 5 — внедрение получившейся клетки снова в нефотосинтезирующую (например, при симбиозе этих водорослей с инфузориями). Цветом обозначен геном предков эукариот, митохондрий и пластид.
История cимбиогенеза
В 1920-е годы теория была развита Б. М. Козо-Полянским, было высказано предположение, что симбионтами являются и митохондрии. Затем долгое время о симбиогенезе практически не упоминали в научной литературе. Второе рождение расширенная и конкретизированная теория получила уже в работах Линн Маргулис начиная с 1960-х годов.
Симбиотическое происхождение митохондрий и пластид
Существуют свидетельства того, что первоначально эндосимбиотические предки митохондрий не могли ни импортировать белки, ни экспортировать АТФ. Вероятно, первоначально они получали от клетки-хозяина пируват, а выгода для хозяина состояла в обезвреживании аэробными симбионтами токсичного для нуклеоцитоплазмы кислорода.
Пластиды, подобно митохондриям, имеют свои собственные прокариотические ДНК и рибосомы. По-видимому, хлоропласты произошли от фотосинтезирующих бактерий, поселившихся в своё время в гетеротрофных клетках протистов, превратив их в автотрофные водоросли.
Доказательства симбиогенеза
Митохондрии и пластиды:
- имеют две полностью замкнутые мембраны. При этом внешняя сходна с мембранами вакуолей, внутренняя — бактерий.
- размножаются бинарным делением (причём делятся иногда независимо от деления клетки), никогда не синтезируются de novo.
- генетический материал — кольцевая ДНК, не связанная с гистонами (По доле ГЦ ДНК митохондрий и пластид ближе к ДНК бактерий, чем к ядерной ДНК эукариот)
имеют свой аппарат синтеза белка — рибосомы и др. - рибосомы прокариотического типа — c константой седиментации 70S. По строению 16s рРНК близки к бактериальной.
- некоторые белки этих органелл похожи по своей первичной структуре на аналогичные белки бактерий и не похожи на соответствующие белки цитоплазмы.
Проблемы симбиогенеза
Примеры эндосимбиозов
В наши дни существует ряд организмов, содержащих внутри своих клеток другие клетки в качестве эндосимбионтов. Они, однако, не являются сохранившимися до наших дней первичными эукариотами, у которых симбионты еще не интегрировались в единое целое и не потеряли своей индивидуальности. Тем не менее, они наглядно и убедительно показывают возможность симбиогенеза.
- Mixotricha paradoxa — наиболее интересный с этой точки зрения организм. Для движения она использует более 250 000 бактерий Treponema spirochetes, прикреплённых к поверхности её клетки. Митохондрии у этого организма вторично потеряны, но внутри его клетки есть сферические аэробные бактерии, заменяющие эти органеллы.
- Амёбы рода Pelomyxa также не содержат митохондрий и образуют симбиоз с бактериями.
- Инфузории рода Paramecium постоянно содержат внутри клеток водоросли, в частности, Paramecium bursaria образует эндосимбиоз с зелёными водорослями рода хлорелла (Chlorella).
- Одноклеточная жгутиковая водоросль Cyanophora paradoxa содержит цианеллы — органоиды, напоминающие типичные хлоропласты красных водорослей, но отличающиеся от них наличием тонкой клеточной стенки, содержащей пептидогликан (размер генома цианелл такой же, как у типичных хлоропластов, и во много раз меньше, чем у цианобактерий).
Гипотезы эндосимбиотического происхождения других органелл
Эндосимбиоз — наиболее широко признанная версия происхождения митохондрий и пластид. Но попытки объяснить подобным образом происхождение других органелл и структур клетки не находят достаточных доказательств и наталкиваются на обоснованную критику.
Клеточное ядро, нуклеоцитоплазма
Смешение у эукариот многих свойств, характерных для архей и бактерий, позволило предположить симбиотическое происхождение ядра от метаногенной архебактерии, внедрившейся в клетку миксобактерии. Гистоны, к примеру, обнаружены у эукариот и некоторых архей, кодирующие их гены весьма схожи. Другая гипотеза, объясняющая сочетание у эукариот молекулярных признаков архей и эубактерий, состоит в том, что на некотором этапе эволюции похожие на архей предки нуклеоцитоплазматического компонента эукариот приобрели способность к усиленному обмену генами с эубактериями путём горизонтального переноса генов.
В последнее десятилетие сформировалась также гипотеза вирусного эукариогенеза. В её основании лежит ряд сходств устройства генетического аппарата эукариот и вирусов: линейное строение ДНК, её тесное взаимодействие с белками и др. Было показано сходство ДНК-полимеразы эукариот и поксивирусов, что сделало именно их предков основными кандидатами на роль ядра.
Жгутики и реснички
Линн Маргулис в книге Symbiosis in Cell Evolution (1981) предположила в том числе происхождение жгутиков и ресничек от симбиотических спирохет. Несмотря на сходство размеров и строения указанных органелл и бактерий и существование Mixotricha paradoxa, использующей спирохет для движения, в жгутиках не было найдено никаких специфически спирохетных белков. Однако известен общий для всех бактерий и архей белок FtsZ, гомологичный тубулину и, возможно, являющийся его предшественником. Жгутики и реснички не обладают такими признаками бактериальных клеток, как замкнутая наружная мембрана, собственный белоксинтезирующий аппарат и способность к делению. Данные о наличии ДНК в базальных тельцах, появившиеся в 1990-е годы, были впоследствии опровергнуты. Увеличение числа базальных телец и гомологичных им центриолей происходит не путём деления, а путём достраивания нового органоида рядом со старым.
Пероксисомы
Кристиан де Дюв обнаружил пероксисомы в 1965 году. Ему же принадлежит предположение, что пероксисомы были первыми эндосимбионтами эукариотической клетки, позволившими ей выживать при нарастающем количестве свободного молекулярного кислорода в земной атмосфере. Пероксисомы, однако, в отличие от митохондрий и пластид, не имеют ни генетического материала, ни аппарата для синтеза белка. Было показано, что эти органеллы формируются в клетке de novo в ЭПР и нет никаких оснований считать их эндосимбионтами.

Пятьдесят лет назад, в 1967 году, Линн Маргулис (Lynn Margulis) опубликовала развернутое изложение симбиогенетической теории, согласно которой эукариоты (организмы с клеточными ядрами) возникли в результате серии объединений разных клеток между собой. Современная поправка к этой теории гласит, что в основе становления эукариот, по-видимому, была не общая тенденция, охватившая многие эволюционные ветви (как предполагала Маргулис), а уникальное событие, приведшее к слиянию клеток археи и протеобактерии. В результате образовалась сложная клетка с митохондриями, которая и стала первым эукариотом. Дальнейшие симбиогенетические события — например, захват водорослей, ставших хлоропластами, — действительно происходили много раз, но с возникновением эукариот как таковых они не связаны.
Выход в 1967 году статьи Линн Маргулис (будем для удобства звать ее так) стал началом обновления биологических представлений, которое многие авторы расценили как смену парадигм — то есть, иными словами, как самую настоящую научную революцию (И. М. Мирабдуллаев, 1991. Эндосимбиотическая теория — от фантастики к парадигме). Суть интриги тут проста. Со времен Чарльза Дарвина биологи были убеждены, что основным способом эволюции является дивергенция — расхождение ветвей. Линн Маргулис была первой, кто сумел по-настоящему убедительно объяснить научному сообществу, что механизм некоторых крупных эволюционных событий, скорее всего, был принципиально другим. В центре интересов Маргулис оказалась проблема происхождения эукариот — организмов, клетка которых обладает сложной внутренней структурой с ядром. К эукариотам относятся животные, растения, грибы и многие одноклеточные — амебы, жгутиконосцы, инфузории и прочие. Маргулис показала, что ранняя эволюция эукариот вовсе не сводилась к дивергенции — она включала в себя слияние эволюционных ветвей, причем неоднократное. Дело в том, что по меньшей мере два типа эукариотных органелл — митохондрии, благодаря которым мы можем дышать кислородом, и хлоропласты, осуществляющие фотосинтез, — происходят не от того предка, что основная часть эукариотной клетки (рис. 1). И митохондрии, и хлоропласты — это бывшие бактерии, изначально совсем не родственные эукариотам (протеобактерии в случае митохондрий и цианобактерии — в случае хлоропластов). Эти бактерии были поглощены клеткой древнего эукариота (либо предка эукариот) и продолжили жить внутри нее, сохраняя до поры собственный генетический аппарат.
Таким образом, эукариотная клетка — это, по выражению Маргулис, мультигеномная система. И возникла она в результате симбиоза, то есть взаимовыгодного сожительства разных организмов (точнее, эндосимбиоза, один из участников которого живет внутри другого). Соответствующие эволюционные ветви при этом, разумеется, слились. Такой взгляд на эволюцию получил название теории симбиогенеза.
Сейчас теория симбиогенеза общепринята. Она подтверждена настолько строго, насколько вообще можно подтвердить какую бы то ни было теорию, касающуюся крупномасштабной эволюции. Но научные концепции, в отличие от религиозных догматов, никогда не остаются статичными. Естественно, что общая картина симбиогенеза выглядит для нас сейчас не совсем так (а местами и совсем не так), как представляла ее себе Линн Маргулис полвека назад.
Логика классика
К пятидесятилетию выхода знаменитой статьи о симбиогенезе Journal of Theoretical Biology подготовил специальный выпуск, целиком посвященный творческому наследию Линн Маргулис. В этот выпуск входит обстоятельная статья известного британского биохимика и популяризатора науки Ника Лейна (Nick Lane), в которой современное состояние проблемы происхождения эукариот сравнивается с классическими идеями на эту тему. Лейн нисколько не сомневается, что в основных утверждениях (касающихся происхождения митохондрий и хлоропластов) Маргулис была права; в наше время в этом не сомневается, кажется, никто из серьезных ученых, ибо данные молекулярной биологии на этот счет однозначны. Но дьявол, как известно, живет в деталях. В данном случае мы можем, погрузившись в детали, найти там много нового и интересного, а главное — убедиться, что тема происхождения эукариот далеко не исчерпана.
Начнем с того, что кое-какие частные предположения Маргулис оказались неверны. Это нормально: учитывая огромную скорость развития биологии, просто невероятно, чтобы в статье, опубликованной полвека назад, было точно угадано абсолютно всё. Новые факты, которые не могли быть в свое время известны автору, обязательно внесут какие-нибудь коррективы. Так получилось и тут. Прежде всего, Маргулис настаивала на симбиотическом происхождении не только митохондрий и хлоропластов, но и эукариотных жгутиков. Она считала, что предками жгутиков были закрепившиеся на эукариотной клетке длинные спирально закрученные подвижные бактерии, похожие на современных спирохет (см. рис. 1). Увы, эта гипотеза не получила никаких молекулярно-биологических подтверждений, и сейчас ее больше никто не поддерживает.
Однако по большому счету всё это частности. Мышление Линн Маргулис было синтетическим: она не ограничивалась объяснениями отдельных фактов, а стремилась свести их в целостную систему, описывающую эволюцию живых организмов в контексте истории Земли (рис. 2). Современные научные знания позволяют проверить эту систему представлений на прочность.

Древо и сеть
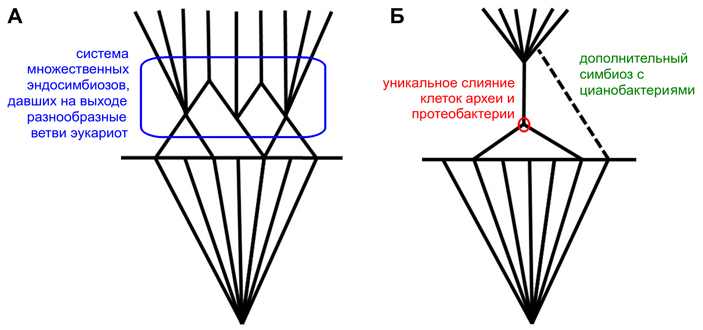

Надо сказать, что к концу XX века в эволюционной биологии (и особенно в палеонтологии) и без того завоевала определенную популярность идея, что большинство крупных эволюционных событий имеет закономерный и системный характер. Подобное событие охватывает сразу много эволюционных ветвей, в которых под действием общей наследственности параллельно возникают примерно одни и те же признаки (см., например: А. Г. Пономаренко, 2004. Артроподизация и ее экологические последствия). Примерами таких событий называли маммализацию (происхождение млекопитающих), ангиоспермизацию (происхождение цветковых растений), артроподизацию (происхождение членистоногих), тетраподизацию (происхождение наземных позвоночных), орнитизацию (происхождение птиц) и многое другое. Казалось, что становление эукариот — эукариотизация — великолепно вписывается в этот ряд.
Проблема митохондрий
Начнем с того, что обсуждавшаяся Еськовым гипотеза насчет красных водорослей теперь устарела. Молекулярные исследования показывают, что эволюционная линия красных водорослей находится глубоко внутри древа эукариот (они достаточно близкие родственники зеленых растений), и их независимая эукариотизация крайне маловероятна.
Но гораздо серьезнее другое. Если симбиогенез был закономерным, долгим, многоступенчатым процессом, да еще и шел параллельно в разных эволюционных ветвях, то следовало бы ожидать, что мы увидим спектр довольно разнообразных переходных состояний между эукариотами и не-эукариотами. Маргулис именно так и думала. То, что эти переходные состояния не бросаются в глаза, она (насколько можно судить) считала проблемой чисто технической, связанной с недостатком знаний и несовершенством методов. Подтверждается ли это сейчас, когда мы знаем о живых клетках неизмеримо больше, чем знали пятьдесят лет назад?
Обратим внимание, что в этот набор признаков не входят хлоропласты. Они появились далеко не у всех эукариот и не сразу. Кроме того, хлоропласты уж точно приобретались неоднократно, причем разными способами в разных эволюционных ветвях. Хлоропласты бывают как первичные (когда эукариот захватывает цианобактерию), так и вторичные (когда эукариот захватывает другого эукариота с цианобактерией внутри) и даже третичные (когда один эукариот захватывает второго эукариота, внутри которого живет третий эукариот, а уж внутри того — цианобактерия). Здесь эволюция, что называется, разгулялась. С митохондриями ситуация совершенно иная: по признаку их наличия мы не видим никакого особого разнообразия и никаких переходных стадий (если не считать многочисленных фактов вторичной потери, но о происхождении эукариот такие факты не говорят ровно ничего). Если бы сценарий Маргулис был полностью верен, то и с митохондриями, и со жгутиками дело обстояло бы примерно так же, как с хлоропластами, — но этого нет.
Уникальное событие
В чем тут дело? Согласно классической теории, все внутренние симбионты были приобретены эукариотными клетками путем фагоцитоза, то есть захвата ложноножками с изоляцией захваченного объекта и последующим его перевариванием (в данном случае — несостоявшимся). В отношении хлоропластов это, по всей видимости, верно, а вот в отношении митохондрий — очень сомнительно. Предположение, что фагоцитоз появился раньше, чем митохондрии, плохо согласуется с данными биоинформатики. Сравнительный анализ белковых последовательностей показывает, что актиновые микрофиламенты, образующие внутренний каркас любых ложноножек, скорее всего, сначала были неподвижными — белки, позволяющие им еще и сокращаться, появились заметно позже (Е. В. Кунин, 2014. Логика случая). А это означает, что начаться прямо с фагоцитоза эволюция эукариот не могла — митохондрии были приобретены каким-то другим способом.
Здесь открывается большой простор для гипотез. Есть, например, весьма изящно обоснованное предположение, что архейная клетка не поглощала предков митохондрий путем фагоцитоза, а охватила их своими постепенно разраставшимися выростами, которые в конце концов сомкнулись, изолировав митохондрии от внешней среды и заодно создав систему внутриклеточных полостей — то, что сейчас называется эндоплазматической сетью (D. Baum, B. Baum, 2014. An inside-out origin for the eukaryotic cell). Есть другая остроумная гипотеза: предки митохондрий были внутриклеточными паразитами (Z. Wang, M. Wu, 2014. Phylogenomic reconstruction indicates mitochondrial ancestor was an energy parasite). Авторы этой последней идеи обратили внимание, что в группе альфа-протеобактерий, из которой, несомненно, вышли митохондрии, есть немало специализированных внутриклеточных паразитов — например, риккетсии.
Но надо подчеркнуть, что всё это пока лишь предположения. Загадка происхождения митохондрий, не говоря уж о происхождении ядра, до сих пор не разгадана.
Случайность и необходимость
Итак, верна ли гипотеза сериального эндосимбиоза? Да — в том смысле, что в истории эукариот действительно много раз случались симбиотические события. Лучше всего это иллюстрирует долгая, богатая и неплохо сейчас изученная история хлоропластов (P. Keeling et al., 2013. The number, speed, and impact of plastid endosymbioses in eukaryotic evolution). Нет — в том смысле, что сериальный эндосимбиоз не был предпосылкой возникновения эукариот как группы. Эндосимбиотическое событие, которое привело к возникновению эукариот, было, насколько мы сейчас можем судить, уникальным.
Источник: Nick Lane. Serial endosymbiosis or singular event at the origin of eukaryotes? // Journal of Theoretical Biology. 7 December 2017. V. 434. P. 58–67.
Читайте также: