
Поглощение воды корнем кратко
Обновлено: 05.07.2024
Поглощение воды и растворенных в ней питательных веществ является одной из наиболее важных и сложных функций корня.
Корни могут усваивать только жидкую воду, которая присутствует в почве в различных формах: гравитационную, капиллярную, пленочную и гигроскопическую, парообразную.
Вода удерживается в почве с различной силой. Водоудерживающая способность почвы зависит от наличия в ней коллоидных и осмотически активных веществ. Для того чтобы преодолеть водоудерживающие силы почвы, корни развивают в ней сосущую силу, которая должна превышать осмотическое давление почвенного раствора. Наиболее корнедоступной формой почвенной влаги является гравитационная и капиллярная вода. Труднодоступна для растений пленочная вода. Гигроскопическая вода удерживается почвой с силой 101 300 кПа (1000 атм), для растений она практически недоступна.
Различают пассивное и активное поглощение воды корнем. При пассивном поглощении силы, обеспечивающие поступление воды в ткани корня, формируются за счет дефицита воды в тканях растения и наличия ее в свободном состоянии в окружающей почвенной среде. В этом случае корни выполняют пассивную роль передачи воды из почвы в обезвоженные ткани растения. Механизм пассивного поглощения воды объясняется транспирацией надземных частей растений в основном листьями и хвоей. При этом капилляры сосудисто-волокнистых пучков ксилемы обеспечивают большую прочность на разрыв заключенных в них нитей воды. В результате транспирации в листьях и хвое создается дефицит диффузионного давления, что приводит к снижению давления воды в тканях ксилемы. Это снижение давления по законам единой гидростатической системы передается до самых тонких окончаний корней. Сила, поднимающая нити воды в ксилеме, бывает настолько велика, что в период наибольшей интенсивности транспирации, обычно приуроченной к летнему полдню, может наблюдаться уменьшение диаметра ствола до такой степени, что это свободно фиксируется более или менее точными измерительными приборами.
При относительно достаточном наличии воды в почве пассивное поглощение может осуществляться и через старые корни, причем в довольно больших количествах.
При активном поглощении воды силы, обеспечивающие этот процесс, формируются и развиваются в самом корне и проявляются в виде активной сосущей силы. Сосущая сила обеспечивает подачу воды в ткани корня не только при влажности почвы, превышающей влажность тканей растения, но даже тогда, когда степень увлажненности корня выше, чем относительная влажность почвы.
Наличие активного поглощения воды корнями подтверждается явлением корневого давления. Корневым давлением называется сила, которая поднимает воду вверх по сосудам. В результате корневого давления из живых пней или повреждений на стволе дерева может обильно выделяться пасока. Это явление широко используется при добыче сока березы, клена. Существенное влияние при этом имеют наличие и состояние воды в почве. При избытке увлажненности жизнедеятельность корней угнетается в результате недостатка кислорода, что вызывает снижение всасывающей активности корней. При слишком малом количестве почвенной влаги, сосущей силы корней может быть недостаточно для того, чтобы преодолеть силы сцепления, удерживающие воду около почвенных частиц. Сосущая сила корней с уменьшением насыщенности почвы водой от полевой влагоемкости до влажности увядания увеличивается от 0,1 до 15 атм (соответственно 10,1 и 1519,5 кПа).
Фактором, уменьшающим интенсивность поглощения воды корнями, является высокая концентрация солей в почвенном растворе. Сильная задержка в росте растений в результате недостатка влаги проявляется, когда осмотическое давление почвенного раствора достигает уровня 2—3 атм (до 303,9 кПа). При этом даже если влажность почвы соответствует полевой влагоемкости, всасывание воды корнями может почти полностью прекратиться. Особенно отрицательно влияет наличие токсичных для молодых корней солей хлоридов и сульфатов.
Непосредственное влияние на интенсивность поглощения воды корнями имеет температура почвы. Низкие температуры значительно ее снижают. При этом на корни древесных пород — выходцев из северных широт отрицательное влияние низких температур проявляется несколько слабее.
Высокие температуры почвы также непосредственно снижают интенсивность всасывающей поверхности корней. Отрицательное влияние высоких температур почвы обычно проявляется через иссушение почвы, а также обусловленное этим повышение концентраций солей в почвенном растворе.
Для большинства растений оптимальной температурой, которой соответствуют наивысшие показатели корневого давления, является температура в пределах 15—30 °С. Снижение температуры до 12°С значительно снижает корневое давление, а повышение более 30 °С вызывает его скачкообразное увеличение с постепенным последующим снижением. При снижении температуры корневой системы до 0°С поступление воды в растения снижается настолько, что даже если надземная часть растения при этом находится при температуре 20—22 °С, то оно завядает. У древесных пород в отдельные вегетационные периоды — это явление приводит к сбрасыванию осенью еще зеленых листьев, не поврежденных морозом.
Основным поставщиком воды в растительный организм являются тонкие сосущие корни, сосредоточенные в корневых мочках. В связи с этим все факторы, отрицательно влияющие на жизнедеятельность сосущих корней, вызывают нарушение влагоснабжения растений. Накопление определенного запаса воды в толстых корнях, стволе и ветвях деревьев в неблагоприятных ситуациях может служить своего рода биологическим буфером, позволяющим смягчить на некоторый период отрицательное воздействие чрезмерного дефицита влаги в почве или временную потерю части всасывающих корней.
В силу медленной миграции воды в почве отдельные участки в зоне расположения корневых мочек оказываются иссушенными в большей степени по сравнению с зонами, менее насыщенными корнями. В таких случаях продвижение осевых ростовых корней обеспечивает освоение корневой системой более увлажненных зон почвы.
При сильном иссушении почвы сосущая сила корней несколько увеличивается в результате повышения осмотического давления в клетках корня за счет накопления сахаров и аккумуляции солей.
В корне существует градиент водного потенциала — от более высокого в клетках, образующих корневые волоски, к более низкому в клетках, примыкающих к ксилеме. Этот градиент поддерживается двумя способами:
- за счет движения воды вверх по ксилеме, при котором, как мы уже говорили, в ксилеме создается натяжение (отрицательное давление) и тем самым понижается водный потенциал ксилемного сока;
- за счет того, что осмотический потенциал ксилемного сока более низкий (более отрицательный) по сравнению с осмотическим потенциалом разбавленного почвенного раствора.
Вода движется через корень по тем же путям, что и в листьях, а именно по апопласту, симпласту и через вакуоли. По мере того как вода поднимается вверх по корневой ксилеме, ее замещает вода из окружающих паренхимных клеток. В результате водный потенциал этой клетки снижается и в нее устремляется вода из соседней клетки 2 благодаря осмосу или просто по симпласту. Тогда в свою очередь снижается и водный потенциал клетки 2, в нее начинает поступать вода из клетки 3 и так далее через весь корень до самого эпидермиса, образующего волоски. Водный потенциал почвенного раствора выше, чем в клетках эпидермиса и в корневых волосках. Следовательно, вода проникает в корень извне путем осмоса.
Поглощение воды корнями. Апопластный транспорт в корне.
На рисунке показано первичное строение корня типичного двудольного растения.
Основная масса воды поглощается более молодыми частями корня в зоне корневых волосков. По мере роста корня в почве на нем постоянно образуются новые корневые волоски, а старые отмирают. Новые корневые волоски возникают на некотором расстоянии позади зоны растяжения. Эти волоски представляют собой трубчатые выросты эпидермальных клеток (рис. 13.17), существенно увеличивающие поверхность, способную поглощать воду и минеральные соли. Они вступают в тесный контакт с частицами почвы.
На рисунке схематично показаны пути движения воды по корню. В корне существует градиент водного потенциала — от более высокого в клетках, образующих корневые волоски, к более низкому в клетках, примыкающих к ксилеме. Этот градиент поддерживается двумя способами:
1) за счет движения воды вверх по ксилеме, при котором, как мы уже говорили, в ксилеме создается натяжение (отрицательное давление) и тем самым понижается водный потенциал ксилемного сока;
2) за счет того, что осмотический потенциал ксилемного сока более низкий (более отрицательный) по сравнению с осмотическим потенциалом разбавленного почвенного раствора.
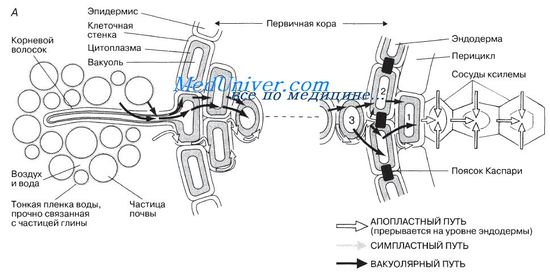
Вода движется через корень по тем же путям, что и в листьях, а именно по апопласту, симпласту и через вакуоли.
По мере того как вода поднимается вверх по корневой ксилеме, ее замещает вода из окружающих паренхимных клеток, например из клетки 1 на рисунке. В результате водный потенциал этой клетки снижается и в нее устремляется вода из соседней клетки 2 благодаря осмосу или просто по симпласту, как описано в разд. 13.3.2 для мезофилла листа. Тогда в свою очередь снижается и водный потенциал клетки 2, в нее начинает поступать вода из клетки 3 и так далее через весь корень до самого эпидермиса, образующего волоски.
Водный потенциал почвенного раствора выше, чем в клетках эпидермиса и в корневых волосках. Следовательно, вода проникает в корень извне путем осмоса.
Апопластный транспорт в корне
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Корень — осевой подземный орган растения, обладающий неограниченным концевым ростом.

Главный корень развивается из зародышевого корешка семени и играет в растении роль центральной оси подземной части.
Придаточные корни растут от побега.
Боковые корни образуются на главном и придаточных корнях.

Рис. 1. Виды корней
Вся совокупность корней растения называется корневой системой.
типы корневых систем
В зависимости от развития тех или иных видов корней выделяют два типа корневых систем (рис. 2). Стержневая корневая система состоит из хорошо развитого главного корня и отходящих от него более мелких боковых корней, которые в свою очередь делятся на боковые корни второго, третьего и т. д. порядков.
Такая корневая система характерна для двудольных растений и хорошо просматривается только у молодых растений, выращенных из семян. У старых многолетних растений главный корень со временем замедляет рост, а боковые корни догоняют его или даже перерастают.
Мочковатая корневая система состоит из многочисленных придаточных и боковых корней. Главный корень не развивается или развивается слабо.
Мочковатая корневая система характерна для однодольных растений.

Рис. 2. Типы корневых систем
внутреннее строение корня
В строении корня различают несколько зон, каждая из которых имеет определенное строение и выполняет определенные функции (рис. 3).
Зона деления состоит из мелких постоянно делящихся клеток верхушечной меристемы. Это зона находится на кончиках всех корней растения. Благодаря верхушечной меристеме осуществляется рост корня в длину.
Корневой чехлик — несколько слоёв плотно сросшихся клеток с утолщенными стенками.
Функция корневого чехлика:
- механическая защита зоны деления;
- выделение слизистых веществ для более легкого проникновения в почву.
Клетки снаружи корневого чехлика постоянно разрушаются, а с внутренней стороны он нарастает благодаря клеткам меристемы.
Пикировка корня — удаление кончика главного корня — производится с целью прекращения роста главного корня и усиления роста боковых корней: общая площадь корневого питания увеличивается.

Рис. 3. Зоны корня
Зона растяжения (роста). В ней клетки растут, вытягиваясь в длину, благодаря чему и происходит удлинение корня.
В этой же зоне начинается дифференцировка клеток. Поверхностные клетки превращаются в клетки эпидермы. В центре формируются клетки проводящих тканей.
Зона всасывания. Зона всесывания снаружи покрыта тонкой покровной тканьюэпиблемой (или ризодермой). В этой зоне клеткиэпиблемы образуют выросты —корневые волоски. Корневые волоски представляют собой длинные тонкие нитевидные клеточные выросты, в которые перемещается ядро клетки. По мере роста корня они разрушаются, эпидерма замещается пробкой и зона всасывания замещается зоной проведения.
Функция корневых волосков: поглощение из почвы воды и минеральных веществ.
Зона проведения продолжается до наземных частей растения. В ней находятся сосуды ксилемы, по которым от корня поднимается вода с минеральными веществами, и ситовидные трубки флоэмы, по которым в корень поступают органические вещества из листьев.
Корни подавляющего большинства растений выполняют шесть основных функций:
- Корни удерживают растение в определённом положении. Эта функция очевидна для наземных растений, особенно значима она для крупных деревьев с большой массой ветвей и листьев. У многих водных растений закрепление на дне позволяет выгодно распределить в пространстве листья. У плавающих растений, например у ряски, корни не позволяют растению переворачиваться.
- Корни осуществляют почвенное питание растения, поглощая из почвы воду с растворёнными в ней минеральными веществами, и проведение веществ к побегу (рис. 1).
- У некоторых растений в главном корне осуществляется хранение запасных питательных веществ, таких как крахмал и другие углеводы.
- В корнях происходит образование определённых веществ, нужных организму растения. Так, в корнях осуществляется восстановление нитратов до нитритов, синтез некоторых аминокислот и алкалоидов.
- Корни могут осуществлять симбиоз с грибами и микроорганизмами, обитающими в почве (микориза, клубеньки представителей семейства Бобовые).
- С помощью корней может осуществляться вегетативное размножение (например, корневыми отпрысками). Корневыми отпрысками размножаются, например, одуванчик, слива, малина, сирень.

Поглощение воды и минеральных веществ корнем
Эта функция возникла у растений в связи с выходом на сушу.
Поглощение воды и минеральных веществ растением происходит независимо друг от друга, так как эти процессы основаны на различных механизмах действия. Вода проходит в клетки корня пассивно, а минеральные вещества поступают в клетки корня в основном в результате активного транспорта, идущего с затратами энергии.

Рис. 1. Горизонтальный транспорт воды:
1 — корневой волосок; 2 — апопластный путь; 3 — симпластный путь; 4 — эпиблема (ризодерма) 5 — эндодерма; 6 — перицикл; 7 — сосуды ксилемы; 8 — первичная кора; 9 — плазмодесмы; 10 — пояски Каспари.
Вода поступает в растение в основном по закону осмоса. Корневые волоски имеют огромную вакуоль с концентрированным клеточным соком, обладающую большим осмотическим потенциалом, который обеспечивает поступление воды из почвенного раствора в корневой волосок.
ГОРИЗОНТАЛЬНЫЙ ТРАНСПОРТ ВЕЩЕСТВ
Вода попадает в тело растения через ризодерму, поверхность которой сильно увеличена благодаря наличию корневых волосков.
В этой зоне в проводящем цилиндре корня формируется проводящая система корня — сосуды ксилемы, необходимая для обеспечения восходящего тока воды и минеральных веществ.
Вода с минеральными солями поглощается корневыми волосками. Эндодерма перекачивает эти вещества в проводящий цилиндр, создавая корневое давление и не позволяя воде выходить назад. Вода с солями поступает в сосуды проводящего цилиндра и поднимается транспирационным током по стеблю к листьям.
Горизонтальный транспорт воды и минеральных веществ происходит по трём путям (рис. 1):
- апопластный путь (основной для воды и минеральных веществ) — путь через межклеточные пространства и клеточные стенки;
- симпластный путь (для транспорта органических и минеральных веществ) — путь через систему протопластов (внутреннего содержимого) клеток, соединённых посредством плазмодесм (цитоплазматических мостиков);
- вакуолярный путь (только транспорт воды; в корне минимален) — путь из вакуоли в вакуоль через другие компоненты смежных клеток (плазматические мембраны, цитоплазма и тонопласт вакуолей).
В корне вода передвигается по межклетникам до эндодермы (апопластный путь). Здесь её дальнейшему продвижению мешают водонепроницаемые клеточные стенки, пропитанные суберином (пояски Каспари). Поэтому вода попадает в стелу по симпласту через пропускные клетки (вода проходит через плазматическую мембрану под контролем цитоплазмы пропускных клеток эндодермы). Благодаря этому происходит регуляция движения воды и минеральных веществ из почвы в ксилему. В стеле вода уже не встречает сопротивления и поступает в проводящие элементы ксилемы.
ВЕРТИКАЛЬНЫЙ ТРАНСПОРТ ВЕЩЕСТВ
Корни осуществляют проведение воды и минеральных веществ к наземным органам растения.
Вертикальное перемещение воды происходит по мёртвым клеткам ксилемы, которые не способны толкать воду к листьям. Это движение поддерживается транспирационной функцией листьев.
Корень представляет собой нижний концевой двигатель, подающий воду в сосуды стебля под давлением.
Корневое давление — сила, с которой корень нагнетает воду в стебель.
Корень активно перекачивает минеральные и органические вещества в сосуды ксилемы; в результате возникает повышенное осмотическое давление в сосудах корня относительно с давлением почвенного раствора. Величина корневого давления может достигать 3 атм. Доказательством наличия корневого давления служит, например,гуттация (выделение капелек воды листьями).
Верхний концевой двигатель, обеспечивающий вертикальный транспорт воды, возникает в результате транспирации (испарения воды с поверхности листьев).
При непрерывном испарении воды создаётся возможность для нового притока воды к листьям. Сосущая сила листьев у деревьев может достигать 20 атм.
В сосудах ксилемы вода движется в виде непрерывных водяных нитей. Молекулы воды движутся друг за другом за счет когезии (сцепления друг с другом) и адгезии (прилипания к стенкам сосудов).
Таким образом, поднятие воды по растению осуществляется благодаря верхнему и нижнему двигателям водного тока и силам сцепления молекул воды в сосудах. Основной движущей силой является транспирация.
ОСМОС И ТУРГОР
В живых клетках корня происходит первый отбор веществ, допускаемых внутрь растения. Участие живых клеток в принятии веществ обусловливает избирательную способность растения, благодаря которой различные вещества поглощаются в разных количествах. Так как поступление в сильной степени зависит от потребления, растение принимает на различных стадиях развития то одни соли, то другие. Чем сильнее развита корневая система, тем активнее идёт поглощение воды и солей.
Часто возникают ситуации, когда корни растений выполняют некоторые дополнительные функции или одна из основных функций требует большего развития. В таких случаях образуются видоизменения корней
Вода является основной составной частью растительных организмов. Её содержание доходит до 90 % от массы организмов, и она участвует прямо или косвенно во всех жизненных проявлениях. Вода- это та среда, в которой протекает все процессы обмена веществ. Она составляет основную часть цитоплазмы, поддерживает ее структуру, устойчивость входящих в состав цитоплазмы коллоидов, обеспечивает определенную конфирмацию молекул белка. Высокое содержание воды придает содержимому клетки (цитоплазме) подвижный характер. Вода - непосредственный участник многих химических реакций. Все реакции гидролиза, многочисленные окислительно-восстановительные реакции идут с участием воды.
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой (тургор) обеспечивает прочность тканей, сохранение структуры травянистых растений, определенную ориентировку организмов растений в пространстве. Рост клеток в фазе растяжения идет главным образом за счет накопления воды в вакуоли.
Таким образом, вода обеспечивает протекание процессов обмена, , связь организма со средой. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой.
Растение может испытывать недостаток влаги не только в тех случаях, когда ее мало в почве. Сильная кислотность почвы и высокая концентрация легкорастворимых солей в почве могут ограничивать всасывающую силу корней при достаточном содержании воды. Такое состояние почвы в отличие от физической сухости называютфизиологической сухостью.
Морфоанотомические основы поглощения и движения воды.
Корневая система как орган поглощения воды.
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Меристемы- образовательные ткани- расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля).
Рост корней отличается большой скоростью.Только благодаря такому интенсивному росту корневые системы растения могут использовать скудно рассеянную в почве воду. Важное значение имеет явление гидротропизма, при котором рост корневой системы как бы идет из более иссушенных слоев почвы к более влажным. В зависимости от типа растений распределение корневой системы в почве различно. У некоторых растений корневая система проникает на большую глубину, у других главным образом распространяется в ширину.
С физиологической точки зрения корневая система не однородна. Довольно не вся поверхность корня участвует в поглощении воды. В каждом корне различают несколько зон. Правда, не всегда все зоны выражены одинаково четко.
Окончание корня с наружи защищено корневым чехликом, напоминающим округлый колпачок, таящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика суживаются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усилено делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона-зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки, затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а так же расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.
Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры. Кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системных крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток- эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином .При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения, как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня. При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление волосков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является основной зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет 5-10 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче.

Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными организмами, в связи с тем что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем.
Большое значение для формирования корневых систем играет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватывают атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков.
Важное значение для развития корневых систем имеет аэрация. Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания. Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений - их усиленному ветвлению.
Введение
Вода является основной составной частью растительных организмов. Её содержание доходит до 90 % от массы организмов, и она участвует прямо или косвенно во всех жизненных проявлениях. Вода- это та среда, в которой протекает все процессы обмена веществ. Она составляет основную часть цитоплазмы, поддерживает ее структуру, устойчивость входящих в состав цитоплазмы коллоидов, обеспечивает определенную конфирмацию молекул белка. Высокое содержание воды придает содержимому клетки (цитоплазме) подвижный характер. Вода - непосредственный участник многих химических реакций. Все реакции гидролиза, многочисленные окислительно-восстановительные реакции идут с участием воды.
Водный ток обеспечивает связь между отдельными органами растений. Питательные вещества передвигаются по растению в растворенном виде. Насыщенность водой (тургор) обеспечивает прочность тканей, сохранение структуры травянистых растений, определенную ориентировку организмов растений в пространстве. Рост клеток в фазе растяжения идет главным образом за счет накопления воды в вакуоли.
Таким образом, вода обеспечивает протекание процессов обмена, , связь организма со средой. Для нормальной жизнедеятельности клетка должна быть насыщенна водой.
Основным источником влаги является вода, находящаяся в почве, и основным органом поглощения воды является корневая система. Роль этого органа прежде всего заключается в том, что благодаря огромной поверхности обеспечивается поступление воды в растения из возможно большого объема почвы. Сформировавшаяся корневая система представляет собой сложный орган с хорошо дифференцированной внешней и внутренней структурой.
Растение может испытывать недостаток влаги не только в тех случаях, когда ее мало в почве. Сильная кислотность почвы и высокая концентрация легкорастворимых солей в почве могут ограничивать всасывающую силу корней при достаточном содержании воды. Такое состояние почвы в отличие от физической сухости называютфизиологической сухостью.
Морфоанотомические основы поглощения и движения воды.
Корневая система как орган поглощения воды.
Рост корня, его ветвление продолжается в течение всей жизни растительного организма, то есть практически он не ограничен. Меристемы- образовательные ткани- расположены на верхушке каждого корня. Доля меристематических клеток сравнительно велика (10% по массе против 1% у стебля).
Рост корней отличается большой скоростью.Только благодаря такому интенсивному росту корневые системы растения могут использовать скудно рассеянную в почве воду. Важное значение имеет явление гидротропизма, при котором рост корневой системы как бы идет из более иссушенных слоев почвы к более влажным. В зависимости от типа растений распределение корневой системы в почве различно. У некоторых растений корневая система проникает на большую глубину, у других главным образом распространяется в ширину.
С физиологической точки зрения корневая система не однородна. Довольно не вся поверхность корня участвует в поглощении воды. В каждом корне различают несколько зон. Правда, не всегда все зоны выражены одинаково четко.
Окончание корня с наружи защищено корневым чехликом, напоминающим округлый колпачок, таящий из живых тонкостенных продолговатых клеток. Корневой чехлик служит защитой для точки роста. Клетки корневого чехлика суживаются, что уменьшает трение и способствует проникновению корня в глубь почвы. Под корневым чехликом расположена меристематическая зона. Меристема состоит из многочисленных мелких, усилено делящихся, плотно упакованных клеток, почти целиком заполненных протоплазмой. Следующая зона-зона растяжения. Здесь клетки увеличиваются в объеме (растягиваются). Одновременно в этой зоне появляются дифференцированные ситовидные трубки, затем следует зона корневых волосков. При дальнейшем увеличении возраста клеток, а так же расстояния от кончика корня корневые волоски исчезают, начинается кутинизация и опробковение клеточных оболочек. Поглощение воды происходит главным образом клетками зоны растяжения и зоны корневых волосков.
Поверхность корня в зоне корневых волосков покрыта ризодермой. Это однослойная ткань с двумя видами клеток, формирующими и не формирующими корневые волоски. В настоящее время показано, что клетки, формирующие корневые волоски, отличаются особым типом обмена веществ. У большинства растений клетки ризодермы обладают тонкими стенками. Вслед за ризодермой до перицикла идут клетки коры. Кора состоит из нескольких слоев паренхимных клеток. Важной особенностью коры является развитие системных крупных межклетников. На границе коры и центрального цилиндра развивается один слой плотно прилегающих друг к другу клеток- эндодерма, для которой характерно наличие поясков Каспари. Цитоплазма в клетках эндодермы плотно прилегает к клеточным оболочкам. По мере старения вся внутренняя поверхность клеток эндодермы, за исключением пропускных клеток, покрывается суберином .При дальнейшем старении сверху могут накладываться еще слои. По-видимому, именно клетки эндодермы служат основным физиологическим барьером для передвижения, как воды, так и питательных веществ. В центральном цилиндре расположены проводящие ткани корня. При рассмотрении структуры корня в продольном направлении важно отметить, что начало роста корневых волосков, появление волосков Каспари в стенках эндодермы и дифференциация сосудов ксилемы происходят на одном и том же расстоянии от апикальной меристемы. Именно эта зона является основной зоной снабжения растений питательными веществами. Обычно поглощающая зона составляет 5-10 см в длину. Величина ее зависит от скорости роста корня в целом. Чем медленнее растет корень, тем зона поглощения короче.
Надо отметить, что в целом корневые системы значительно менее разнообразны по сравнению с надземными организмами, в связи с тем что среда их обитания более однородна. Это не исключает того, что корневые системы изменяются под влиянием тех или иных условий. Хорошо показано влияние температуры на формирование корневых систем. Как правило, оптимальная температура для роста корневых систем несколько ниже по сравнению с ростом надземных органов того же растения. Все же сильное понижение температуры заметно тормозит рост корней и способствует образованию толстых, мясистых, мало ветвящихся корневых систем.
Большое значение для формирования корневых систем играет влажность почвы. Распределение корней по горизонтам почвы часто определяется распределением воды в почве. Обычно в первый период жизни растительного организма корневая система растет чрезвычайно интенсивно и, как следствие, скорее достигает более влажных слоев почвы. Некоторые растения развивают поверхностную корневую систему. Располагаясь близко к поверхности, сильно ветвящиеся корни перехватывают атмосферные осадки. В засушливых районах часто глубоко и мелко укореняющиеся виды растений растут рядом. Первые обеспечивают себя влагой за счет глубоких слоев почвы, вторые за счет усвоения выпадающих осадков.
Важное значение для развития корневых систем имеет аэрация. Именно недостаток кислорода является причиной плохого развития корневых систем на заболоченных почвах. Растения, приспособленные к росту на плохо аэрируемых почвах, имеют в корнях систему межклетников, которые вместе с межклетниками в стеблях и листьях составляют единую вентиляционную систему.
Большое значение имеют условия питания. Показано, что внесение фосфорных удобрений способствует углублению корневых систем, а внесение азотных удобрений - их усиленному ветвлению.
Поглощение корнями воды и минеральных веществ — наиболее важная их функция, неразрывно связана со всем комплексом процесса обмена веществ растительного организма.
Вода, поглощенная корневыми волосками, проделывает сложный путь по растению. К сожалению, факторы, обеспечивающие как процесс поглощения воды корневыми волосками, так и ее перемещение, еще изучены недостаточно.

Механизм поглощения воды
Имеется много различных теорий, объясняющих эти процессы. Согласно гидростатическим законам величина тургорного давления во всех частях клетки одинакова, поэтому всасывающая сила больше в той части, где больше осмотическое давление/
S = P — T, где S — всасывающая сила; P — осмотическое давление; T — тургорное давление.
В механизме поглощения питательных веществ корнем различают два качественно различных процесса: метаболический (активный) и неметаболический (пассивный).
Метаболическое поглощение питательных веществ связывается, в первую очередь, с дыханием клеток. Дыхание определяет существование активной поверхности цитоплазмы, скорость ее обновления, насыщенность акцепторами минеральных соединений, возможность взаимодействия с ними.
Неметаболический механизм поглощения веществ связывают с проникновением их в клетку по градиенту концентрации. Этот процесс не зависит от обмена веществ живой клетки. Основной движущей силой его признают диффузию и осмос.
Через корневые волоски вода поступает в клетки основной ткани корня. Далее через пропускные клетки — в сосуды центрального цилиндра, а затем в надземную часть растения по сосудам проводящей зоны. Сосуды корня в надземной части растения становятся сосудами стебля. Место перехода корня в стебель несколько утолщено и называется корневой шейкой.
Читайте также: