
С4 путь фотосинтеза кратко
Обновлено: 05.07.2024
Фотосинтез — это преобразование энергии света в энергию химических связей органических соединений.
Фотосинтез характерен для растений, в том числе всех водорослей, ряда прокариот, в том числе цианобактерий, некоторых одноклеточных эукариот.
В большинстве случаев при фотосинтезе в качестве побочного продукта образуется кислород (O2). Однако это не всегда так, поскольку существует несколько разных путей фотосинтеза. В случае выделения кислорода его источником является вода, от которой на нужды фотосинтеза отщепляются атомы водорода.
Фотосинтез состоит из множества реакций, в которых участвуют различные пигменты, ферменты, коферменты и др. Основными пигментами являются хлорофиллы, кроме них — каротиноиды и фикобилины.
Выделяют две фазы фотосинтеза — световую и темновую. Первая зависит от светового излучения (hν), которое необходимо для протекания реакций. Темновая фаза является светонезависимой.
У растений фотосинтез протекает в хлоропластах. В результате всех реакций образуются первичные органические вещества, из которых потом синтезируются углеводы, аминокислоты, жирные кислоты и др. Обычно суммарную реакцию фотосинтеза пишут в отношении глюкозы — наиболее распространенного продукта фотосинтеза:
Атомы кислорода, входящие в молекулу O2, берутся не из углекислого газа, а из воды. Углекислый газ – источник углерода, что более важно. Благодаря его связыванию у растений появляется возможность синтеза органики.
Представленная выше химическая реакция есть обобщенная и суммарная. Она далека от сути процесса. Так глюкоза не образуется из шести отдельных молекул углекислоты. Связывание CO2 происходит по одной молекуле, которая сначала присоединяется к уже существующему пятиуглеродному сахару.
Для прокариот характерны свои особенности фотосинтеза. Так у бактерий главный пигмент — бактериохлорофилл, и не выделяется кислород, так как водород берется не из воды, а часто из сероводорода или других веществ. У сине-зеленых водорослей основным пигментом является хлорофилл, и при фотосинтезе выделяется кислород.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.
Темновая фаза фотосинтеза
Впервые темновую фазу фотосинтеза подробно изучили Кальвин, Бенсон, Бэссем. Открытый ими цикл реакций в последствии был назван циклом Кальвина, или C3-фотосинтезом. У определенных групп растений наблюдается видоизмененный путь фотосинтеза – C4, также называемый циклом Хэтча-Слэка.
В темновых реакциях фотосинтеза происходит фиксация CO2. Темновая фаза протекает в строме хлоропласта.
Восстановление CO2 происходит за счет энергии АТФ и восстановительной силы НАДФ·H2, образующихся в световых реакциях. Без них фиксации углерода не происходит. Поэтому хотя темновая фаза напрямую не зависит от света, но обычно также протекает на свету.
Цикл Кальвина
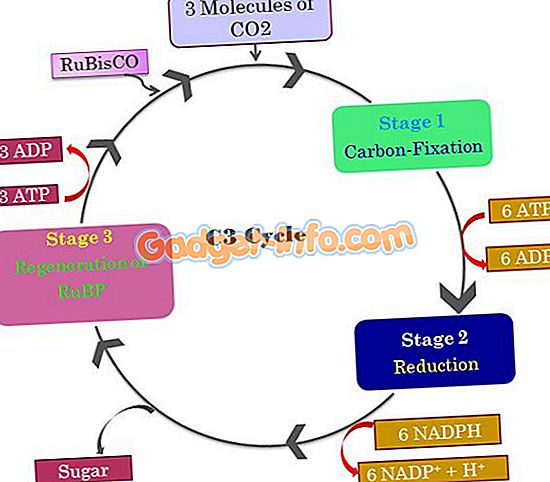
Первая реакция темновой фазы – присоединение CO2 (карбоксилирование) к 1,5-рибулезобифосфату (рибулезо-1,5-дифосфат) – РиБФ. Последний представляет собой дважды фосфорилированную рибозу. Данную реакцию катализирует фермент рибулезо-1,5-дифосфаткарбоксилаза, также называемый рубиско.
В результате карбоксилирования образуется неустойчивое шестиуглеродное соединение, которое в результате гидролиза распадается на две трехуглеродные молекулы фосфоглицериновой кислоты (ФГК) – первый продукт фотосинтеза. ФГК также называют фосфоглицератом.
ФГК содержит три атома углерода, один из которых входит в состав кислотной карбоксильной группы (-COOH):
Из ФГК образуется трехуглеродный сахар (глицеральдегидфосфат) триозофосфат (ТФ), включающий уже альдегидную группу (-CHO):
ФГК (3-кислота) → ТФ (3-сахар)
На данную реакцию затрачивается энергия АТФ и восстановительная сила НАДФ · H2. ТФ — первый углевод фотосинтеза.
После этого большая часть триозофосфата затрачивается на регенерацию рибулозобифосфата (РиБФ), который снова используется для связывания CO2. Регенерация включает в себя ряд идущих с затратой АТФ реакций, в которых участвуют сахарофосфаты с количеством атомов углерода от 3 до 7.
В таком круговороте РиБФ и заключается цикл Кальвина.
Из цикла Кальвина выходит меньшая часть образовавшегося в нем ТФ. В перерасчете на 6 связанных молекул углекислого газа выход составляет 2 молекулы триозофосфата. Суммарная реакция цикла с входными и выходными продуктами:
При этом в связывании участвую 6 молекул РиБФ и образуется 12 молекул ФГК, которые превращаются в 12 ТФ, из которых 10 молекул остаются в цикле и преобразуются в 6 молекул РиБФ. Поскольку ТФ — это трехуглеродный сахар, а РиБФ — пятиуглеродный, то в отношении атомов углерода имеем: 10 * 3 = 6 * 5. Количество атомов углерода, обеспечивающих цикл не изменяется, весь необходимый РиБФ регенерируется. А шесть вошедших в цикл молекул углекислоты затрачиваются на образование двух выходящих из цикла молекул триозофосфата.
На цикл Кальвина в расчете на 6 связанных молекул CO2 затрачивается 18 молекул АТФ и 12 молекул НАДФ · H2, которые были синтезированы в реакциях световой фазы фотосинтеза.
Расчет ведется на две выходящие из цикла молекулы триозофосфата, так как образующаяся в последствии молекула глюкозы, включает 6 атомов углерода.
Триозофосфат (ТФ) — конечный продукт цикла Кальвина, но его сложно назвать конечным продуктом фотосинтеза, так как он почти не накапливается, а, вступая в реакции с другими веществами, превращается в глюкозу, сахарозу, крахмал, жиры, жирные кислоты, аминокислоты. Кроме ТФ важную роль играет ФГК. Однако подобные реакции происходят не только у фотосинтезирующих организмов. В этом смысле темновая фаза фотосинтеза – это то же самое, что цикл Кальвина.
Из ФГК путем ступенчатого ферментативного катализа образуется шестиуглеродный сахар фруктозо-6-фосфат, который превращается в глюкозу. В растениях глюкоза может полимеризоваться в крахмал и целлюлозу. Синтез углеводов похож на процесс обратный гликолизу.
Фотодыхание
Кислород подавляет фотосинтез. Чем больше O2 в окружающей среде, тем менее эффективен процесс связывания CO2. Дело в том, что фермент рибулозобифосфат-карбоксилаза (рубиско) может реагировать не только с углекислым газом, но и кислородом. В этом случае темновые реакции несколько иные.
Содержащая пять атомов углерода молекула рибулозобифосфата реагирует уже не с CO2, а с O2. В результате чего образуются по одной молекуле фосфогликолата (C2) и фосфоглицериновой кислоты (C3), а не две ФГК как обычно.
Фотодыхание — это зависимое от света потребление кислорода и выделение углекислого газа. То есть обмен газов происходит как при дыхании, но протекает в хлоропластах и зависит от светового излучения. От света фотодыхание зависит лишь потому, что рибулозобифосфат образуется только при фотосинтезе.
При фотодыхании происходит возврат атомов углерода из гликолата в цикл Кальвина в виде фосфоглицериновой кислоты (фосфоглицерата).
2 Гликолат (С2) → 2 Глиоксилат (С2) →2 Глицин (C2) - CO2 → Серин (C3) →Гидроксипируват (C3) → Глицерат (C3) → ФГК (C3)
Как видно, возврат происходит не полный, так как один атом углерода теряется при превращении двух молекул глицина в одну молекулу аминокислоты серина, при этом выделяется углекислый газ.
Кислород необходим на стадиях превращения гликолата в глиоксилат и глицина в серин.
Превращения гликолата в глиоксилат, а затем в глицин происходят в пероксисомах, синтез серина в митохондриях. Серин снова поступает в пероксисомы, где из него сначала получается гидрооксипируват, а затем глицерат. Глицерат уже поступает в хлоропласты, где из него синтезируется ФГК.
Фотодыхание характерно в основном для растений с C3-типом фотосинтеза. Его можно считать вредным, так как энергия бесполезно тратится на превращения гликолата в ФГК. Видимо фотодыхание возникло из-за того, что древние растения были не готовы к большому количеству кислорода в атмосфере. Изначально их эволюция шла в атмосфере богатой углекислым газом, и именно он в основном захватывал реакционный центр фермента рубиско.
C4-фотосинтез, или цикл Хэтча-Слэка
Если при C3-фотосинтезе первым продуктом темновой фазы является фосфоглицериновая кислота, включающая три атома углерода, то при C4-пути первыми продуктами являются кислоты, содержащие четыре атома углерода: яблочная, щавелевоуксусная, аспарагиновая.
С4-фотосинтез наблюдается у многих тропических растений, например, сахарного тростника, кукурузы.
С4-растения эффективнее поглощают оксид углерода, у них почти не выражено фотодыхание.
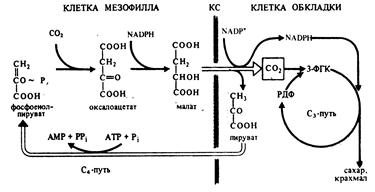
Растения, в которых темновая фаза фотосинтеза протекает по C4-пути, имеют особое строение листа. В нем проводящие пучки окружены двойным слоем клеток. Внутренний слой — обкладка проводящего пучка. Наружный слой — клетки мезофилла. Хлоропласты клеток слоев отличаются друг от друга.
Для мезофильных хлоропласт характерны крупные граны, высокая активность фотосистем, отсутствие фермента РиБФ-карбоксилазы (рубиско) и крахмала. То есть хлоропласты этих клеток адаптированы преимущественно для световой фазы фотосинтеза.
В хлоропластах клеток проводящего пучка граны почти не развиты, зато высока концентрация РиБФ-карбоксилазы. Эти хлоропласты адаптированы для темновой фазы фотосинтеза.
Углекислый газ сначала попадает в клетки мезофилла, связывается с органическими кислотами, в таком виде транспортируется в клетки обкладки, освобождается и далее связывается также, как у C3-растений. То есть C4-путь дополняет, а не заменяет C3.
В мезофилле CO2 присоединяется к фосфоенолпирувату (ФЕП) с образованием оксалоацетата (кислота), включающего четыре атома углерода:
Реакция происходит при участии фермента ФЕП-карбоксилазы, обладающего более высоким сродством к CO2, чем рубиско. К тому же ФЕП-карбоксилаза не взаимодействует с кислородом, а значит не затрачивается на фотодыхание. Таким образом, преимущество C4-фотосинтеза заключается в более эффективной фиксации углекислоты, увеличению ее концентрации в клетках обкладки и следовательно более эффективной работе РиБФ-карбоксилазы, которая почти не расходуется на фотодыхание.
Оксалоацетат превращается в 4-х углеродную дикарбоновую кислоту (малат или аспартат), которая транспортируется в хлоропласты клеток обкладки проводящих пучков. Здесь кислота декарбоксилируется (отнятие CO2), окисляется (отнятие водорода) и превращается в пируват. Водород восстанавливает НАДФ. Пируват возвращается в мезофилл, где из него регенерируется ФЕП с затратой АТФ.
Оторванный CO2 в хлоропластах клеток обкладки уходит на обычный C3-путь темновой фазы фотосинтеза, т. е. в цикл Кальвина.
Фотосинтез по пути Хэтча-Слэка требует больше энергозатрат.
Считается, что C4-путь возник в эволюции позже C3 и во многом является приспособлением против фотодыхания.
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов:
1) мелкие гранальные пластиды в клетках мезофилла листа;
2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки.
К С4-растениям относится ряд культурных растений преимущественно тропического и субтропического происхождения – кукуруза, просо, сорго, сахарный тростник и многие злостные сорняки – свинорой, сыть округлая, просо куриное, гумай, щетинник и др. Как правило, это высокопродуктивные растения, устойчиво осуществляющие фотосинтез при значительных повышениях температуры и в засушливых условиях.
Акцептором CO2 является ФЕП фосфоенолпировиноградная кислота;
Фотосинтез разделен в пространстве
Конечными продуктами являются: органические кислоты, фермент ФЕП-карбоксилаза;
Отсутствует процесс фотодыхания;
Процесс карбоксилирования осуществляется дважды и это позволяет CO2 поступать при закрытых устьицах.
Характерным признаком растений С4-пути является то, что образование продуктов цикла Кальвина происходит в хлоропластах, расположенных непосредственно около проводящих пучков. Это благоприятствует оттоку ассимилятов и, как следствие, повышает интенсивность фотосинтеза.
1. карбоксилирование (происходит в клетках мезофилла);
2. декарбоксилирование и синтез углеводов (в клетках обкладки проводящих пучков).
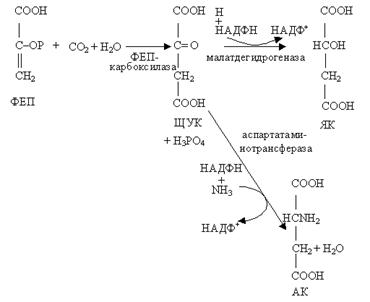
Карбоксилированию подвергаются фосфоенолпировиноградная кислота (ФЕП) при участии ФЕП-карбоксилазы и образуется щавелевоуксусная кислота (ЩУК), которая восстанавливается до яблочной кислоты (малат) или аминируется с образованием аспарагиновой кислоты.
ЩУК, малат и аспарагиновая кислоты являются четырехуглеродными соединениями.
В клетках обкладки яблочная кислота декарбоксилируется ферментом малатдегидрогеназой до пировиноградной кислоты (пируват, ПВК) и С02. Реакция декарбоксилирования может варьировать у разных групп растений с использованием разных ферментов. С02 поступает в хлоропласты клеток обкладки и включается в цикл Кальвина—присоединяется к РДФ. Пируват возвращается в клетки мезофилла и превращается в первичный акцептор С02 — ФЕП. Таким образом, при С4-пути реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Акцепторы С02 (ФЕП и РДФ) регенерируют, что и создает возможность непрерывного функционирования циклов. Фиксация С02 с участием ФЕП и образованием малата или аспартата служит своеобразным насосом для поставки С02 в хлоропласты обкладки, функционирующих по С3-пути.
Исследования показали, что в растениях, в которых процесс фотосинтеза протекает по С4-пути, имеются два типа клеток и хлоропластов:
1) мелкие гранальные пластиды в клетках мезофилла листа;
2) крупные пластиды, часто лишенные гран, в клетках обкладки, окружающих сосудистые пучки.
К С4-растениям относится ряд культурных растений преимущественно тропического и субтропического происхождения – кукуруза, просо, сорго, сахарный тростник и многие злостные сорняки – свинорой, сыть округлая, просо куриное, гумай, щетинник и др. Как правило, это высокопродуктивные растения, устойчиво осуществляющие фотосинтез при значительных повышениях температуры и в засушливых условиях.
Акцептором CO2 является ФЕП фосфоенолпировиноградная кислота;
Фотосинтез разделен в пространстве
Конечными продуктами являются: органические кислоты, фермент ФЕП-карбоксилаза;
Отсутствует процесс фотодыхания;
Процесс карбоксилирования осуществляется дважды и это позволяет CO2 поступать при закрытых устьицах.
Характерным признаком растений С4-пути является то, что образование продуктов цикла Кальвина происходит в хлоропластах, расположенных непосредственно около проводящих пучков. Это благоприятствует оттоку ассимилятов и, как следствие, повышает интенсивность фотосинтеза.
1. карбоксилирование (происходит в клетках мезофилла);
2. декарбоксилирование и синтез углеводов (в клетках обкладки проводящих пучков).
Карбоксилированию подвергаются фосфоенолпировиноградная кислота (ФЕП) при участии ФЕП-карбоксилазы и образуется щавелевоуксусная кислота (ЩУК), которая восстанавливается до яблочной кислоты (малат) или аминируется с образованием аспарагиновой кислоты.

ЩУК, малат и аспарагиновая кислоты являются четырехуглеродными соединениями.
В клетках обкладки яблочная кислота декарбоксилируется ферментом малатдегидрогеназой до пировиноградной кислоты (пируват, ПВК) и С02. Реакция декарбоксилирования может варьировать у разных групп растений с использованием разных ферментов. С02 поступает в хлоропласты клеток обкладки и включается в цикл Кальвина—присоединяется к РДФ. Пируват возвращается в клетки мезофилла и превращается в первичный акцептор С02 — ФЕП. Таким образом, при С4-пути реакция карбоксилирования происходит дважды. Это позволяет растению создавать запасы углерода в клетках. Акцепторы С02 (ФЕП и РДФ) регенерируют, что и создает возможность непрерывного функционирования циклов. Фиксация С02 с участием ФЕП и образованием малата или аспартата служит своеобразным насосом для поставки С02 в хлоропласты обкладки, функционирующих по С3-пути.
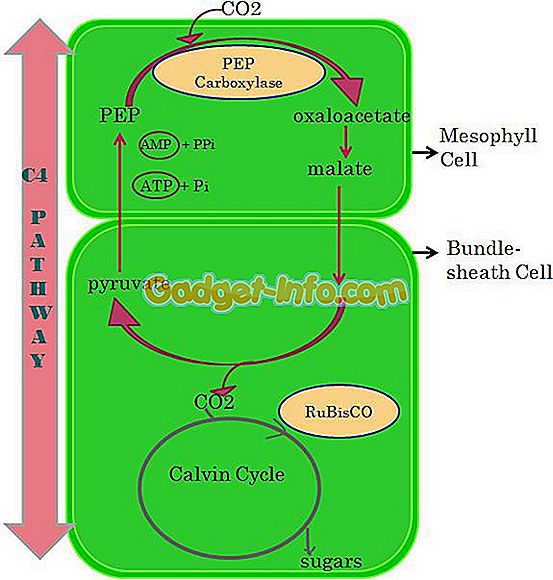
Цикл Кальвина не единственный путь фиксации углерода в темновых реакциях. У некоторых растений первый продукт фиксации не трехуглеродная молекула 3-фосфоглицерата, как в цикле Кальвина, а четырехуглеродное соединение оксалоацетат (который образуется и в цикле Кребса). Растения, которые осуществляют этот путь, называются С4- растениями в отличие от С3-растений, где функционирует только цикл Кальвина. (С4-путь называют также циклом Хэтча Слэка в честь австралийских физиологов растений, которые сыграли решающую роль в его открытии.) Оксалоацетат образуется, когда С02 фиксируется на фосфоенолпирувате.

Эта реакция катализируется ферментом фосфоенолпируваткарбоксилазой (рис. 7-17). Оксалоацетат затем восстанавливается до малата или превращается с добавлением аминогруппы в аспартат. Эти реакции происходят в клетках мезофилла. Следующий этап поразителен: малат (или аспартат в зависимости от вида растения) перемещается из клеток мезофилла в клетки обкладок сосудистых пучков листа, где декарбоксилируется с образованием С02 и пирувата. С02 затем вступает в цикл Кальвина, реагирует с РБФ с образованием фосфоглицерата и других промежуточных соединений цикла, в то время как пируват возвращается в клетки мезофилла, где реагирует с АТР, образуя фосфоенолпируват (рис. 7-18).
Таким образом, анатомия растения способствует пространственному разделению С4- пути и цикла Кальвина в листьях С4-растений. Два первичных карбоксилирующих фермента используют различные формы С02 в качестве субстрата. Рибулозо- бисфосфаткарбоксилаза использует С02, который присутствует в клетке в концентрации около 1520 мкМ в равновесии с воздухом.
В то же время фосфоенолпируваткарбокси- лаза использует гидратированную форму двуокиси углерода HCOj. При pH 8,0 HCOj обнаруживается в клетке в концентрации 1520 мкМ в равновесии с воздушной фазой. Рибулозобисфосфаткарбоксилаза найдена в хлоропластах, в то время как фосфоенолпируваткарбоксилаза находится в основном веществе цитоплазмы.

Кроме того, в хлоропластах клеток обкладки во время фотосинтеза, как правило, образуются более крупные и многочисленные зерна крахмала, чем в хлоропластах мезофилла. Эффективность С4-растений Фиксация С02 у С4-растений требует более высоких энергетических затрат, чем у С3-растений. В С4-пути на каждую ассимилированную молекулу С02 должна образоваться молекула фосфоенолпирувата; при этом тратятся две высокоэнергетические фосфатные группы АТР. С4-растениям нужно затратить пять молекул АТР на фиксацию одной молекулы С02, а С3-растениям только три молекулы АТР.

Может возникнуть вопрос, почему у С4-растений в процессе эволюции возник такой сложный и энергетически дорогой способ обеспечения цикла Кальвина углекислотой. Это станет ясно, если уточнить, что фотосинтез у С3-растений всегда сопровождается фотодыханием процессом, в ходе которого на свету потребляется кислород и выделяется С02 (рис. 7-20). Фотодыхание это расточительный процесс. По сравнению с митохондриальным дыханием фотодыхание не сопровождается окислительным фосфорилированием и не дает АТР. Более того, фотодыхание отвлекает восстановитель, образуемый в световой стадии, от биосинтеза глюкозы на восстановление кислорода.
При нормальных атмосферных условиях до 50% углерода, ассимилированного при фотосинтезе у С3-растений, может реокисляться до С02 в процессе фото дыхания. Таким образом, столь активное фотодыхание у С3-растений ограничивает в значительной степени их эффективность, а у С4-растений оно практически отсутствует. Основной субстрат, окисляемый при фотодыхании у С3- растений, это гликолевая кислота. Она окисляется в пероксисомах фотосинтезирующих клеток и образуется при окислительном расщеплении рибулозобисфосфата с помощью рубилозобисфосфаткарбоксилазы основного фермента, превращающего С02 в фосфоглицерат.
Как это происходит?
Высокая концентрация С02 и низкая 02 ограничивают фото дыхание. Соответственно С4-растения имеют значительное преимущество перед С3-растениями, поскольку С02, фиксируемый в С4-пути, перекачивается активно из клеток мезофилла в клетки обкладки, тем самым поддерживая высокое отношение С02/02 в участках, где сосредоточена активность рибулозобисфосфаткарбоксилазы. Это благоприятствует реакции карбоксилирования. Кроме того, цикл Кальвина и фотодыхание проходят во внутреннем слое клеток обкладок проводящих пучков, и СО,, выделяемый при фотодыхании, может быть рефиксирован в наружном слое клеток мезофилла в С4-пути, который здесь осуществляется.
Таким образом, С02, образующийся при фотодыхании, не выходит из листа. Более того, С4-растения по сравнению с С3-растениями лучше утилизируют внешний С02; это обусловлено тем, что фосфоенолпируваткарбоксилаза не ингибируется 02. В результате скорость нетто-фотосинтеза у С4- трав, таких, как кукуруза (Zea mays), сахарный тростник (Saccharum officinale), сорго (Sorghum vulgare), может быть в 23 раза выше, чем у С3-трав, таких, как пшеница (Triticum aestivum), рожь (Secale cereale), овес (Avena sativa) и рис (Oryza sativa). С4-растения появились в процессе эволюции преимуще- ственно в тропиках; особенно хорошо они адаптированы к высокой инсоляции, повышенным температурам и засухе.
Оптимальная температура для осуществления фотосинтеза у С4-растений выше, чем у С3-растений; С4-растения процветают даже при температурах, которые были бы детальны для многих С3-ВИД0В. С4-растения, обладая более эффективным способом использования С02, могут достигать той же скорости фотосинтеза, что и С3-растения, но с меньшей потерей воды, поскольку у них может быть открыто меньшее число устьиц. Анализ географического распределения С4- видов в Северной Америке показывает, что в основном они наиболее многочисленны в климатических зонах с высокими температурами.
Однако существуют различия между однодольными и двудольными в характере реакции на высокую температуру. Например, С4-злаки преобладают в районах с очень высокой температурой, приходящейся на вегетационный сезон (рис. 7-21). В то же время С4-двудольные широко распространены в тех районах, где вегетационный сезон характеризуется чрезмерной засушливостью. Яркую иллюстрацию особого типа роста у С4-растений можно найти на газонах, которые в северных районах США состоят из С3-трав, таких, как мятлик луговой (Роа pratensis) или полевица тонкая (Agrostis tenuis).
Росичка кровяная (Di- gitaria sanguinalis), которая часто подавляет эти темно-зеленые тонколистные травы своими желтовато-зелеными широкими листьями, относится к С4-злакам и растет быстрее в жаркое лето, чем умеренные С3-злаки; В настоящее время известно, что все растения с С4-фото- синтезом это цветковые из 19 семейств (3 однодольных и 16 двудольных растений), однако нет семейства, которое состояло бы только из С4-видов. Несомненно, этот путь фотосинтеза возникал в ходе эволюции многократно и независимо.
П. Рейвн, Р. Эверт, С. Айкхорн, Современная ботаника В 2-х томах, Том 1, Перевод с английского канд. биол. наук В. Н. Гладковой, проф. М. Ф. Даниловой, д-ра биол. наук И. М. Кислюк, канд. биол. наук Н. С. Мамушиной под редакцией акад. А. Л. Тахтаджяна

Усвоение углекислого газа из солнечного света для процесса фотосинтеза, а затем превращение его в глюкозу (энергию), синтезирующую различные продукты, является ключевым отличием между ними. Таким образом, во время фиксации CO2, когда фотосинтетические растения производят 3-фосфоглицериновую кислоту (PGA) или 3-углеродную кислоту в качестве первого продукта, это называется C3-путь .
Но когда фотосинтезирующее растение, прежде чем идти по пути С3, производит щавелевоуксусную кислоту (ОАА) или 4 -углеродное соединение в качестве своего первого стабильного продукта, его называют С4 или путь Хэтча и Шлака . Но когда растения поглощают энергию солнечного света в дневное время и используют эту энергию для усвоения или фиксации углекислого газа в ночное время, это называется метаболизмом толстой кислоты или САМ .
За этими процедурами следуют растения, определенные виды бактерий и водорослей для производства энергии, независимо от среды их обитания. Синтез энергии с использованием углекислого газа и воды в качестве основного источника для получения питательных веществ из воздуха и воды называется фотосинтезом. Это основной процесс для живого существа, которое производит пищу самостоятельно
В этом материале мы рассмотрим существенное различие между тремя типами путей, по которым следуют растения, и несколькими микроорганизмами, и небольшое описание о них.
Сравнительная таблица
| Основа для сравнения | C3 путь | C4 путь | CAM |
|---|---|---|---|
| Определение | Такие растения, чей первый продукт после ассимиляции углерода от солнечного света - это 3-углеродная молекула или 3-фосфоглицериновая кислота для производство энергии называется растениями С3, а путь называется путём С3. Это чаще всего используется растениями. | Растения в тропической зоне преобразуют энергию солнечного света в молекулу углерода С4 или в щавелево-уксусную кислоту, что происходит до цикла С3 и затем он далее преобразуется в энергию, называется растениями С4, а путь называется путём С4. Это более эффективно, чем путь C3. | Растения, которые накапливают энергию от солнца, а затем преобразуют ее в энергию в течение ночи, следуют за CAM или crassulacean кислотой. метаболизм. |
| Клетки участвуют | Клетки мезофилла. | Мезофилловая клетка, пучки оболочек клеток. | И С3, и С4 в одних и тех же клетках мезофилла. |
| пример | Подсолнечник, шпинат, фасоль, рис, хлопок. | Сахарный тростник, сорго и кукуруза. | Кактусы, орхидеи. |
| Можно увидеть в | Все фотосинтезирующие растения. | В тропических растениях | Полузасушливое состояние. |
| Типы растений, использующих этот цикл | Мезофитный, гидрофитный, ксерофитный. | Мезофильных. | Ксерофитные. |
| Фотодыхание | Присутствует в высоком темпе. | Не легко обнаружить. | Обнаруживается днем. |
| Для производства глюкозы | Требуется 12 NADPH и 18 ATP. | Требуется 12 NADPH и 30 ATP. | Требуется 12 NADPH и 39 ATP. |
| Первый стабильный продукт | 3-фосфоглицерат (3-PGA). | Оксалоацетат (ОАА). | Оксалоацетат (ОАА) ночью, 3 ПГА в дневное время. |
| Кальвин циклоператор | В одиночестве. | Наряду с циклом Хэтча и Слэка. | С3 и хэтч и слабый цикл. |
| Оптимальная температура для фотосинтеза | 15-25 ° C | 30-40 ° С | > 40 градусов ° C |
| Карбоксилирующий фермент | RuBP карбоксилаза. | В мезофилле: PEP карбоксилаза. В связке оболочка: RuBP карбоксилаза. | В темноте: ПКП-карбоксилаза. В свете: RUBP карбоксилаза. |
| Соотношение CO2: ATP: NADPH2 | 1: 3: 2 | 1: 5: 2 | 1: 6, 5: 2 |
| Начальный акцептор СО2 | Рибулозо-1, 5-biphophate (RuBP). | Фосфоенолпируват (ПКП). | Фосфоенолпируват (ПКП). |
| Анатомия Кранца | Отсутствует. | Настоящее время. | Отсутствует. |
| Точка компенсации CO2 (промилле) | 30-70. | 6-10. | 0-5 в темноте. |
Определение пути С3 или цикла Кальвина.
Растения С3 известны как растения холодного сезона или умеренные . Они лучше всего растут при оптимальной температуре от 65 до 75 ° F с температурой почвы, подходящей на уровне 40-45 ° F. Эти типы растений демонстрируют меньшую эффективность при высокой температуре .
Основным продуктом растений C3 является 3-углеродная кислота или 3-фосфоглицериновая кислота (PGA) . Это считается первым продуктом при фиксации углекислого газа. Путь C3 проходит в три этапа: карбоксилирование, восстановление и регенерация.
Растения С3 превращаются в СО2 непосредственно в хлоропласте. С помощью рибулозобифосфаткарбоксилазы (RuBPcase) получают две молекулы 3-углеродной кислоты или 3-фосфоглицериновой кислоты . Этот 3-фосфоглицерик оправдывает название пути как C3.
На другом этапе NADPH и ATP фосфорилируют с получением 3-PGA и глюкозы. И затем цикл снова начинается с регенерации RuBP.
Путь C3 является одностадийным процессом, происходящим в хлоропласте. Эта органелла действует как хранилище солнечной энергии. Из общего количества растений, присутствующих на земле, 85 процентов используют этот путь для производства энергии.
Растения С3 могут быть многолетними или однолетними. Они очень белковые, чем растения C4. Примерами однолетних растений С3 являются пшеница, овес и рожь, а многолетние растения включают в себя феску, райграс и фруктовый сад. Растения С3 содержат больше белка, чем растения С4.
Определение пути С4 или пути Люка и Слака.
Растения, особенно в тропическом регионе, следуют по этому пути. Перед циклом Calvin или C3 некоторые растения следуют по пути C4 или Hatch and Slack. Это двухстадийный процесс, в котором получают щавелевоуксусную кислоту (ОАА), которая представляет собой 4-углеродное соединение . Это происходит в клетках оболочки мезофилла и связки, присутствующих в хлоропласте.
Когда 4-углеродное соединение произведено, оно отправляется в ячейку оболочки пучка, здесь молекула 4-углерода дополнительно расщепляется на диоксид углерода и соединение 3-кабона. В конце концов, путь C3 начинает генерировать энергию, где 3-углеродное соединение выступает в качестве предшественника.
Растения С4 также известны как теплые или тропические растения . Они могут быть многолетними или однолетними. Идеальная температура для этих растений - 90-95 ° F. Растения С4 гораздо эффективнее используют азот и собирают углекислый газ из почвы и атмосферы. Содержание белка низкое по сравнению с растениями С3.
Эти растения получили свое название от продукта под названием оксалоацетат, который является 4-угольной кислотой. Примерами многолетних растений C4 являются индийская трава, бермудские острова, просевная трава, крупный синий, а однолетние растения C4 - это суданга, кукуруза, жемчужное просо.
Определение растений CAM
Примечательное замечание, которое отличает этот процесс от вышеупомянутых двух, заключается в том, что при этом типе фотосинтеза организм поглощает энергию солнечного света в дневное время и использует эту энергию в ночное время для ассимиляции углекислого газа.
Это своего рода адаптация во время периодической засухи. Этот процесс допускает обмен газами в ночное время, когда температура воздуха ниже, и происходит потеря водяного пара.
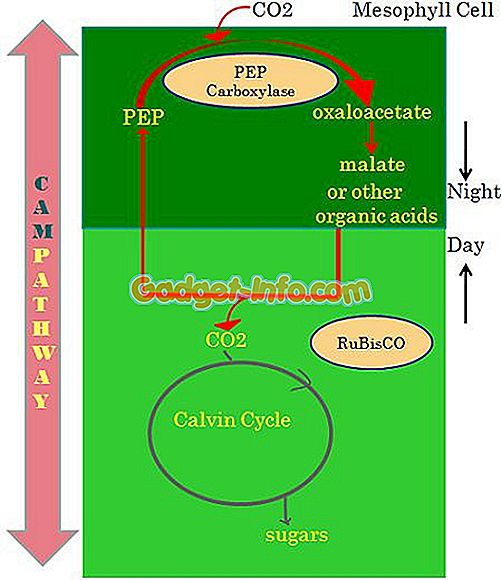
Около 10% сосудистых растений адаптированы к фотосинтезу САМ, но в основном обнаруживаются у растений, выращенных в засушливых районах. Растения, такие как кактус и молочай, являются примерами. Даже орхидеи и бромелии приспособили этот путь из-за нерегулярного водоснабжения.
В дневное время малат декарбоксилируется для обеспечения CO2 для фиксации цикла Бенсона-Кальвина в закрытых устьицах. Главной особенностью растений CAM является ассимиляция ночью CO2 в яблочную кислоту, хранящуюся в вакуоле. PEP карбоксилаза играет основную роль в производстве малата.
Основные отличия растений C3, C4 и CAM.
Выше мы обсудим процедуру получения энергии этих различных типов, ниже мы обсудим ключевые различия между тремя:
- Путь C3 или растения C3 можно определить как те виды растений, у которых первым продуктом после ассимиляции углерода от солнечного света является 3-углеродная молекула или 3-фосфоглицериновая кислота для производства энергии. Это наиболее часто используется растениями; В то время как растения в тропической зоне преобразуют энергию солнечного света в молекулу углерода С4 или щавелевоуксусную кислоту, этот цикл происходит до цикла С3, а затем с помощью ферментов он осуществляет дальнейший процесс получения питательных веществ, называется растениями С4, а путь называется как путь С4. Этот путь более эффективен, чем путь C3. С другой стороны, растения, которые накапливают энергию солнца в дневное время, а затем преобразуют ее в энергию ночью, следуют за метаболизмом CAM или crassulacean кислотой .
- Клетки, участвующие в пути C3, представляют собой клетки мезофилла, а клетки, участвующие в пути C4, представляют собой клетки мезофилла, клетки оболочки пучка, но CAM следует за C3 и C4 в одних и тех же клетках мезофилла.
- Примером С3 являются Подсолнечник, Шпинат, Бобы, Рис, Хлопок, тогда как примером растений С4 является Сахарный тростник, Сорго и Кукуруза, а Кактусы, орхидеи, являются примерами растений САМ.
- C3 можно увидеть во всех фотосинтезирующих растениях, в то время как за C4 следуют тропические растения, а CAM - полузасушливые растения.
- Типы растений, использующих цикл C3, являются мезофитными, гидрофитными, ксерофитными, но C4 следует за мезофитными растениями, а Xerophytic следует за CAM.
- Фотодыхание присутствует с большей скоростью, но его трудно обнаружить в C4 и CAM.
- 12 NADPH и 18 ATP в цикле C3; 12 НАДФН и 30 АТФ в С4 и 12 НАДФН и 39 АТФ необходимы для производства глюкозы.
- 3-фосфоглицерат (3-PGA) является первым стабильным продуктом пути C3; Оксалоацетат (ОАА) для пути С4 и Оксалоацетат (ОАА) ночью, 3 PGA в дневное время в САМ.
- Оптимальная температура для фотосинтеза в С3 составляет 15-25 ° С; 30-40 ° C в растениях C4 и> 40 ° C в CAM
- Карбоксилирующий фермент представляет собой RuBP-карбоксилазу в растениях C3, но в растениях C4 это PEP-карбоксилаза (в мезофилле) и RuBP-карбоксилазу (в оболочке пучка), тогда как в CAM это PEP-карбоксилаза (в темноте) и RuBP-карбоксилаза (в свете).
- Соотношение CO2: ATP: NADPH2 1: 3: 2 в C3, 1: 5: 2 в C4 и 1: 6, 5: 2 в CAM.
- Исходным акцептором CO2 является рибулозо-1, 5-бифосфат (RuBP) в пути C3 и фосфоенолпируват (PEP) в C4 и CAM.
- Анатомия Кранца присутствует только в пути С4 и отсутствует у растений С3 и САМ.
- Точка компенсации CO2 (ppm) составляет 30-70 для установки C3; 6-10 в C4 растениях и 0-5 в темноте в CAM.
Вывод
Мы все знаем о том, что растения готовят пищу в процессе фотосинтеза. Они превращают атмосферный углекислый газ в растительную пищу или энергию (глюкозу). Но поскольку растения растут в разных местах обитания, они имеют разные атмосферные и климатические условия; они отличаются в процессе получения энергии.
Как и в случае, когда пути C4 и CAM являются двумя адаптациями, возникшими в результате естественного отбора, для выживания растений с высокой температурой и засушливой области. Таким образом, мы можем сказать, что это три различных биохимических метода растений для получения энергии, и C3 является наиболее распространенным среди них.
Читайте также: