
Проблема клеточной пролиферации в медицине кратко
Обновлено: 04.07.2024
Антонова Л.В. 1 Матвеева В.Г. 1 Понасенко А.В. 1 Головкин А.С. 1 Артымук Н.В. 2 Тришкин А.Г. 2 Бикметова Е.С. 2
2 ГОУ ВПО Кемеровская государственная медицинская академия Минздравсоцразвития России, Кемерово, Россия
Цель исследования - изучить динамику пролиферативной активности и апоптотических изменений эндотелиальных клеток (ЭК) человека в условиях гипоксии и последующей реоксигенации. Различные модели гипоксии ((10% О2, 5% СО2, 85% Ar) и (5% О2, 5% СО2, 90% Ar)) создавали в условиях мультигазового биореакторного комплекса в непрерывном 3-х суточном режиме с использованием автоматической коррекции газового состава через каждые 2 минуты и поддержанием температуры в колбах биореактора 37ºС. Контролем для проводимых исследований явилось культивирование ЭК при 20% О2, 5% СО2 и 37ºС. В ходе эксперимента выявлено, что снижение концентрации кислорода с 20% до 5% привело к увеличению в 9 раз частоты возникавшего некроза ЭК. Через 2 суток культивирования ЭК при 10% О2 и 5%О2 коэффициенты клеточной пролиферации превысили таковые при стандартных условиях культивирования в 1,6 и 1,8 раз, соответственно. Увеличение сроков культивирования ЭК при 10% О2 и 5% О2 способствовало повышению коэффициентов пролиферации в период реоксигенации. Максимум подобных проявлений зафиксирован после 3-дневного культивирования ЭК при 5% О2 и последующей 3-дневной реоксигенации. Коэффициент пролиферации при этом оказался в 1,8 раза выше по сравнению с 3-дневным культивированием ЭК в стандартных условиях.

1. Буравкова Л.Б., Анохина Е.Б. Влияние гипоксии на стромальные клетки-предшественники из костного мозга крыс на ранних этапах культивирования // Бюллетень экспериментальной биологии и медицины. - 2007. - Т.143, №4. - С. 386-389.
2. Влияние гипоксии на длительно культивируемые стромальные клетки-предшественники, выделенные из костного мозга крыс / Е.Б. Анохина, Л.В. Буравкова, А.И. Во- ложин, О.В. Григорьев // Патологическая физиология и экспериментальная терапия. - 2006. - Т.4. - С 26-28.
3. Действие гипоксии и реоксигенации на культивируемые эндотелиальные клетки человека / О.А. Антонова, С.А. Локтионова, О.Н. Шустова, Н.В. Голубева, А.В. Мазуров // Кардиологический вестник. - 2009. - Т. IV (XVI), № 2. - С. 12-18.
4. Повреждение и активация эндотелиальных клеток при гипоксии in vitro / О.А. Антонова, С.А. Локтионова,
Н.В. Голубева, Ю.А. Романов, А.В. Мазуров // Бюллетень экспериментальной биологии и медицины. - 2007. - №144. - С. 384-386.
7. Hypoxia-induced apoptosis in endothelial cells and embrionic stem cells / C.N. Lee, W.F. Cheng, Chang M.C., Y.N. Su, C.A. Chen, F.J. Hsieh // J. Apoptosis. - 2005. - V. 10, №4. - Р. 887-894.
8. Mechanisms of hypoxia-induced endothelial cell death: role of p53 in apoptosis / A. Stempien-Otero, A. Karsan, C.J. Cornejo, H. Xiang, T. Eunson, R.S. Morrison, M. Kay, R. Winn, J. Harlan // J. Biol. Chem. - 1999. - V.274, №12. - Р. 8039-8045.
9. Oxidative stress in cultured cerebral endothelial cells induces chromosomal aberrations, micronuclei, and apoptosis / N. Bresgen, G. Kahlhuber, I. Krizbai et al. // J. Neurosci. Res. - 2003. - № 72. - Р. 327-333.
Гипоксия представляет собой универсальный патологический процесс, который сопровождает и определяет возникновение большинства сердечно-сосудистых заболеваний. Развитие сердечно-сосудистых событий предусматривает как периоды кислородного голодания в ишемизированных участках тканей, так и последующее восстановление кровотока - реперфузию. При этом нарушение кровоснабжения в период гипоксии и последующая реоксигенация приводят к изменениям состояния эндотелиальных клеток (ЭК), которые правильнее рассматривать как систему эндотелиальных клеток, способную к саморегуляции в ответ на изменения условий среды [5]. Известно, что временная гипоксия способна вызывать активацию эндотелия [4, 10], а резкое повышение содержания кислорода (реоксигенация), может оказывать как повреждающее [3, 9], так и активирующее [6, 10] действие на эндотелиальные и прочие типы клеток. Возможность стимуляции апоптотических изменений культивируемых эндотелиоцитов в ответ на гипоксию и реоксигенацию освещалась многими авторами. К сожалению, данные, получаемые при изучении влияния различных концентраций кислорода на пролиферативную активность и апоптоз эндотелиальных клеток малочисленны и противоречивы в силу отличия режимов проводимых экспериментов, что приводит к необходимости дальнейшего изучения данного вопроса.
Цель исследования - изучить динамику пролиферативной активности и апоптотических изменений эндотелиальных клеток человека в условиях гипоксии и последующей реоксигенации.
Материал и методы исследования
Подсчет эндотелиальных клеток после снятия с поверхности культурального пластка 0,5% раствором трипсина (Sigma Aldrich, США) проводили в камере Горяева. Фенотип снятых HUVEC определяли методом проточной цитофлуориметрии на цитофлуориметре FACS Calibur (Becton Dickinson, США) с использованием моноклональных антител CD146 и CD34 (BD Biosciences, США), меченных флуорохромами. Жизнеспособность HUVEC оценивали методом проточной цитофлуориметрии по содержанию клеток в состоянии апоптоза и некроза относительно всей популяции. Для этой цели использовали комбинированную окраску: Annexin V, меченый PE, и ДНК-связывающий краситель 7AAD (BD Biosciences, США). Клетки в состоянии апоптоза определялись за счет высокоафинного связывания Annexin V-РЕ с фосфатидилсерином, появляющегося на внешнем монослое цитоплазматической мембраны. Дополнительная окраска 7-AAD позволила выделить раннюю и позднюю стадию клеточного апоптоза и идентифицировать некроз.
Оставшуюся часть снятой после гипоксии культуры HUVEC высевали в три покрытых коллагеном I типа 96-луночных планшета для проведения МТТ-теста колориметрическим способом с оценкой результатов через 24, 48 и 72 часа после культивирования в условиях СО2-инкубатора при 20% О2, 5% СО2 и 37°С. МТТ-тест проводили с использованием MTT Cell Proliferation Assay Kit (Invitrogen, США). Для определения пролиферативной активности эндотелиальных клеток в условиях гипоксии через 1, 2 и 3 сут из второй колбы биореактора вынимали по одному 96-луночному планшету с HUVEC и проводили МТТ-тест. Один стрип выступал в качестве контроля культуральной среды и не содержал HUVEC. Оценку результатов МТТ-теста осуществляли с помощью вычисления коэффициента пролиферации (КП) по формуле:
где ОПопыт - это оптическая плотность экстракта красителя, поглощенного эндотелиальными клетками в ходе МТТ-теста; ОПконтроль - это оптическая плотность красителя, содержащегося после МТТ-теста в лунках с культуральной средой без клеток.
Анализируемые показатели имели ненормальное распределение, поэтому достоверность различий определяли с помощью непараметрических критериев. Для оценки достоверности различий трех и более показателей внутри одной группы использовали критерий Фридмана, при проведении попарных сравнений - критерий Вилкоксона с поправкой Бонферрони. При сравнении двух несвязанных групп (показатели при нормоксии и гипоксии) использовали U-критерий Манна-Уитни, при сравнении трех несвязанных групп (эффекты гипоксии и реоксигенации по сравнению с нормоксией) - критерий Крискала - Уоллеса. Данные представляли как медиана и квартильный размах (Ме (25%; 75%)). При всех видах статистического анализа учитывался уровень статистической значимости 95% (р + , CD 34 - . В процессе 3-дневного культивирования HUVEC при 20% О2 клеточный фенотип и количество жизнеспособных клеток достоверно не менялись. Снижение концентрации кислорода от 10 до 5% в процессе культивирования также не привело к изменению клеточного фенотипа и процента жизнеспособных эндотелиоцитов (93,5% при 20% О2 до 92,9% - при 5% О2), однако способствовало увеличению процента клеток, пребывавших в состоянии некроза за счет практически полного исчезновения HUVEC в состоянии раннего апоптоза (рис. 1).

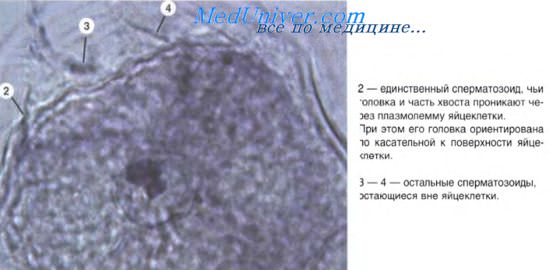
Рис. 1. Изменение частоты клеточного апоптоза и некроза в зависимости от условий культивирования эндотелиальных клеток: * - p 5
2.1.10. Механизмы развития пролиферации в очаге воспаления
Пролиферация является завершающей фазой развития воспаления, обес-печивающей репаративную регенерацию тканей на месте очага аль-
терации. Пролиферация развивается с самого начала воспаления наряду с яв-лениями альтерации и экссудации.
Восстановление и замещение поврежденных тканей начинается с выхода из сосудов молекул фибриногена и образования фибрина, который формиру-ет своеобразную сетку, каркас для последующего клеточного размножения. Уже по этому каркасу распределяются в очаге репарации быстро образую-щиеся фибробласты. Деление, рост и перемещение фибробластов возможны только после их связывания с фибрином или коллагеновыми волокнами. Эта связь обеспечивается особым белком – фибронектином. Размножение фиб-робластов начинается по периферии зоны воспаления, обеспечивая формиро-вание фибробластического барьера. Хемотаксис, активация и пролиферация фибробластов осуществляются под воздействием:
1. Факторов роста фибробластов.
2. Тромбоцитарного фактора роста.
3. Цитокинов - ФНО, ИЛ-1.
6. Трансформирующего фактора роста b.
Сначала фибробласты не зрелые и не обладают достаточной синтетиче-ской активностью. Созреванию предшествует внутренняя структурно-функциональная перестройка фибробластов: гипертрофия ядра и ядрышка, гиперплазия ЭПС, повышение содержания ферментов, особенно щелочной фосфатазы, неспецифической эстеразы, b-глюкуронидазы. Только после пе-рестройки фибробласты начинают синтезировать коллаген, эластин, коллаге-нассоциированные белки и протеогликаны. Коллагеногенез стимулируется следующими биологически активными веществами - ФНО, ИЛ-1, ИЛ-4, фак-тором роста фибробластов, тромбоцитарным фактором роста.
Интенсивно размножающиеся фибробласты продуцируют кислые муко-полисахариды – основной компонент межклеточного вещества соединитель-ной ткани (гиалуроновую кислоту, хондроитинсерную кислоту, глюкозамин, галактозамин). При этом зона воспаления не только инкапсулируется, но и начинаются постепенная миграция клеточных и бесклеточных компонентов соединительной ткани от периферии к центру, формирование соединитель-нотканного остова на месте первичной и вторичной альтерации.
Наряду с фибробластами размножаются и другие тканевые и гематоген-ные клетки. При разрушении базальных мембран сосудов в зоне альтерации происходит миграция клеток эндотелия по градиенту ангиогенных факторов. Просвет новообразующегося капилляра формируется путем слияния внекле-точных пространств соседних эндотелиоцитов. Вокруг новообразующихся капилляров концентрируются тучные клетки, макрофаги, нейтрофилы, кото-рые освобождают биологически активные вещества, способствующие про-лиферации капилляров.
Важнейшими факторами, стимулирующими ангиогенез, являются:
1. Факторы роста фибробластов (основной и кислый).
2. Сосудистый эндотелиальный фактор роста.
3. Трансформирующие факторы роста .
4. Эпидермальный фактор роста.
Фибробласты вместе с вновь образованными сосудами создают грануля-ционную ткань. Это, по существу, молодая соединительная ткань, богатая клетками и тонкостенными капиллярами, петли которых выступают над по-верхностью ткани в виде гранул.
Основными функциями грануляционной ткани являются: защитная – пре-дотвращение влияния факторов окружающей среды на очаг воспаления и ре-паративная – заполнение дефекта и восстановление анатомической и функ-циональной полноценности поврежденных тканей.
Формирование грануляционной ткани не строго обязательно. Это зависит от величины и глубины повреждения. Грануляционная ткань обычно не раз-вивается при заживлении ушибленных кожных ранок или мелких поврежде-ний слизистой оболочки (Кузин М.И., Костюченок Б.М. и соавт., 1990). Гра-нуляционная ткань постепенно превращается в волокнистую ткань, называе-мую рубцом. В рубцовой ткани уменьшается количество сосудов, они запус-тевают, уменьшается количество макрофагов, тучных клеток, снижается ак-тивность фибробластов. Небольшая часть клеточных элементов, распола-гающаяся среди коллагеновых нитей, сохраняет активность. Предполагают, что сохранившие активность тканевые макрофаги принимают участие в рас-сасывании рубцовой ткани и обеспечивают формирование более мягких руб-цов.
Параллельно с созреванием грануляций происходит эпителизация раны. Она начинается в первые часы после повреждения, и уже в течение первых суток образуются 2-4 слоя клеток базального эпителия. Скорость эпителиза-ции обеспечивается следующими процессами: миграцией, делением и диф-ференцировкой клеток. Эпителизация небольших ран осуществляется, в ос-новном, за счет миграции клеток из базального слоя. Раны более крупные эпителизируются за счет миграции и митотического деления клеток базаль-ного слоя, а также дифференцировки регенерирующего эпидермиса. Новый эпителий образует границу между поврежденным и подлежащим слоем, он препятствует обезвоживанию тканей раны, уменьшению в ней электролитов и белков, а также предупреждает инвазию микроорганизмов.
В процессе пролиферации участвуют и органспецифические клеточные элементы органов и тканей. С точки зрения возможностей пролиферации ор-ганспецифических клеточных элементов все органы и ткани могут быть рас-классифицированы на три группы:
К первой группе могут быть отнесены органы и ткани, клеточные элемен-ты которых обладают активной или практически неограниченной пролифе-рацией, достаточной для полного восполнения дефекта структуры в зоне воспаления (эпителий кожи, слизистых оболочек дыхательных путей, слизи-стой желудочно-кишечного тракта, мочеполовой системы; гемопоэтическая ткань и др.).
Ко второй группе относятся ткани с ограниченными регенерационными способностями (сухожилия, хрящи, связки, костная ткань, периферические нервные волокна).
К третьей группе относятся те органы и ткани, где органоспецифические клеточные элементы не способны к пролиферации (сердечная мышца, клетки ЦНС). Основными факторами, регулирующими процессы пролиферации и дифференцировки клеток в очаге воспаления, являются:
1. Факторы роста, продуцируемые макрофагами, лимфоцитами, тромбо-цитами, фибробластами и другими клетками, стимулированными в зоне вос-паления. К ним относятся:
- факторы роста эпидермиса (стимулятор пролиферации и созрева-ния эпителия, стимулятор ангиогенеза);
- трансформирующий фактор роста- (стимулятор ангиогенеза);
- трансформирующий фактор роста- (хемоаттрактант фибробла-стов, стимулятор синтеза коллагена, фибронектина, ангиогенеза, ингибитор протеолиза);
- тромбоцитарный фактор роста (стимулятор миграции, пролифе-рации и синтеза белка в клетках-мишенях, обладает провоспалительным эф-фектом);
- фактор роста эндотелиоцитов;
- фактор роста фибробластов кислый и основной (стимуляторы пролиферации всех клеток сосудистой стенки);
- колониестимулирующие факторы (гранулоцитарный и макрофа-гальный стимуляторы дифференцировки, пролиферации и функциональной активности клеток гранулоцитарного и моноцитарного ряда) - цитокины (ФНО, ИЛ-1, ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-5, ИЛ-6, ИЛ-7) , продуцируемые Т- и В-лимфоцитами, мононуклеарами, тучными клетками, фибробластами, эндоте-лиоцитами, обеспечивают хемотаксис, фиброгенез, ингибируют апоптоз, стимулируют процессы пролиферации в очаге воспаления. Ингибиторами роста для некоторых клеток служат те же цитокины, которые стимулируют пролиферацию других – это ФНО, трансформирующий фактор роста и - интерферон (Зайчик А.Ш., Чурилов Л.П., 1999);
- фактор роста нервов (стимулятор пролиферации, роста, морфоге-неза симпатических нейронов, эпителиальных клеток). Ростовые факторы, взаимодействуя с рецепторами на клетках- мишенях, могут непосредственно стимулировать синтез ДНК в клетках или подготавливать внутриклеточные рецепторы и ферменты к митотической деятельности.
2. Пептид гена, родственного кальцитонину, стимулирует пролифе-рацию эндотелиальных клеток, а субстанция Р индуцирует выработку ФНО в мак-рофагах.
3. Простагландины группы Е потенцируют регенерацию путем усиления кровоснабжения.
4. Кейлоны и антикейлоны, продуцируемые различными клетками, дейст-вуя по принципу обратной связи, могут активировать и угнетать митотиче-ские процессы в очаге воспаления (Бала Ю.М., Лифшиц В.М., Сидельникова В.И., 1988).
5. Полиамины (путресцин, спермидин, спермин), обнаруживаемые во всех клетках млекопитающих, жизненно необходимы для роста и деления клеток.
Они обеспечивают стабилизацию плазматических мембран и суперспи-ральной структуры ДНК, защиту ДНК от действия нуклеаз, стимуляцию транскрипции, метилирование РНК и связывание ее с рибосомами, актива-цию ДНК-лигаз, эндонуклеаз, протеинкиназ и многие другие клеточные про-цессы. Усиленный синтез полиаминов, способствующих пролиферативным процессам, отмечается в очаге альтерации (Березов Т.Т., Федорончук Т.В., 1997).
6. Циклические нуклеотиды: цАМФ ингибирует, а цГМФ активирует про-цессы пролиферации.
Морфологически процесс заживления раны может протекать различно, в зависимости от анатомического субстрата поражения, степени инфицирован-ности, общего состояния организма, характера лечебных мероприятий (Ку-зин М.И., Костюченок Б.М. и соавт., 1990). Однако в любом случае течение раневого процесса отражает один из классических типов заживления:
1. Заживление первичным натяжением.
2. Заживление вторичным натяжением.
3. Заживление под струпом.
Заживление раны первичным натяжением. Такой тип заживления харак-теризуется сращением краев раны без видимой промежуточной ткани, путем соединительнотканной организации раневого канала. Заживление первичным натяжением является наиболее экономичным видом заживления. Для зажив-ления первичным натяжением необходимы следующие условия:
1. Небольшая зона повреждения.
2. Плотное соприкосновение краев раны.
3. Сохранение жизнеспособности краев раны.
4. Отсутствие очагов некроза и гематомы.
5. Асептичность раны.
Заживление раны вторичным натяжением происходит при обширных по-вреждениях тканей, при наличии в ране нежизнеспособных тканей, гематомы и при развитии инфекции в ране. Любой из этих факторов ведет к заживле-нию вторичным натяжением. При различных вариантах течения заживления вторичным натяжением речь идет о заживлении гнойной раны, то есть о за-живлении через нагноение и гранулирование. На 5-6-е сутки после альтера-ции, после отторжения некротизированных клеток в ране появляются ост-ровки грануляций, которые, постепенно разрастаясь, заполняют всю полость раны. Изменения характера грануляций всегда объективно отражают ослож-нения заживления, которые могут наступить под влиянием местных и общих факторов. Реорганизация рубца проявляется активной эпителиизацией раны. Эпителий нарастает на поверхность грануляций в виде голубовато-белой каймы очень медленно. Помимо эпителизации заживлению способствует фе-номен раневой контракции - равномерного концентрического сокращения краев и стенок раны. Этот феномен объясняется появлением в грануляцион-ной ткани в период регенерации фибробластов, обладающих способностью к сокращению.
Заживление раны под струпом характерно для незначительных повреж-дений (ссадины, царапины, небольшие по площади ожоги 1-й и 2-й степени). Раневой процесс начинается свертыванием излившейся крови или только лимфы, которая подсыхает с образованием струпа. Под ним происходит бы-страя регенерация эпидермиса, и струп затем отторгается. Весь процесс длится 3-7 дней. Если заживление под струпом происходит без осложнений, то рана заживает первичным натяжением; если под струпом начинается на-гноение, то заживление идет по типу вторичного натяжения. В ряде случаев может развиться вялотекущее флегмонозное поражение жировой клетчатки, окружающей рану. В такой ситуации необходимы хирургическая обработка раны и удаление струпа (Кузин М.И., Костюченок Б.М. и соавт., 1990).
Эмбриональный гистогенез. Пролиферация клеток. Клеточный рост, миграция и межклеточные взаимодействия.
В развитии низших и высших позвоночных отчетливо прослеживается единая общебиологическая закономерность, выражающаяся в появлении зародышевых листков и обособлении основных зачатков органов и тканей. Процесс образования тканей из материала эмбриональных зачатков составляет суть учения о гистогенезах.
Эмбриональный гистогенез, по определению А.А. Клишова (1984), — это комплекс координированных во времени и пространстве процессов пролиферации, клеточного роста, миграции, межклеточных взаимодействий, дифференциации, детерминации, программированной гибели клеток и некоторых других. Все названные процессы в той или иной мере протекают в зародыше, начиная с самых ранних стадий его развития.
Пролиферация. Основной способ деления тканевых клеток — это митоз. По мере увеличения числа клеток возникают клеточные группы, или популяции, объединенные общностью локализации в составе зародышевых листков (эмбриональных зачатков) и обладающие сходными гистогенетическими потенциями. Клеточный цикл регулируется многочисленными вне- и внутриклеточными механизмами. К внеклеточным относятся влияния на клетку цитокинов, факторов роста, гормональных и нейрогенных стимулов. Роль внутриклеточных регуляторов играют специфические белки цитоплазмы. В течение каждого клеточного цикла существуют несколько критических точек, соответствующих переходу клетки из одного периода цикла в другой. При нарушении внутренней системы контроля клетка под влиянием собственных факторов регуляции элиминируется апоптозом, либо на некоторое время задерживается в одном из периодов цикла.
Метод радиографического анализа клеточных циклов в различных тканях выявил особенности соотношения клеточной репродукции и дифференцировки. Например, если в тканях (кроветворные ткани, эпидермис) имеется постоянный фонд пролиферирующих клеток, за счет которых обеспечивается непрерывное возникновение новых клеток взамен погибающих, то эти ткани относятся к обновляющимся. Другие ткани, например, некоторые соединительные, характеризуются тем, что в них увеличение количества клеток происходит параллельно с их дифференцировкой, клетки в этих тканях характеризуются низкой митотической активностью. Это растущие ткани. Наконец, нервная ткань характеризуется тем, что все основные процессы репродукции заканчиваются в период эмбрионального гистогенеза (когда формируется основной запас стволовых клеток, достаточный для последующего развития ткани). Поэтому она отнесена к стабильным (стационарным) тканям. Продолжительность жизни клеток в обновляющихся, растущих и стабильных тканях разная.
Наряду с обновлением клеточной популяции, в самих клетках постоянно наблюдается обновление внутриклеточных структур (внутриклеточная физиологическая регенерация).
Клеточный рост, миграция и межклеточные взаимодействия. Рост клеток проявляется в изменении их размеров и формы. При усилении функциональной активности и внутриклеточных биосинтезов наблюдается увеличение объема клетки. Если объем клетки превышает некую норму, то говорят о ее гипертрофии, и наоборот, при снижении функциональной активности происходит уменьшение объема клетки, а при переходе некоторых нормативных параметров возникает атрофия клетки. Рост клетки не беспределен и определяется оптимальным ядерно-цитоплазменным отношением.
Важное значение для гистогенеза имеют процессы перемещения клеток. Миграция клеток наиболее характерна для периода гаструляции. Однако и в период гисто- и органогенеза происходят перемещения клеточных масс (например, смещения миобластов из миотомов в места закладки скелетных мышц; движение клеток из нервного гребня с образованием спинномозговых ганглиев и нервных сплетений, миграция гоноцитов и т. д.). Миграция осуществляется с помощью нескольких механизмов. Так, различают хемотаксис — движение клеток в направлении градиента концентрации какого-либо химического агента (перемещения спермиев к яйцеклетке, предшественников Т-лимфоцитов из костного мозга в закладку тимуса).
Гаптотаксис — механизм перемещения клеток по градиенту концентрации адгезионной молекулы (движение клеток протока пронефроса у амфибий по градиенту щелочной фосфатазы на поверхности мезодермы). Контактное ориентирование — когда в какой-либо преграде остается один канал для перемещения (описан у рыб при образовании плавников).
Контактное ингибирование — этот способ перемещения наблюдается у клеток нервного гребня. Суть способа заключается в том, что при образовании ламеллоподии одной клеткой и контакта ее с другой клеткой, ламеллоподия прекращает рост и постепенно исчезает, но в другой части мигрирующей клетки при этом формируется новая ламеллоподия.
В процессе миграции клеток важную роль играют межклеточные взаимодействия. Существует несколько механизмов такого взаимодействия (контактного и дистантного). Выделяется большая группа молекул клеточной адгезии (МКА). Так, кадгерины — это Са2+-зависимые МКА, отвечают за межклеточные контакты при образовании тканей, за формообразование и др. В молекуле кадгерина различают внеклеточный, трансмембранный и внутриклеточный домены. Например, внеклеточный домен ответственен за адгезию клеток с одинаковыми кадгеринами, а внутриклеточный — за форму клетки. Другой класс МКА — это иммуноглобулиновое суперсемейство Са2+-независимых МКА, обеспечивающих, например, адгезию аксонов к сарколемме мышечных волокон, или миграцию нейробластов вдоль радиальных глиоцитов в закладке коры большого мозга и др. Следующий класс МКА — это мембранные ферменты — гликозилтранферазы. Последние по типу "ключ-замок" соединяются с углеводными субстратами — гликозаминогликанами надмембранного комплекса клетки, осуществляя таким образом прочное сцепление клеток.
Кроме механизмов межклеточного взаимодействия, существуют механизмы взаимодействия клеток с субстратом. Они включают формирование рецепторов клетки к молекулам внеклеточного матрикса. К последним относят производные клеток, среди которых наиболее изученными адгезионными молекулами являются коллаген, фибронектин, ламинин, тенасцин и некоторые другие. Коллагены, среди которых различают несколько десятков типов, входят в состав межклеточного вещества рыхлой волокнистой соединительной ткани, базальной мембраны и пр. Фибронектин, секретируемый клетками, является связывающей молекулой между мигрирующей клеткой и межклеточным матриксом. Ламинин — компонент базальной мембраны, также связывает мигрирующие клетки с межклеточным матриксом (справедливо по отношению к эпителиоцитам и нейробластам).
Для осуществления связи мигрирующих клеток с межклеточным матриксом клетки формируют специфические рецепторы. К ним относятся, например, синдекан, который обеспечивает контакт эпителиоцита с базальной мембраной за счет сцепления с молекулами фибронектина и коллагена. Интегрины клеточных поверхностей связывают с внеклеточной стороны молекулы внеклеточного матрикса, а внутри клетки — белки цитоскелета (например, актиновые микрофиламенты). Так возникает связь внутри- и внеклеточных структур, что позволяет клетке использовать для перемещения собственный сократительный аппарат. Наконец, существует большая группа молекул, формирующих клеточные контакты, осуществляющие коммуникацию между клетками (щелевые контакты), механическую связь (десмосомы, плотные контакты).
Дистантные межклеточные взаимодействия осуществляются путем секреции гормонов и факторов роста (ФР). Последние — это вещества, оказывающие стимулирующее влияние на пролиферацию и дифференцировку клеток и тканей. К ним относятся, например, ФР, полученный из тромбоцитов и влияющий на переход клеток в фазу размножения (гладких миоцитов, фибробластов, глиоцитов); эпидермальный ФР — стимулирует пролиферацию эпителиоцитов, производных эктодермы; ФР фибробластов — стимулирует пролиферацию фибробластов. Особо выделяется большая группа пептидов (соматотропины, соматомедины, инсулин, лактоген), влияющих на развитие клеток плода.
Факторы, тормозящие пролиферацию и дифференцировку клеток, также принимают кооперативное участие в процессах развития клеток и тканей. К ним относятся, например, бета-интерферон и трансформирующий ФР. Последний, однако, в отношении разных клеточных типов действует по-разному: блокирует размножение многих типов эпителиоцитов, но стимулирует размножение соединительнотканных клеток.
- Вернуться в оглавление раздела "гистология"
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.

В обзоре представлены процессы клеточной пролиферации в сосудистой стенке человека и экспериментальных животных при формировании атеросклеротической бляшки. Показаны типы активно пролиферирующих клеток — лимфоциты, макрофаги, эндотелиоциты — и выявлены зоны в сосудистой стенке, где эта пролиферация происходит. Определены факторы, которые способствуют и препятствуют клеточной пролиферации при росте атеросклеротической бляшки. В обзоре рассмотрены все стадии формирования атеросклеротических поражений — начиная от нормальных участков и липидных пятен и кончая выраженными фиброзными бляшками. Установлена связь между клеточной пролиферацией и воспалительным процессом в сосудистой стенке человека. Обсуждается вопрос о роли клеточной пролиферации при дестабилизации атеросклеротической бляшки. Если при атеросклерозе этот процесс до сих пор мало изучен, то при формировании нестабильной атеросклеротической бляшки у человека он совершенно неизвестен. На основании собственных данных сделано заключение о важной роли процессов клеточной пролиферации в формировании нестабильной атеросклеротической бляшки у человека.
Ключевые слова
Полный текст
Введение
Оценка атерогенеза с позиции иммунного воспаления позволила рассматривать кинетику клеток стенки артерий с учетом экспрессии цитокинов и межклеточной кооперации: макрофаг – Т-лимфоцит – гладкомышечная клетка. Экспрессия негранулярными лейкоцитами (моноцитами/макрофагами и лимфоцитами) провоспалительных цитокинов и факторов роста сопровождается пролиферацией клеток. В фундаментальном исследовании А.Н. Восканьянц, В.А. Нагорнева [1] показано, что при атерогенезе в стенке артерий происходит активная пролиферация мононуклеарных клеток на всех стадиях формирования атеросклеротических поражений. Именно пролиферирующие в стенке артерий клетки включаются в реакции иммунного воспаления. Авторы полагают, что клетки, выходящие из деления, составляют пул активированных клеток, участвующих в пара- и аутокринной регуляции экспрессии цитокинов и поддерживают иммунное воспаление по принципу замкнутой реакции с саморегуляцией.
В частности, в этой работе впервые проведенный количественный анализ пролиферирующих клеток при атерогенезе у человека показал, что в среднем в липидных пятнах индекс пролиферации мононуклеаров составил 30 %. Авторы отмечают, что эта тенденция сохраняется в липидных и фиброзных (краевые отделы) бляшках. На основании этих данных высказано предположение, что источником макрофагов, включающихся в реакции иммунного воспаления и участвующих в презентации антигенов Т-клеткам, являются макрофаги, мигрирующие в стенку артерий и пролиферирующие in situ. Кроме этого, авторы продемонстрировали, что индекс пролиферации гладкомышечных клеток (ГМК) в липидных пятнах и липидно-фиброзных бляшках в целом совпадает с данными, отражающими пролиферацию мононуклеаров. Однако в фиброзных бляшках этот показатель резко возрастает, что может быть связано с формированием фиброзной покрышки бляшек, для которой необходимо увеличение пула активированных ГМК, синтезирующих волокнистые структуры, в первую очередь соединительнотканные волокна [2, 3].
Схожую точку зрения высказывают и другие исследователи: циркулирующие в крови моноциты проникают в сосудистую стенку, где дифференцируются в макрофаги и затем активно пролиферируют в очаге атеросклеротического поражения [4, 5].
Факторы, способствующие и препятствующие клеточной пролиферации в сосудистой стенке
Существует точка зрения, что моноциты подвергаются дифференциации и пролиферации под действием факторов, секретируемых поврежденным эндотелием [6]. В частности, воздействие на моноциты лейкотриенов LTB-4, моноцитарных колониестимулирующих факторов, гранулоцитарно-моноцитарно колониестимулирующих факторов приводит к пролиферации моноцитов, которые затем экспрессируют скавенджер-рецепторы и, захватывая модифицированные липопротеины низкой плотности, превращаются в макрофаги. Макрофаги, в свою очередь захватившие большое количество модифицированных липопротеинов низкой плотности, превращаются в пенистые клетки [7, 6]. При участии моноцитарных колониестимулирующих факторов также появляются макрофаги, не трансформирующиеся в пенистые клетки и в дальнейшем секретирующие провоспалительные цитокины (интерлейкин-1b и фактор некроза опухоли) [8].
Важную роль в атерогенезе в настоящее время приписывают белкам β-аррестинам. Известно, что β-аррестин-2 способствует патогенезу атеросклероза. У человека коронарные артерии с атеросклеротическими поражениями содержат в два раза больше мРНК β-аррестина, чем неизмененные коронарные артерии. Доказано также, что β-аррестин увеличивает пролиферацию и миграцию ГМК и, соответственно, гиперплазию неоинтимы. Кроме того, обнаружено, что β-аррестин-1 не оказывает данные эффекты и подавляет процессы пролиферации и миграции ГМК в естественных условиях. Таким образом, β-аррестин-1 и β-аррестин-2 являются антагонистами [19].
В качестве фактора, способствующего уменьшению пролиферативной активности ГМК, рассматривают ангиотензин (1–7) — недавно описанный компонент ренин-ангиотензиновой системы, образующийся из ангиотензина I и ангиотензина II с помощью ангиотензинпревращающего фермента-2 [20, 21]. Ангиотензин (1–7) ингибирует миграцию ГМК и их пролиферацию, а также способствует стабилизации бляшки за счет снижения уровня провоспалительных цитокинов и матриксных металлопротеиназ.
Таким образом, образование пенистых клеток макрофагального происхождения, как и образование пенистых клеток из ГМК, в совокупности с процессами пролиферации макрофагов [6] и ГМК под действием ряда факторов может представлять собой основную стадию образования утолщений интимы при развитии атеросклеротических поражений.
Помимо моноцитов/макрофагов и ГМК дисфункция эндотелиальных клеток является важным звеном в патогенезе атеросклероза. Ряд факторов может влиять на пролиферативную активность эндотелиальных клеток. Так, липопротеины высокой плотности оказывают антиатерогенное действие в том числе и благодаря их модулирующему влиянию на способность эндотелиальных клеток к пролиферации. Было установлено, что липопротеины высокой плотности способствуют пролиферации эндотелия и уменьшают апоптоз эндотелиальных клеток [22]. Показано большое значение эндотелиальных микроРНК (MIR), которые играют решающую роль в процессе развития сосудов, а также в эндотелиальном ответе на гемодинамический стресс и воспаление [23]. Выяснилось, что MIR-126-5p способствует пролиферации эндотелиальных клеток, регенерации эндотелия коронарных сосудов и препятствует образованию атеросклеротических поражений [21]. Кроме этих факторов упоминаемый выше ангиотензин (1–7) стимулирует пролиферацию эндотелиальных клеток-предшественников, которые способствуют регенерации поврежденного эндотелиального слоя.
Напротив, отрицательно влиять на эндотелий могут факторы комплемента, белки острой фазы и цитозольные рецепторы вирусных нуклеиновых кислот RIG-1, MDA-5. Процесс активации MDA-5 и RIG-1 индуцирует выработку интерферона-γ дендритными клетками и макрофагами и способствует снижению пролиферативной активности эндотелиальных клеток [24].
Клеточная пролиферация в различных типах атеросклеротических поражений
Анализ литературы позволил выявить, что пролиферативная активность зависит от типа атеросклеротического поражения.
В 1995 г. группа ученых во главе с M.D. Rekhter [25] изучала атеросклеротические бляшки, в результате были определены топографические закономерности распределения пролиферирующих клеток в зависимости от их типа. Были проанализированы образцы атеросклеротических бляшек, извлеченных из сонных и внутренних грудных артерий в ходе эндартерэктомии. Известно, что данные атеросклеротические бляшки являлись причиной стеноза, но ни одна из них не вызывала полную окклюзию сосуда. В качестве контроля использовали участок внутренней грудной артерии, полученный после коронарного шунтирования.
Из пролиферирующих клеток в интиме преобладали моноциты/макрофаги (46,0 % PCNA-позитивных клеток), ГМК было 19,7 %, эндотелиальных клеток микрососудов — 14,3 %, Т-клеток — 13,1 %.
В медии из пролиферирующих клеток преобладали ГМК (44,4 %), затем следовали эндотелиальные клетки микрососудов (20 %), моноциты/макрофаги (13 %), Т-клетки (14,3 %).
Участки локализации пенистых клеток характеризовались большим числом пролиферирующих макрофагов (66,5 %), тогда как неоваскуляризированные участки были PCNA-позитивными по эндотелиальным клеткам микрососудов (23,3 %), моноцитам/макрофагам (26,3 %), ГМК (14 %) и в меньшей степени по Т-клеткам (8,2 %).
Фиброзная покрышка бляшки, где степень пролиферации была минимальной, характеризовалась пролиферацией Т-клеток (34,1 %), моноцитов/макрофагов (14,5 %), ГМК (11 %). При этом пролиферации среди эндотелиальных клеток не наблюдалось, что, как отмечают авторы, может быть связано с особенностями проведения эндартерэктомии, а не с отсутствием способности эндотелиальных клеток к пролиферации [25].
Таким образом, эти авторы подтверждают данные А.Н. Восканьянц и В.А. Нагорнева [1] о пролиферации гематогенных клеток в атеросклеротической бляшке. Существует, однако, точка зрения, что при формировании атеросклеротических поражений пролиферируют в основном оседлые клетки сосудистой стенки, а не клетки гематогенного происхождения [26].
Роль клеточной пролиферации при дестабилизации атеросклеротической бляшки у человека
Если при атеросклерозе процесс клеточной пролиферации мало изучен, то при формировании нестабильной атеросклеротической бляшки у человека он совершенно неизвестен. А согласно современным данным, не атеросклеротическая бляшка вообще, а именно ее нестабильная форма служит причиной наиболее тяжелых и острых осложнений атеросклероза и ишемической болезни сердца и приводит к развитию острого коронарного синдрома [27, 28]. Именно поэтому с целью выяснения вопроса о роли клеточной пролиферации в прогрессировании атеросклероза и дестабилизации атеросклеротической бляшки у человека нами было предпринято исследование, которое позволило установить ряд новых фактов.
Иммуногистохимический анализ показал, что в нормальных участках интимы артерий у человека либо присутствуют единичные пролиферирующие клетки, либо они отсутствуют совсем. На начальных стадиях формирования атеросклеротических поражений в липидном пятне наблюдается высокая пролиферативная активность эндотелиальных клеток. При формировании нестабильной атеросклеротической бляшки в ее покрышке выявляются многочисленные мононуклеарные и гладкомышечные клетки, находящиеся в стадии активной пролиферации (рис. 1, a). Важно, что пролиферация мононуклеарных и гладкомышечных клеток происходит не только в поверхностных, но и в глубоких отделах нестабильной атеросклеротической бляшки. Интересно, что на далекозашедших стадиях деструкции покрышки нестабильной атеросклеротической бляшки в зонах ее выраженного повреждения и даже в районах отслоения и разрыва отмечается высокая клеточная пролиферативная активность (рис. 1, b).
В стабильных атеросклеротических поражениях высокая пролиферативная активность характерна прежде всего для ГМК. Так, в нижних отделах покрышки стабильной бляшки, на границе с атероматозным ядром, обнаружена выраженная пролиферация ГМК, что, видимо, связано с упрочнением покрышки и созданием мощного соединительнотканного барьера между кровью и очагом отложения липидов. Интересно, что одновременно многочисленные пролиферирующие ГМК выявлены на границе медии и адвентиции непосредственно под стабильной атеросклеротической бляшкой (рис. 1, c). В самой же покрышке стабильной атеросклеротической бляшки мононуклеарные клетки, находящиеся в стадии пролиферации, либо встречаются редко, либо отсутствуют совсем (рис. 1, d).

Рис. 1. Клеточная пролиферация в различных типах атеросклеротических поражений у человека (а, b, c, d — иммунопероксидазный метод на выявление PCNA-положительных клеток с помощью моноклональных антител. ×750): а — многочисленные пролиферирующие мононуклеарные (m) и гладкомышечные (d′) клетки в покрышке нестабильной атеросклеротической бляшки; b — в поврежденной покрышке нестабильной атеросклеротической бляшки видны многочисленные пролиферирующие лимфоциты (1) и макрофаги (2); c — пролиферирующие гладкомышечные клетки, располагающиеся на границе медии и адвентиции под стабильной атеросклеротической бляшкой; d — отсутствие пролиферации в плотной фиброзной покрышке стабильной атеросклеротической бляшки
Fig. 1. Cellular proliferation in various types of atherosclerotic lesions in human (a, b, c, d — an immunoperoxcidase method on PCNA identification-positive cells by means of monoclonal antibodies. ×750): a — numerous proliferating mononuclear (m) and smooth muscule (d′) cells in a cap of unstable atherosclerotic plaque; b — in the destructive cap of unstable atherosclerotic plaque numerous proliferating lymphocytes (1) and macrophages (2) are visible; c — the proliferating smooth muscule cells which are located on border of media and adventitia under a stable atherosclerotic plaque; d — lack of a proliferation in the dense fibrous cap of a stable atherosclerotic plaque
Заключение
Результаты исследования показывают, что на всех стадиях формирования нестабильной атеросклеротической бляшки, начиная с самой ранней, наблюдается выраженная клеточная пролиферация. Уже на стадии липидного пятна обнаружены многочисленные пролиферирующие эндотелиальные клетки, что связано с необходимостью замещения поврежденного эндотелия в начальный период формирования иммуновоспалительных реакций в сосудистой стенке. Обращает на себя внимание активная пролиферация мононуклеарных клеток — лимфоцитов и макрофагов — как в поверхностных, так и в глубоких отделах интимы и покрышки на всех стадиях формирования нестабильной атеросклеротической бляшки. Вероятно, именно пролиферирующие в стенке артерий клетки принимают участие в реакции иммунного воспаления, так как макрофаги гематогенного происхождения участвуют преимущественно в скавенджер-захвате модифицированных форм липопротеинов низкой плотности и трансформируются в пенистые клетки, составляющие основу бляшки. Незначительное содержание пролиферирующих мононуклеаров в стабильных атеросклеротических бляшках, по-видимому, обусловлено тем, что их плотная фиброзная покрышка не пропускает эти клетки в интиму. Выраженная пролиферация ГМК, наблюдающаяся под покрышкой стабильной бляшки на границе с атероматозным ядром, обусловлена созданием соединительнотканного барьера между кровью и ядром.
Список сокращений
ГМК — гладкомышечные клетки; ИФР-1 — инсулиноподобный фактор роста; MIR — микроРНК.
Читайте также: