
Осмотические явления в клетке и их значение в жизни растения кратко
Обновлено: 05.07.2024
Под осмосом понимается диффузия воды через полупроницаемые мембраны. Полупроницаемость мембран - это их свойство пропускать воду и не пропускать растворенные вещества. Явление осмоса было открыто в 1828 г. французским ботаником Дютроше. Позже Пфеффером были сконструированы физические приборы - осмометры (осмотические ячейки).
Осмометр представляет собой систему с тремя элементами: полупроницаемая мембрана, раствор определенной концентрации и вода. В этой системе энергия молекул чистой воды (водный потенциал) выше, чем воды в растворе. Поэтому вода будет проходить по градиенту водного потенциала из наружного сосуда во внутренний сосуд и подниматься по трубке, соединенной с внутренним сосудом. При этом будет повышаться гидростатическое давление, под которым находится раствор в осмометре. При некоторой высоте столба раствора в трубке скорости диффузии воды из наружного сосуда во внутренний и из внутреннего в наружный сравняются, и подъем жидкости в трубке прекратится. Давление, которое отвечает такому равновесию, служит количественной характеристикой процесса осмоса. Оно называется осмотическим давлением. Следовательно, осмотическое давление равно тому давлению, которое нужно приложить к раствору, чтобы привести его в равновесие с чистым растворителем, отделенным от раствора полупроницаемой мембраной.
Живая растительная клетка обладает свойствами только что разобранной осмотической системы. Поскольку молекулы воды проходят через клеточные мембраны намного быстрее, чем растворенные вещества, можно с известной долей погрешности говорить о полупроницаемости мембран. Роль полупроницаемой мембраны (хоть и не идеальной) выполняют, прежде всего, плазмалемма и тонопласт, раствора клеточный сок вакуоли клетки.
Передвижение воды через полупроницаемые мембраны подчиняется следующим законам:
1. Осмотическое давление прямо пропорционально концентрации раствора - числу частиц в единице объема независимо от размеров и природы частиц (молекул или ионов).
2. Всякое вещество, концентрация которого равна 1 моль/л, развивает осмотическое давление, равное 22,4 атм (для неэлектролита) при О°С.
3. Осмотическое давление пропорционально абсолютной температуре Т = 273 + t °С.
4. Осмотическое давление раствора, содержащего несколько веществ, равно сумме осмотических давлений, вызываемых каждым из них.
Осмотическое давление подчиняется в общем виде закону Вант-Гоффа и выражается уравнением Р = CRTi, где Р - осмотическое давление; С - молярная концентрация раствора; R - газовая постоянная, равная 0,0821 л · атм / град. моль; Т - абсолютная температура; i - изотонический коэффициент, показывающий отношение числа частиц (молекул и ионов) в растворе к исходному числу молекул растворенного вещества. Для растворов неэлектролитов, например для сахара, изотонический коэффициент равен 1, для электролитов - больше 1.
Если клетку поместить в воду, то она, как и осмометр, будет поглощать ее и тем сильнее, чем выше концентрация клеточного сока. Но до каких пор клетка будет поглощать воду? При всасывании клеткой воды объем клеточного сока увеличивается, и цитоплазма начинает давить на оболочку клетки, растягивая ее. Оболочка, обладая значительной прочностью и вместе с тем эластичностью, в свою очередь оказывает сопротивление всякой силе, стремящейся ее растянуть, и тем большее, чем более она растянута. После прекращения действия растягивающей силы стенка снова возвращается к своим прежним размерам. Напряженное состояние клеточной стенки, вызванное давлением клеточного содержимого, носит название тургора, а противодавление оболочки на протопласт - тургорного давления.
По мере поступления воды в клетку эндосмос постепенно уменьшается в силу противодавления клеточной стенки. В какой-то момент внутренние и внешние силы диффузии становятся равными. Это состояние называется полным насыщением клетки водой. В данном случае клетка находится в равновесии с водой, осмотическое давление клеточного сока уравновешено тургорным давлением: Р = Т. Иначе можно это записать так: Р - Т = 0. В этом состоянии вода не входит в клетку и не выходит из нее, как бы высоко ни было ее осмотическое давление Р. Примером этого могут служить клетки водорослей и водных высших растений. Их клетки всегда находятся в состоянии почти полного насыщения.
Клетки наземных растений, наоборот, почти никогда не бывают полностью насыщенными водой. Благодаря потере воды листьями в процессе транспирации они обычно не достигают состояния полного тургора. Часть осмотического давления Р остается неуравновешенной тургорным давлением Т, и эта часть осмотического давления способна при погружении такой клетки в воду вызывать приток воды в клетку и некоторое увеличение размеров последней. В данном случае Р будет больше Т: Р = T+S или S = Р - Т. Эта дополнительная часть осмотического давления получила название сосущей силы S . Название не совсем удачное, ибо это не сила, а давление, как и Р и Т, разность между которыми она представляет. Следует подчеркнуть, что именно сосущая сила клетки, а не осмотическое давление клеточного сока определяет поглощение воды клеткой.
По отношению к концентрации клеточного сока внешние растворы могут быть: гипертоническими, осмотическое давление которых больше осмотического давления клеточного сока; изотоническими, осмотическое давление которых равно осмотическому давлению клеточного сока; гипотоническими, у которых это давление меньше, чем давление клеточного сока.
При помещении живой растительной клетки не в воду, а в гипертонический раствор тургор будет стремиться к нулю, а сосущая сила - к величине полного осмотического давления S = Р. В данном случае будут происходить следующие явления: раствор оттянет воду из клетки, клеточная стенка сократится, сократится и цитоплазма клетки, тургор исчезнет. Раствор начнет входить в пространство между стенкой и цитоплазмой, т. е. цитоплазма будет отходить от клеточной стенки. Это отставание цитоплазмы от оболочки клетки при погружении последней в гипертонический раствор называется плазмолизом. Плазмолиз свидетельствует о том,что имеются различия в проницаемости клеточной стенки и цитоплазмы, и, что самое главное, показывает, что цитоплазма жива.
Если плазмолизированную клетку снова поместить в воду, то происходит обратная картина: клетка всасывает воду, объем протопласта увеличивается; цитоплазма начинает плотно примыкать к стенке клетки. Переход плазмолизированной клетки в нормальное состояние носит название деплазмолиза.
Различают уголковый, вогнутый (судорожный) и выпуклый виды плазмолиза. При уголковом плазмолизе цитоплазма отстает только по уголкам клетки. При вогнутом плазмолизе цитоплазма становится угловатой, сохраняет в виде выступов те участки, которые более плотно связаны с клеточной стенкой. Длительный вогнутый плазмолиз можно получить, предварительно обработав клетки солями двухвалентных металлов, например кальция. Выпуклый плазмолиз быстро проявляется при низкой вязкости цитоплазмы, например, при воздействии одновалентных металлов. В этом случае разбухшая цитоплазма в виде колпачка прикрывает сократившийся тонопласт.
В природной обстановке, например при пересадке растений на засоленную почву, осмотическое давление почвенного раствора может быть выше, чем осмотическое давление клеточного сока. В этом случае не только будет поглощаться такими клетками, но, наоборот, будет выходить из клеток, вызывая обезвоживание последних, а затем и отмирание растения.
Если потеря тургора происходит не в растворе, как при плазмолизе, а от высыхания в воздухе в жаркие дни, то наблюдается явление циторриза.Цитоплазма при этом от клеточной стенки не отстает, а остается как бы припаянной к ней. Стенки клеток, выпячиваясь внутрь, деформируются, стремятся разорвать цитоплазму с определенной силой (силой циторриза). В данном случае тургор - величина отрицательная, а сосущая сила равна сумме осмотического и тургорного давлений: S = Р - (-Т) = Р + Т. Сила циторриза может достигать величины порядка 12-15 атм. Зависимость всех трех рассматриваемых величин от степени насыщения клетки водой показана на схеме.
Внешние условия существенно влияют на показатели осмотического давления и сосущей силы. Они очень низкие у водных растений (за исключением растений морей и других засоленных водоемов) и растений болот и высокие - у растений засушливых мест. У морских водорослей осмотическое давление достигает 25 - 30 атм. Клетки растений, произрастающих на засоленных почвах, развивают давление, измеряемое 100-150 атм. Растения открытых местообитаний отличаются более высокими величинами осмотического давления и сосущей силы, чем растения затененных мест.
Камбиальные клетки стволов древесных растений имеют осмотическое давление 20 - 30 атм. Паренхимные клетки содержат клеточный сок в концентрациях, соответствующих осмотическому давлению 4 - 5 атм. Сосущая сила клеток хвои ели в таежной зоне достигает 15 - 35 атм, а поглощающих корней колеблется от 6 до 8 атм (у сосны - от 6 до 11 атм).
Для определения осмотического давления используют плазмолитический и криоскопический методы. Плазмолитический метод основан на том, что в растворах плазмолитиков (веществ, вызывающих плазмолиз) разной концентрации начало уголкового плазмолиза клеток будет наблюдаться в том из них, осмотическое давление которого почти соответствует осмотическому давлению клеточного сока (изотонический раствор). Криоскопический метод основан на определении температуры замерзания отжатого клеточного сока.
Сосущую силу можно определить, основываясь: на измерении размеров ткани, помещенной в растворы различных концентраций (метод Уршпрунга); на измерении концентрации растворов, в которые помещены кусочки ткани с помощью рефрактометра (метод Н.А. Максимова и Н.с. Петинова) или по методу струек В.С. Шардакова. Размеры клеток и концентрация внешнего раствора не изменятся при помещении в него ткани в том случае, когда сосущие силы раствора и клеток совпадают.
Осмотические свойства клеток не только определяют поглощение воды растением, но и лежат в основе многих физиологических процессов, определяют различные виды движений у растений, в частности движения устьиц. От осмотических явлений в значительной мере зависит рост клеток. Упругое состояние тканей и органов растения (их тургесцентность) влияет на внешний вид и форму растения. Большая роль принадлежит осмотическим явлениям в передвижении воды и растворенных веществ, устойчивости растений к неблагоприятным внешним воздействиям, регулировании такого сложного процесса, как фотосинтез.
Жизнедеятельность клетки характеризуется непрерывно протекающими в ней процессами обмена веществ, причем цитоплазма избирательно реагирует на воздействие разных факторов внешней среды. В поглощении и выделении веществ большую роль играют процессыдиффузии и осмоса. Осмотическими называют явления, происходящие в системе, состоящей из двух растворов, разделенных полупроницаемой мембраной.
Содержание
22. Осмотические явления в клетке и их значения в жизни растений.
23. Поступление воды в растения. Влияние аэрации, температуры почвы и других внешних факторов на деятельность корневой системы.
31. Источники углерода для растений.
32. Роль аденозинфосфатов и других макроэргических соединений
40.Зависимость дыхания от условий среды.
41 Структура и функции белков протоплазмы.
42.Зависимость фотосинтеза от внешних условий.
Работа содержит 1 файл
ТИТУЛ ЛИСТ.docx
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА
ФГОУ ВПО «ДАЛЬНЕВОСТОЧНЫЙ ГОСУДАРСТВЕННЫЙ
КОНТРОЛЬНАЯ РАБОТА №1
Выполнил: студент 3 курса
Проверила: Куркова И.В.
22. Осмотические явления в клетке и их значения в жизни растений.
23. Поступление воды в растения. Влияние аэрации, температуры почвы и других внешних факторов на деятельность корневой системы.
31. Источники углерода для растений.
32. Роль аденозинфосфатов и других макроэргических соединений
40.Зависимость дыхания от условий среды.
41 Структура и функции белков протоплазмы.
42.Зависимость фотосинтеза от внешних условий.
22. Осмотические явления в клетке и их значения в жизни растений.
Жизнедеятельность клетки характеризуется непрерывно протекающими в ней процессами обмена веществ, причем цитоплазма избирательно реагирует на воздействие разных факторов внешней среды. В поглощении и выделении веществ большую роль играют процессыдиффузии и осмоса. Осм отическими называют явления, происходящие в системе, состоящей из двух растворов, разделенных полупроницаемой мембраной. В растительной клетке роль полупроницаемых пленок выполняют пограничные слои цитоплазмы: плазмалемма и тонопласт.
Плазмалемма - наружная мембрана цитоплазмы, прилегающая к клеточной оболочке. Тонопласт - внутренняя мембрана цитоплазмы, окружающая вакуоль. Вакуоли представляют собой полости в цитоплазме, заполненные клеточным соком - водным раствором углеводов, органических кислот, солей, белков с низким молекулярным весом, пигментов.
Концентрация веществ в клеточном соке и во внешней среде (в почве, водоемах) обычно не одинаковы. Если внутриклеточная концентрация веществ выше, чем во внешней среде, вода из среды будет диффундировать в клетку, точнее в вакуоль, с большей скоростью, чем в обратном направлении, т. е. из клетки в среду. Чем больше концентрация содержащихся в клеточном соке веществ, тем сильнее сосущая сила- сила, с которой клетка . При увеличении объема клеточного сока, вследствие поступления в клетку воды, увеличивается его давление на цитоплазму, плотно прилегающую к оболочке. При полном насыщении клетки водой она имеет максимальный объем. Состояние внутреннего напряжения клетки, обусловленное высоким содержанием воды и развивающимся давлением содержимого клетки на ее оболочку носит название тургора. Тургор обеспечивает сохранение органами формы (например, листьями, неодревесневшими стеблями) и положения в пространстве, а также сопротивление их действию механических факторов. С потерей воды связано уменьшение тургора и увядание.
Если клетка находится в гипертоническом растворе, концентрация которого больше концентрации клеточного сока, то скорость диффузии воды из клеточного сока будет превышать скорость диффузии воды в клетку из окружающего раствора. Вследствие выхода воды из клетки объем клеточного сока сокращается, тургор уменьшается. Уменьшение объема клеточной вакуоли сопровождается отделением цитоплазмы от оболочки - происходит плазмолиз.
В ходе плазмолиза форма плазмолизированного протопласта меняется. Вначале протопласт отстает от клеточной стенки лишь в отдельных местах, чаще всего в уголках. Плазмолиз такой формы называют уголковым.
Затем протопласт продолжает отставать от клеточных стенок, сохраняя связь с ними в отдельных местах, поверхность протопласта между этими точками имеет вогнутую форму. На этом этапе плазмолиз называют вогнутым.
Постепенно протопласт отрывается от клеточных стенок по всей поверхности и принимает округлую форму. Такой плазмолиз носит название выпук лого.
Если у протопласта связь с клеточной стенкой в отдельных местах сохраняется, то при дальнейшем уменьшении объема в ходе плазмолиза протопласт приобретает неправильную форму. Протопласт остается связанным с оболочкой многочисленными ни тями Гехта. Такой плазмолиз носит название судорожного.
При длительном нахождении клеток в растворе нитрата калия (15 мин. и более) цитоплазма набухает в удлиненных клетках, там, где протопласт не касается клеточных стенок, образуются так называемые колпачки цитоплазмы. Такой плазмолиз носит название колпа чкового.
Рис. Плазмолиз растительной клетки:
А - клетка в состоянии тургора; Б - уголковый; В - вогнутый; Г - выпуклый; Д - судорожный; Е - колпачковый.
1 - оболочка, 2 - вакуоль, 3 - цитоплазма, 4 - ядро, 5 - нити Гехта.
Если плазмолизированную клетку поместить в гипотонический раствор, концентрация которого меньше концентрации клеточного сока, вода из окружающего раствора будет диффундировать внутрь вакуоли. В результате увеличения объема вакуоли повысится давление клеточного сока на цитоплазму, которая начинает приближаться к стенкам клетки, пока не примет первоначальное положение - произойдетдеплазмолиз.
23. Поступление воды в растения. Влияние аэрации, температуры почвы и других внешних факторов на деятельность корневой системы.
Еще в 1864 г. Ю. Сакс установил, что поступление воды через корневые системы зависит от температуры. С понижением температуры скорость поступления воды резко сокращается. Это может оказать заметное влияние на растительный организм, особенно в осенний период, когда испарение идет еще достаточно интенсивно, а поступление воды задерживается из-за пониженной температуры почвы. В результате растения завядают и даже могут погибнуть от обезвоживания Причин, по которым понижение температуры вызывает уменьшение поступления воды, по-видимому, несколько:
1) повышается вязкость воды и, как следствие, снижается ее подвижность;
2) уменьшается проницаемость цитоплазмы для поды;
3) тормозится рост корней;
4) уменьшается скорость всех метаболических процессов.
Последнее обстоятельство, по-видимому, должно сказаться косвенно, через уменьшение поступления солей и, как следствие, торможение работы нижнего концевого двигателя (корневого давления). Торможение поступления йоды в корневую систему при действии пониженной температуры можно легко продемонстрировать в простом опыте. Так, если положить кусок льда на поверхность почвы в сосуде с растением, то в условиях интенсивной транспирации растение через два часа обнаружит признаки завядания. Если затем удалить лед, то растение снова приобретет тургор.
Снижение аэрации почвы также тормозит поступление воды. Это можно наблюдать, когда после сильного дождя все промежутки почвы заполнены водой и вместе с тем на солнце при сильном испарении растения завядают. Это связано С тем, что все условия, снижающие метаболизм (недостаток кислорода, избыток С02, дыхательные яды) снижают поступление ионов и, как следствие, уменьшают поступление воды. Вместе с тем исследования показали, что особенно резкое подавление поступления воды происходит при увеличении содержания С02. Возможно, избыток углекислого газа помимо ингибирования дыхания, повышает вязкость воды, снижает проницаемость цитоплазмы. Большое значение имеет содержание воды в почве, а также концентрация почвенного раствора. Естественно, вода поступает в корень только тогда, когда водный потенциал корня ниже (более отрицателен) водного потенциала почвы. В том случае, если почвенный раствор имеет более отрицательный осмотический потенциал, вода не только не будет поступать в корень, но будет выделяться из него. Особенное значение это имеет для засоленных почв. Именно поэтому растения, растущие на этих почвах (галофиты), имеют резко отрицательный осмотический потенциал.
Всякое уменьшение влажности почвы снижает поступление воды. Чем меньше воды в почве, тем с большей силой она удерживается и тем меньше ее водный потенциал. Для того чтобы в растение поступила вода, должен существовать градиент водного потенциала в системе почва — растение — атмосфера. Надо также учитывать, что уменьшение содержания воды в корне затрудняет ее дальнейшее продвижение к сосудам ксилемы. Это объясняется тем, что при уменьшении содержания воды сопротивление ее передвижению по клеткам корня растет. Подсыхающие клеточные оболочки оказывают значительное сопротивление передвижению воды. Сопротивление корневой системы передвижению воды при се недостатке настолько велико, что, по мнению Н. А. Максимова, это может снижать транспирацию, т. е. являться средством для ее регуляции. С физиологической точки зрения удобно выделить следующие формы почвенной влаги, различающиеся по степени доступности для растений. Гравитационная вода заполняет крупные промежутки между частицами почвы, она хорошо доступна растениям. Водный потенциал этой формы воды зависит от осмотической концентрации и составляет -0,1 бар. Однако, как правило, она легко стекает в нижние горизонты под влиянием силы тяжести, вследствие чего бывает в почве лишь после дождей. Капиллярная вода заполняет капиллярные поры в почве. Эта вода также хорошо доступна для растений, удерживается в капиллярах силами поверхностного натяжения и поэтому не только не стекает вниз, но и поднимается вверх от грунтовых вод . Пленочная вода окружает коллоидные частицы почвы. Вода из периферических слоев гидратационных оболочек может поглощаться клетками корня. Вместе с тем чем ближе к коллоидным частицам располагаются молекулы воды, тем с большей силой они удерживаются и, как следствие, менее доступны для растений. Гигроскопическая вода адсорбируется сухой почвой при помещении ее в атмосферу с относительной влажностью воздуха 95%.
31. Источники углерода для растений.
В ходе фотосинтеза растение вырабатывает из неорганических веществ необходимые для всего живого органические вещества. Диоксид углерода, содержащийся в воздухе, проникает в лист через особые отверстия в эпидермисе листа, которые называют устьицами в форме угольного ангидрида — СО2. При участии хлорофилла листа и солнечного света, угольный ангидрид разлагается с выделением О2.
К органическим веществам относятся все соединения углерода за исключением его оксидов и нитридов. В наибольшем количестве образуются при фотосинтезе такие органические вещества, как углеводы , аминокислоты и жирные кислоты Из неорганических веществ для синтеза всех этих соединений требуются вода и диоксид углерода. Растения могут поглощать эти элементы в форме их оксидов, нитрата и сульфата или в других, более восстановленных формах, таких, как аммиак или сероводород У наземных растений все неорганические соединения, за исключением углерода, поступают через корни. углерод растения получают из атмосферного воздуха, в котором средняя его концентрация составляет 0,03%. Диоксид углерода поступает в листья, а О2 выделяется из них через небольшие отверстия в эпидермисе, называемые устьицами. Открывание и закрывание устьиц регулируют особые клетки - их называют замыкающими - тоже зеленые и способные осуществлять фотосинтез. Когда на замыкающие клетки падает свет, в них начинается фотосинтез. Накопление его продуктов вынуждает эти клетки растягиваться. При этом устьичное отверстие открывается шире, и СО2 проникает к нижележащим слоям листа, клетки которых могут теперь продолжать фотосинтез. Устьица регулируют и испарение воды листьями, транспирацию, поскольку большая часть водяных паров проходит именно через эти отверстия. Водные растения добывают все необходимые им питательные вещества из воды, в которой живут. СО2 и ион бикарбоната тоже содержатся и в морской, и в пресной воде. Водоросли и другие водные растения получают их непосредственно из воды. Свет в фотосинтезе играет роль не только катализатора, но и одного из реагентов. Значительная часть световой энергии, используемой растениями при фотосинтезе, запасается в виде химической потенциальной энергии в продуктах фотосинтеза. Растения превращают диоксид углерода в кислород и для этого процесса необходим свет. Весь углерод, включенный растением в органические вещества, происходит из диоксида углерода.
32. Роль аденозинфосфатов и других макроэргических соединений
Группа природных веществ, молекулы которых содержат богатые энергией, или макроэргические связи; присутствуют во всех живых клетках и участвуют в накоплении и превращении энергии. Разрыв макроэргических связей в молекулах сопровождается выделением энергии, используемой для биосинтеза, транспорта веществ и других процессов жизнедеятельности.
Все известные макроэргические связи содержат фосфорильную (—РО3Н2) или ацильную группы и могут быть описаны формулой Х—Y, где Х — атом азота, кислорода, серы или углерода, а Y — атом фосфора или углерода. Реакционная способность макроэргических связей. связана с повышенным сродством к электрону атома Y, что обусловливает высокую свободную энергию гидролиза макроэргических связей, составляющую 6—14 ккал/моль.
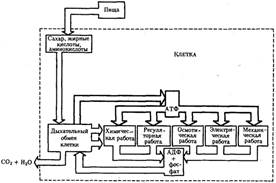
Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы. Одна из них осмотическая (поддержание перепадов концентрации веществ)
Осмос - диффузия (передвижение мол-л по градиенту конц-ции - из обл выс конц в обл низк конц) воды через полупрониц мембраны.
В раст кл-ке: Плазмолиз (когда жарко) - отток воды, содерж-е кл-ки сжим-ся и отходит от клет стенки. Деплазмолиз (прохл-но+полить) - кл-ки набух и прижим к клет стенке, подчин тургорному давл-ю (тургор - внутр гидростатич давл, вызывающее натяж-е кл стенки). Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором).
В жив кл-ке: изотонич р-р - норма, гипертон р-р - сморщив-е, гипотонич р-р - набух-е, затем лопаются-лизис.
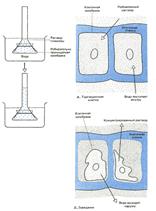
Рис. 1. Осмос в искусственной системе. Трубку, содержащую раствор глюкозы и закрытую с одного конца мембраной,пропускающей воду, но не пропускающей глюкозу, опускают закрытым концом в сосуд с водой. Вода может проходить через мембрану в том и в другом направлении; однако молекулы глюкозы в трубке мешают движению соседних молекул воды, и потому больше воды входит в трубку, чем выходит из нее. Раствор поднимается в трубке до тех пор, пока давление его столба не станет достаточным для того, чтобы вытеснять воду из трубки с такой же скоростью, с какой она поступает внутрь.
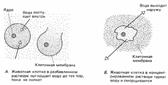
Осмос - процесс одностороннего проникновения молекул растворителя через полунепроницаемую мембрану в сторону большей конц. растворенного вещ-ва. отчего зависит осмос? во-первых, от общей концентрации всех растворенных частиц по обе стороны от мембраны, ну а во-вторых от давления, создаваемого каждым "р-ром" (понятие осмотического давления: такое давление на раствор, обусловленное стремлением системы (ну т.е. клетки) выровнить конц. р-ра в обеих средазх, разделенных мембраной). Наличие воды необходимо для норм. протекания всех ппроцессов, и именно благодаря осмосу происходит "обводнение" клеток и структур. У КЛЕТОК НЕТ СПЕЦ, МЕХАНИЗМА ДЛЯ НАСАСЫВАНИЯ И ОТКАЧИВАНИЯ ВОДЫ НЕПОСРЕДСТВЕННО! - поэтому приток и отток воды регулируется изменением конц. вещ-в. Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором).
Энергия АТФ, непосредственно или будучи перенесена на другие макроэргические соединения (например, креатинфосфат), в разнообразных процессах преобразуется в тот или иной вид работы. Одна из них осмотическая (поддержание перепадов концентрации веществ)
Осмос - диффузия (передвижение мол-л по градиенту конц-ции - из обл выс конц в обл низк конц) воды через полупрониц мембраны.
В раст кл-ке: Плазмолиз (когда жарко) - отток воды, содерж-е кл-ки сжим-ся и отходит от клет стенки. Деплазмолиз (прохл-но+полить) - кл-ки набух и прижим к клет стенке, подчин тургорному давл-ю (тургор - внутр гидростатич давл, вызывающее натяж-е кл стенки). Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором).
В жив кл-ке: изотонич р-р - норма, гипертон р-р - сморщив-е, гипотонич р-р - набух-е, затем лопаются-лизис.
Рис. 1. Осмос в искусственной системе. Трубку, содержащую раствор глюкозы и закрытую с одного конца мембраной,пропускающей воду, но не пропускающей глюкозу, опускают закрытым концом в сосуд с водой. Вода может проходить через мембрану в том и в другом направлении; однако молекулы глюкозы в трубке мешают движению соседних молекул воды, и потому больше воды входит в трубку, чем выходит из нее. Раствор поднимается в трубке до тех пор, пока давление его столба не станет достаточным для того, чтобы вытеснять воду из трубки с такой же скоростью, с какой она поступает внутрь.

Осмос - процесс одностороннего проникновения молекул растворителя через полунепроницаемую мембрану в сторону большей конц. растворенного вещ-ва. отчего зависит осмос? во-первых, от общей концентрации всех растворенных частиц по обе стороны от мембраны, ну а во-вторых от давления, создаваемого каждым "р-ром" (понятие осмотического давления: такое давление на раствор, обусловленное стремлением системы (ну т.е. клетки) выровнить конц. р-ра в обеих средазх, разделенных мембраной). Наличие воды необходимо для норм. протекания всех ппроцессов, и именно благодаря осмосу происходит "обводнение" клеток и структур. У КЛЕТОК НЕТ СПЕЦ, МЕХАНИЗМА ДЛЯ НАСАСЫВАНИЯ И ОТКАЧИВАНИЯ ВОДЫ НЕПОСРЕДСТВЕННО! - поэтому приток и отток воды регулируется изменением конц. вещ-в. Клеточная стенка способна растягиваться до определенного предела, после чего оказывает сопротивление - вытеснение воды из клеток происходит с такой же скоростью, с которой она в них поступает. (! прочность клеточной стенки не даёт раст. клеткам, в отличие от животных, лопнуть под напором).
Жизнедеятельность клетки характеризуется непрерывно протекающими в ней процессами обмена веществ, причем цитоплазма избирательно реагирует на воздействие разных факторов внешней среды. В поглощении и выделении веществ большую роль играют процессы диффузии и осмоса. Осмотическими называют явления, происходящие в системе, состоящей из двух растворов, разделенных полупроницаемой мембраной. В растительной клетке роль полупроницаемых пленок выполняют пограничные слои цитоплазмы: плазмалемма и тонопласт.
Плазмалемма - наружная мембрана цитоплазмы, прилегающая к клеточной оболочке. Тонопласт - внутренняя мембрана цитоплазмы, окружающая вакуоль. Вакуоли представляют собой полости в цитоплазме, заполненные клеточным соком - водным раствором углеводов, органических кислот, солей, белков с низким молекулярным весом, пигментов.
Концентрация веществ в клеточном соке и во внешней среде (в почве, водоемах) обычно не одинаковы. Если внутриклеточная концентрация веществ выше, чем во внешней среде, вода из среды будет диффундировать в клетку, точнее в вакуоль, с большей скоростью, чем в обратном направлении, т. е. из клетки в среду. Чем больше концентрация содержащихся в клеточном соке веществ, тем сильнее сосущая сила - сила, с которой клетка . При увеличении объема клеточного сока, вследствие поступления в клетку воды, увеличивается его давление на цитоплазму, плотно прилегающую к оболочке. При полном насыщении клетки водой она имеет максимальный объем. Состояние внутреннего напряжения клетки, обусловленное высоким содержанием воды и развивающимся давлением содержимого клетки на ее оболочку носит название тургора. Тургор обеспечивает сохранение органами формы (например, листьями, неодревесневшими стеблями) и положения в пространстве, а также сопротивление их действию механических факторов. С потерей воды связано уменьшение тургора и увядание.
Если клетка находится в гипертоническом растворе, концентрация которого больше концентрации клеточного сока, то скорость диффузии воды из клеточного сока будет превышать скорость диффузии воды в клетку из окружающего раствора. Вследствие выхода воды из клетки объем клеточного сока сокращается, тургор уменьшается. Уменьшение объема клеточной вакуоли сопровождается отделением цитоплазмы от оболочки - происходит плазмолиз.
В ходе плазмолиза форма плазмолизированного протопласта меняется. Вначале протопласт отстает от клеточной стенки лишь в отдельных местах, чаще всего в уголках. Плазмолиз такой формы называют уголковым
Затем протопласт продолжает отставать от клеточных стенок, сохраняя связь с ними в отдельных местах, поверхность протопласта между этими точками имеет вогнутую форму. На этом этапе плазмолиз называют вогнутым
Постепенно протопласт отрывается от клеточных стенок по всей поверхности и принимает округлую форму. Такой плазмолиз носит название выпуклого
Если у протопласта связь с клеточной стенкой в отдельных местах сохраняется, то при дальнейшем уменьшении объема в ходе плазмолиза протопласт приобретает неправильную форму. Протопласт остается связанным с оболочкой многочисленными нитями Гехта. Такой плазмолиз носит название судорожного
При длительном нахождении клеток в растворе нитрата калия (15 мин. и более) цитоплазма набухает в удлиненных клетках, там, где протопласт не касается клеточных стенок, образуются так называемые колпачки цитоплазмы. Такой плазмолиз носит название колпачкового
. Плазмолиз растительной клетки:
Если плазмолизированную клетку поместить в гипотонический раствор, концентрация которого меньше концентрации клеточного сока, вода из окружающего раствора будет диффундировать внутрь вакуоли. В результате увеличения объема вакуоли повысится давление клеточного сока на цитоплазму, которая начинает приближаться к стенкам клетки, пока не примет первоначальное положение - произойдет деплазмолиз.
Читайте также: