
Путь энтнера дудорова кратко
Обновлено: 05.07.2024
Сущность энергетического обмена заключается в обеспечении организма энергией, необходимой для проявления жизни. Как уже было отмечено выше, основным источником энергии служит солнечный свет, его энергию улавливают с помощью фотосинтеза растения и фотосинтезирующие бактерии, преобразуя ее в энергию химических структур – глюкозы и других органических соединений. В последующем энергия этих соединений мобилизуется с помощью реакций окисления-восстановления и консервируется в форме АТФ. Молекулы АТФ синтезируются в результате переноса электрона от его первичного донора до конечного акцептора. В зависимости от того, что является конечным акцептором электронов, различают аэробное и анаэробное дыхание. При аэробном дыхании конечным акцептором электронов служит молекулярный кислород (О2), а при анаэробном – различные неорганические соединения: NO3 – , SО4 2– , SO3 2– . Таким образом, энергия мобилизуется в реакциях окисления и восстановления. Окисление – отдача электронов, восстановление – присоединение электронов. Когда отнятие пары электронов или атомов водорода от органического субстрата сопряжено с восстановлением кислорода до воды, это сопровождается значительным изменением свободной энергии (?G 0 ). Оно примерно равно изменению энергии при сжигании одной молекулы водорода (?G 0 = – 57,04 ккал). Перенос электронов по цепи позволяет этой энергии выделяться порциями и превращать часть ее в богатые энергией связи АТФ. Чтобы такая цепь переноса действовала, в ней должен существовать градиент способности к окислению. Способность вещества отдавать электрон или присоединять его (т. е. окисляться или восстанавливаться) количественно выражается в виде его окислительно-восстановительного потенциала.
Переносчики электронов в цепи их переноса участвуют в последовательных реакциях с постепенно увеличивающимися значениями ?E'0 (?E'0 – разность между потенциалами двух полуреакций) и увеличением окислительно-восстановительного потенциала.
Принципиальная схема цепи переноса электронов от первичного донора электронов (атома водорода) до конечного его акцептора О2 выглядит так:
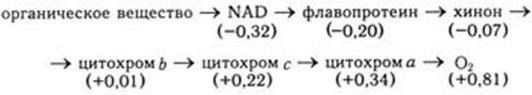
Окислительно-восстановительный потенциал указан в вольтах при стандартных условиях (25 °C, рН = 7,0, все реагенты в концентрации 1,0 М). Однако у бактерий встречаются самые разнообразные варианты этой общей схемы. В связи с этим они по типу дыхания подразделяются на следующие четыре группы:
1) строгие аэробы (размножаются только в присутствии кислорода);
2) микроаэрофилы (нуждаются в уменьшенной концентрации свободного кислорода);
3) факультативные анаэробы (могут потреблять глюкозу и размножаться как в аэробных, так и в анаэробных условиях);
4) строгие анаэробы (размножаются только в бескислородных условиях, т. е. не используют О2 в качестве конечного акцептора электронов).
Максимальная мобилизация энергии из глюкозы происходит при ее окислении через цикл лимонной кислоты (цикл Кребса). Один моль глюкозы С6Н12О6 содержит около 690 ккал (такое количество энергии выделяется при сжигании 180 г глюкозы). На первом этапе потребления глюкозы в отсутствие кислорода (при гликолизе) из одной ее молекулы образуются две молекулы молочной кислоты и синтезируются всего две молекулы АТФ. Каждая молекула АТФ имеет одну богатую энергией (10 ккал) пирофосфатную химическую связь. После расщепления глюкозы до молочной кислоты последняя в присутствии кислорода окисляется и превращается в пировиноградную кислоту, которая далее полностью окисляется через цикл Кребса до СО2 и Н2О. Каждая молекула лактата (пирувата) отдает 6 пар электронов. При переносе каждой пары электронов по цепи переноса часть их энергии используется для образования 3 молекул АТФ (рис. 27).
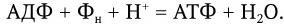
Перемещение протонов по их электрохимическому градиенту с помощью мембранной АТФ-азы служит источником энергии для образования АТФ. Реакция поддерживается градиентом протонов. Однако АТФаза может вызывать и гидролиз АТФ. Это также приведет к перемещению протонов из клетки и созданию их градиента, энергия которого может быть использована для процессов, требующих ее затраты. Некоторые энергообразующие реакции являются общими для аэробных и анаэробных механизмов дыхания. К ним относятся три пути превращения сахаров в основной энергетический метаболит – пировиноградную кислоту: путь Эмбдена – Мейергофа (гликолиз), пентозофосфатный путь (или гексозофосфатный шунт) и путь Энтнера – Дудорова, обнаруженный лишь у некоторых прокариот.
В первом случае (путь Эмбдена – Мейергофа, гликолиз, рис. 29) вначале затрачиваются две молекулы АТФ на образование фруктозо-1,6-дифосфата, который затем расщепляется на фосфоглицериновый альдегид и диоксиацетонофосфат. В результате окисления последних, сопряженного с восстановлением НАД, из каждой образуется по молекуле 1,3-дифосфоглицериновой кислоты. На последующих этапах превращения ее в пировиноградную кислоту происходит так называемое субстратное фосфорилирование, т. е. обе фосфатные группы переносятся на АТФ и, таким образом, на каждую молекулу глюкозы образуются 4 молекулы АТФ. Поскольку две из них затрачиваются на начальных этапах превращения глюкозы, общий выход энергии составляет 2 молекулы АТФ на моль глюкозы.
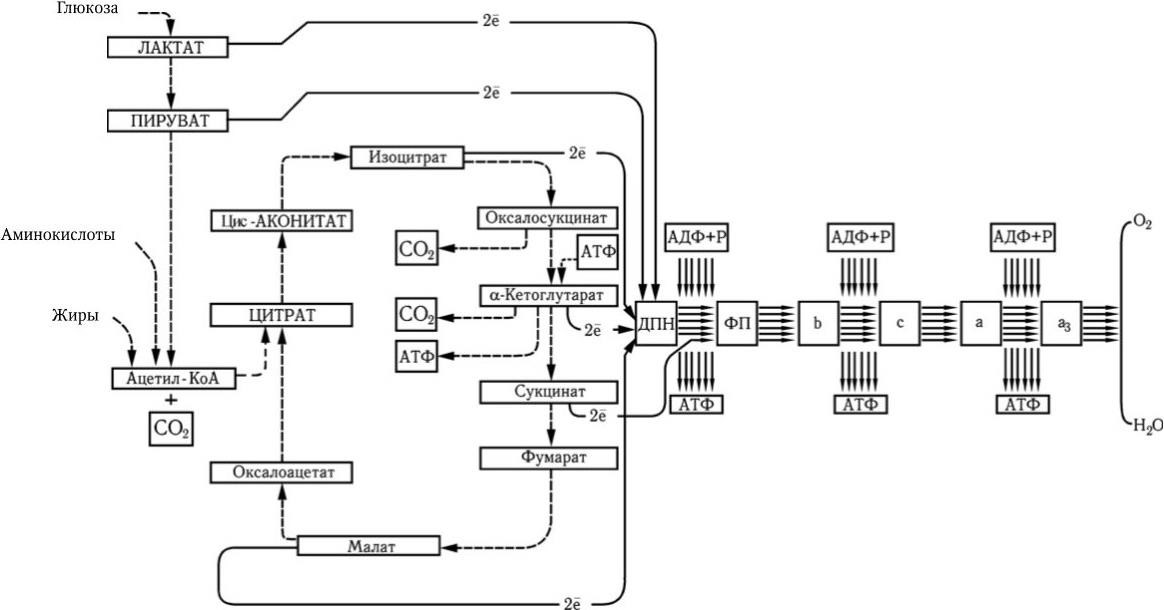
Рис. 27
. Цикл Кребса, или цикл трикарбоновых кислот (ЦТК) (по А. Ленингеру)
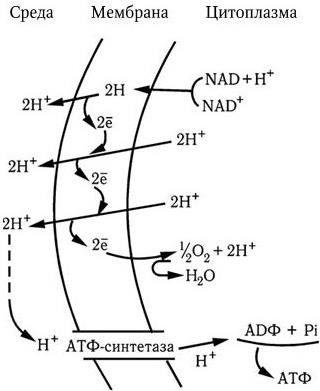
Рис. 28
. Энергизация мембраны. Объяснение в тексте
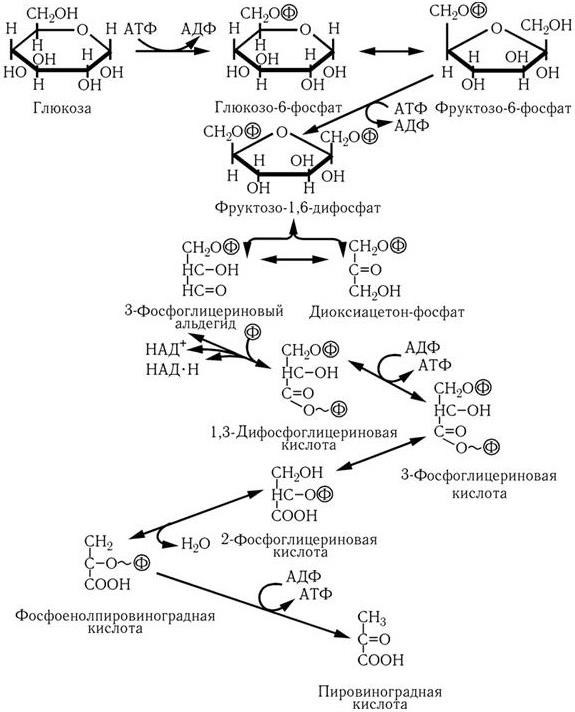
Рис. 29
. Путь Эмбдена–Мейергофа: превращение глюкозы в пировиноградную кислоту
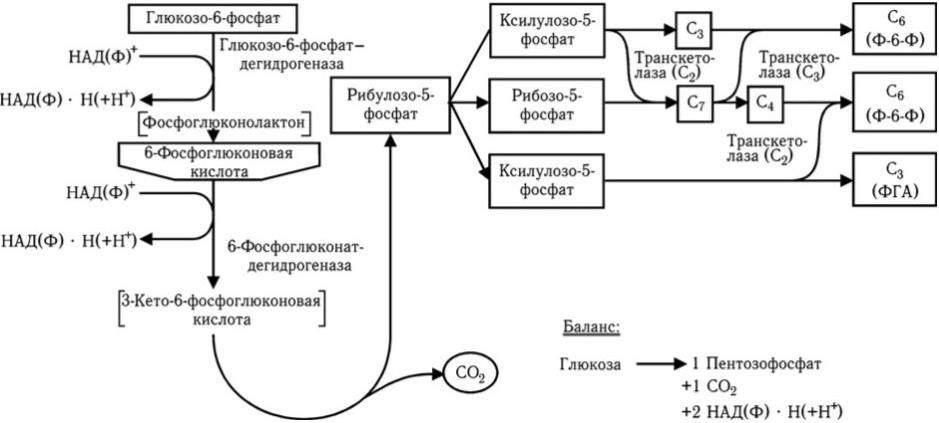
Рис. 30
. Пентозофосфатный путь окисления глюкозы
Пентозофосфатный путь (рис. 30) обеспечивает окисление одного из углеродных атомов глюкозы и не приводит непосредственно к образованию пировиноградной кислоты. Он представляет сложный цикл, при прохождении через который шести молекул происходит полное окисление одной молекулы глюкозо-6-фосфата до СО2 и восстановление шести молекул НАДФ + в НАДФ • Н. Значение этого пути потребления глюкозы заключается в том, что он обеспечивает образование рибозо5-фосфата, необходимого для синтеза нуклеиновых кислот, и большей части НАДФ • Н, нужного для многих биосинтетических реакций.
В случае превращения глюкозы по пути Энтнера – Дудорова (рис. 31) образуется промежуточный продукт, характерный только для этого пути, – 2-кето-3-дезокси-6-фосфоглюконовая кислота, которая далее расщепляется на молекулу пировиноградной кислоты и молекулу 3-фосфоглицеринового альдегида. Последний подвергается дальнейшему превращению по пути Эмбдена – Мейергофа в пировиноградную кислоту. В результате из одной молекулы глюкозы образуются две молекулы пировиноградной кислоты, одна молекула АТФ и две молекулы НАДФ • Н.
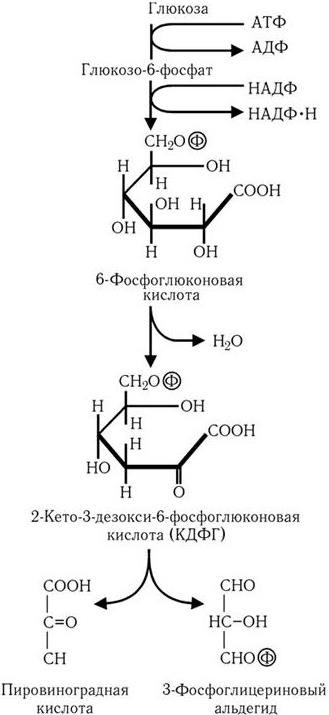
Рис. 31
. Путь Энтнера–Дудорова: превращение глюкозы в пировиноградную кислоту и 3-фосфоглицериновый альдегид
Путь Эмбдена – Мейергофа наиболее широко используется различными бактериями при потреблении глюкозы. От образующегося при этом конечного продукта – пировиноградной кислоты, а также от таких промежуточных продуктов, как эритрозо-4-фосфат и рибозо-5-фосфат, идут различные метаболические пути синтеза двадцати аминокислот (рис. 32). Общая схема обмена веществ у микроорганизмов, обладающих аэробным дыханием и потребляющих гексозы, показана на рис. 33. Поскольку в аэробных условиях высвобождается гораздо больше энергии, чем при брожении, некоторые бактерии осуществляют такой тип дыхания, при котором акцептором водорода (электронов) является связанный кислород. Его носители – нитраты (нитратное дыхание) или сульфаты (сульфатное дыхание). При этом за счет водорода окисляемого субстрата нитраты восстанавливаются до молекулярного азота, а сульфаты – до H2S (рис. 34). Способность таких бактерий переносить электроны на нитраты и сульфаты связана с наличием у них цитохромов и системы переноса электронов. Это позволяет им осуществлять достаточно полное окисление субстрата и получать таким путем гораздо больше энергии, чем при брожении.
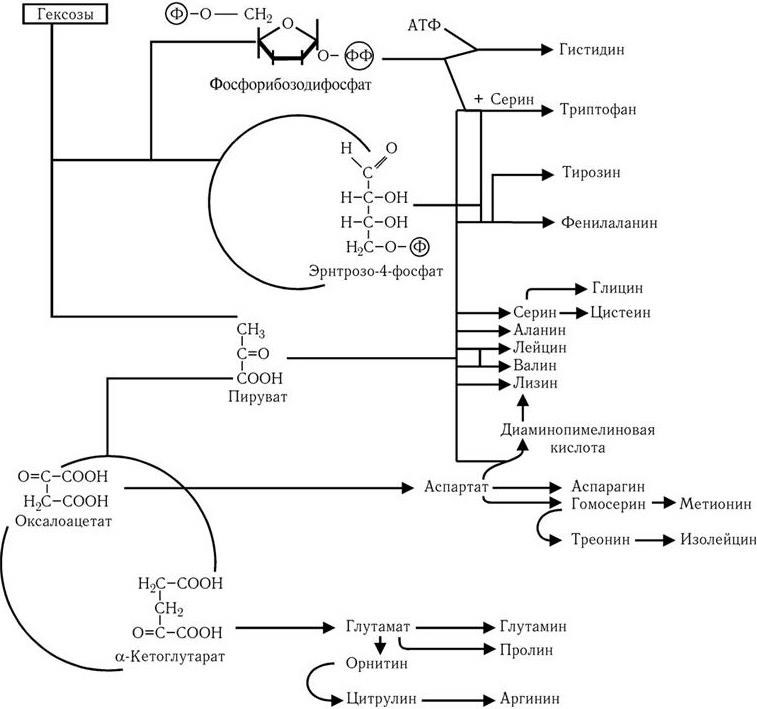
Рис. 32
. Пути образования двадцати аминокислот, необходимых для синтеза белков, из промежуточных продуктов обмена (по Г. Шлегелю)
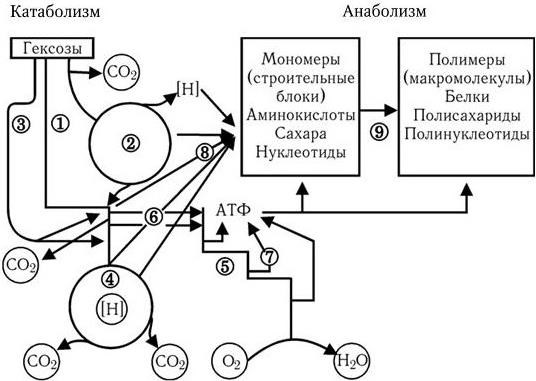
Рис. 33
. Схема обмена веществ у микроорганизмов, потребляющих О2 и гексозы (по Г. Шлегелю):
1 – ФДФ-путь; 2 – ПФ-путь; 3 – КДФГ-путь; 4 – ЦТК; 5 – дыхательная цепь; 6 – фосфорилирование на уровне субстрата; 7 – окислительное фосфорилирование в дыхательной цепи; 8 – синтез мономеров; 9 – синтез полимеров
Поскольку большинство микроорганизмов в качестве источника энергии использует углеводы, и в первую очередь глюкозу, рассмотрим основные пути ее расщепления или катаболизма.
У бактерий возможны три пути катаболизма глюкозы:
1) гликолиз, или фруктозодифосфатный путь, или путь Эмбдена – Мейергофа – Парнаса;
2) окислительный пентозофосфатный путь, или гексозомонофосфатный путь, или путь Варбурга – Диккенса – Хореккера;
3) 2-кето-3-дезокси-6-фосфоглюконатный путь (КДФГ-путь), или путь Энтнера – Дудорова.
Следует отметить, что все перечисленные пути катаболизма глюкозы у микроорганизмов могут протекать при разных типах энергетического метаболизма (аэробное дыхание, анаэробное дыхание, брожение). Все пути катаболизма начинаются с того, что глюкоза, поступившая в клетку, сначала фосфорилируется при участии фермента гексокиназы и АТФ как донора фосфата. Образуется глюкозо-6-фосфат, который представляет метаболически активную форму глюкозы в клетке и служит исходным соединением для любого из трех путей катаболизма углеводов. Пути расщепления глюкозы состоят из многих биохимических реакций, каждая из которых катализируется специфическим ферментом.
Наиболее распространенным путем катаболизма глюкозы у многих микроорганизмов является гликолиз (рис.1). При этом глюкозо-6-фосфат изомеризуется с помощью глюкозофосфатизомеразы и фосфорилируется далее в фруктозо-1,6-дифосфат, который затем расщепляется на 3-фосфоглицериновый альдегид (3-ФГА) и фосфодиоксиацетон. Последний под действием фермента триозофосфатизомеразы превращается в 3-ФГА. Таким образом, из одной молекулы глюкозы образуются две молекулы 3-ФГА. На эти реакции превращения глюкозы в 3-ФГА затрачивается энергия двух молекул АТФ. Далее происходит окисление каждой молекулы 3-ФГА до 1,3-дифосфоглицериновой кислоты (1,3-ФГК). 1,3-ФГК – высокоэнергетическое соединение, содержащее макроэргическую фосфатную связь; оно реагирует с АДФ (фермент фосфоглицераткиназа), отдавая высокоэнергетическую фосфатную группу, в результате чего синтезируется молекула АТФ. Таким образом, энергия, освободившаяся при окислении 3-ФГА, путем субстратного фосфорилирования оказывается аккумулированной в молекуле АТФ. Образуется 3-фосфоглицериновая кислота (3-ФГК):

Далее 3-ФГК под действием фермента фосфоглицеромутазы превращается в 2-ФГК, из которой в результате отщепления воды образуется фосфоенолпировиноградная кислота (ФЕП). Это также высокоэнергетический фосфат, с которого богатая энергией фосфатная группа переносится пируваткиназой на АДФ, образуется молекула АТФ и пировиноградная кислота (ПВК). Это второе фосфорилирование на уровне субстрата:

Таким образом, при распаде одной молекулы глюкозы образуется четыре молекулы АТФ, в которых аккумулируется освободившаяся энергия. Поскольку в начале процесса на активирование глюкозы были затрачены две молекулы АТФ, чистый выход АТФ на одну молекулу глюкозы составляет две молекулы. Суммарное уравнение гликолиза можно записать следующим образом:
Пентозофосфатный путь расщепления углеводов характерен для некоторых представителей семейства Enterobacteriaceae, а также для гетероферментативных молочнокислых бактерий и некоторых маслянокислых бактерий. В этом цикле глюкозо-6-фосфат, образующийся путем активирования глюкозы молекулой АТФ, превращается через ряд промежуточных реакций в 6-фосфоглюконовую кислоту, которая подвергается окислению и декарбоксилированию с образованием рибулозо-5-фосфата, СО2 и НАДФ · Н2 . Рибулозо-5-фосфат включается в сложный цикл, приводящий к образованию из трех его молекул двух молекул глюкозо-6-фосфата и одной молекулы 3-фосфоглицеринового альдегида. Глюкозо-6-фосфат может снова включаться в цикл, а 3-ФГА может быть превращен в пировиноградную кислоту.
С энергетической точки зрения этот путь катаболизма углеводов в 2 раза менее эффективен, чем гликолитический, так как при окислении одной молекулы глюкозы образуется только одна молекула АТФ. Однако большое значение этого пути в том, что он обеспечивает клетки бактерий пентозами (рибулозо-5-фосфатом), которые являются предшественниками нуклеотидов и нуклеиновых кислот. Кроме того, в этом цикле образуются две молекулы НАДФ · Н2, которые необходимы клетке для восстановительных реакций биосинтеза.
Путь Энтнера – Дудорова встречается у прокариот реже других. Он характерен в основном для псевдомонад и уксуснокислых бактерий. От пентозофосфатного пути он отличается тем, что 6-фосфоглюконовая кислота превращается в пировиноградную кислоту и 3-ФГА. Последний может превращаться в пировиноградную кислоту. Из одной молекулы глюкозы при функционировании этого пути синтезируется одна молекула АТФ, по одной молекуле НАДФ · Н2 и НАД · Н2. Следует подчеркнуть, что путь Энтнера – Дудорова является самым кратчайшим механизмом расщепления углеводов до пировиноградной кислоты.
Сравнительная характеристика различных путей катаболизма глюкозы представлена на рис. 1. Рассмотрев пути катаболизма глюкозы, мы можем заключить, что важнейшим продуктом, образующимся в них, является пировиноградная кислота, которая подвергается дальнейшим превращениям. Пируват занимает центральное положение в метаболизме клеток и может служить предшественником многих продуктов.

Рис. 1. Схема путей катаболизма глюкозы в клетках прокариот: А – гликолиз; Б – путь Энтнера-Дудорова; В – пентозофосфатный путь
КДФГ—ПУТЬ – путь расщепления гексоз, известный только у бактерий. Заключается в образовании из глюкозы характерного промежуточного соединения 2–кето–3–дезокси–6–фосфоглюконата с последующим превращением его до двух молекул пирувата. На одну молекулу глюкозы в результате этого пути образуется две молекулы пирувата, одна молекула АТФ и по одной молекуле восстановленных НАДФ и НАД.
Смотреть что такое "Энтнера-Дудорова путь" в других словарях:
качалка — лаб. приспособление для механического встряхивания колб с целью лучшего перемешивания реакционных смесей. В микробиол. практике К. обычно используются для улучшения аэрации среды при выращивании аэробных микроорганизмов. кдфг–путь – см. Энтнера… … Словарь микробиологии
Анаэробные организмы — Аэробные и анаэробные бактерии предварительно идентифицируются в жидкой питательной среде по градиенту концентрации O2: 1. Облигатные аэробные (нуждающиеся в кислороде) бактерии в основном собираются в верхней части пробирки, чтобы поглощать… … Википедия
Актиномицеты — Streptomyces sp … Википедия
Археи — Halobacteria, штамм NRC 1, каждая клетка около 5 мкм длиной … Википедия
Лучистые грибки — ? Актиномицеты Научная классификация Царство: Бактерии Отдел: Актиномицеты … Википедия
Путь Энтнера-Дудорова (КДФГ-путь окисления углеводов) — метаболический путь превращения глюкозы до пирувата альтернативный гликолиза, характерный для некоторых прокариот, преимущественно грамотрицательных бактерий. Названный в честь ученых Майкла Дудорова и Натана Энтнера, что открыли этот путь в 1952 году в бактерии Pseudomonas saccharophila. Путь Энтнера-Дудорова примитивный и менее эффективный чем гликолиз.
Реакции пути Энтнера-Дудорова
Путь Энтнера-Дудорова начинается с фосфорилирования глюкозы в шестом положении. Образован глюкозо-6-фосфат как и в пентозофосфатному пути окисляется до 6-фосфоглюконат в две стадии. Фермент глюкозо-6-фосфатдегидрогеназы катализирует образование 6-фосфоглюконо-δ-лактона. Акцептором электронов в этой реакции в некоторых микроорганизмов может выступать только НАДФ +, тогда как в других и НАДФ + и НАД +. Образованный лактон гидролизуется лактоназою до 6 фосфоглюконат.
6-фосфоглюконат подлежит дегидратации, которую катализирует фермент уникальный для этого метаболического пути — 6-фосфоглюконатдегидратаза (другое название дегидратазы Энтнера-Дурдорова, EDD). Продуктом реакции является 2-кето-3-дезокси-6-фосфоглюконат (КДФГ) — ключевой метаболит пути Энтнера-Дудорова. КДФГ под действием КДФГ-альдолазы расщепляется до одной молекулы пирувата и одной молекулы глицеральдегид-3-фосфата, который далее превращается в реакциях аналогичных гликолитических, во время которых происходит фосфорилирования двух молекул АДФ. Таким образом, на один моль глюкозы в пути Энтнера-Дудорова восстанавливается один моль НАД +, один моль НАДФ + (или два моли НАД +) и синтезируется один моль АТФ (в отличие от гликолиза, где выход АТФ составляет 2 моли на моль глюкозы).
Распространение и модификации
Путь Энтнера-Дудорова значительно менее распространен в природе чем гликолиз, он встречается преимущественно в грамотрицательных бактерий, в частности родов Rhizobium, Pseudomonas, Azotobacter, Agrobacterium. Редким примером грамм-положительной бактерии, использует этот путь Enterococcus faecalis.
Путь Энтнера-Дудорова образует ядро центрального метаболизма и является единственным способом окисления углеводов в анаэробного организма Zymomonas mobilis, у которого отсутствует гликолиз и пентозофосфатный путь. Образован в этом пути пируват вступает в реакции спиртового брожения. Такой путь метаболизма глюкозы очень неэффективен (выход только 1 моль АТФ на 1 моль глюкозы), что компенсируется высокими темпами процесса. Путь Энтнера-Дудорова также протекает линейно и является центральным в катаболизме углеводов в возбудителя гонореи Neisseria gonorrhoeae.
Во многих аэробных организмов, в которых отсутствует фермент гликолиза фосфофруктокиназы, в частности представителей рода Pseudomonas, путь Энтнера-Дудорова протекает в циклической форме: образован в альдолазний реакции глицеральдегид-3-фосфат в реакциях глюконеогенеза превращается обратно в 6-фосфоглюконат.
Читайте также: