
Нервная регуляция мочеиспускания кратко
Обновлено: 02.07.2024
Введение Нейрогенные нарушения мочеиспускания являются типичным проявлением большинства неврологических заболеваний и наблюдаются у 10–90% больных в зависимости от характера и распространенности поражения нервной системы [1,3,7,10,15]. Эти расстройства носят функциональный характер и являются отражением процессов, происходящих в нервной системе (преимущественно в ее центральном отделе). Для каждого конкретного неврологического заболевания характерны индивидуальные симптомокомплексы и отличительные патогенетические механизмы, их формирующие [6]. Причины формирования стойких патологических рефлексов мочеиспускания заключаются в поражении центров мочеиспускания (ишемический инсульт, опухоли головного и спинного мозга), нарушении проведения нервных импульсов от рецепторов мочевого пузыря и уретры к корковым, подкорковым, стволовым или спинальным центрам мочеиспускания (рассеянный склероз, субкортикальная артериосклеротическая энцефалопатия), дегенеративных изменениях нервной системы и дисбалансе нейромедиаторов (болезнь Паркинсона, мультисистемная атрофия) [11,13,16].
Удержание мочи в мочевом пути, выделение мочи регулируется вегетативной нервной системой и соматической нервной системой. Их тесное взаимодействие обеспечивает физиологическую координацию деятельности мускулатуры мочевого пузыря, сфинктеров мочеиспускательного канала. Большое значение в процессе мочеиспускания играют поперечно-полосатые мышцы брюшного пресса, промежности, мочеполовой диафрагмы.
Произвольное и непроизвольное мочеисускание
Мочеиспускание осуществляется рефлекторным путем. Оно бывает произвольным и непроизвольным. Мочеиспускание происходит за счет сокращения и расслабления определенных мышечных групп, которые образуют стенки мочевого пузыря, сфинктеры шейки мочевого пузыря и уретры (внутренний сфинктер уретры и наружный сфинктер уретры). Гладкая мускулатура мочевого пузыря, состоящая из детрузора и внутреннего сфинктера, имеет вегетативную симпатическую и парасимпатическую иннервацию. Парасимпатические центры располагаются в боковых рогах 2 – 4 крестцовых (S 2 – S 4) сегментов спинного мозга. От боковых рогов спинного мозга преганглионарные волокна идут в составе передних корешков, затем отделяются от них в виде тазовых внутренностных нервов, вступают во внутриорганный узел мочепузырного сплетения. Затем потсганглионарные волокна заканчиваются в гладкой мускулатуре мочевого пузыря. Нервные импульсы парасимпатических волокон вызывают сокращение детрузора и расслабление внутреннего сфинктера мочевого пузыря. Это сопровождается опорожнением мочевого пузыря. Симпатические центры располагаются в области 1 - 2 поясничных (L1 – L 2) сегментов спинного мозга, откуда выходят симпатические волокна, идут в составе передних корешков, отделяются от них в виде белых соединительных ветвей, проходят без перерыва через поясничные узлы симпатического ствола и в составе брыжеечных нервов достигают нижнего брыжеечного узла. От нижнего брыжеечного узла постганглионарные волокна в составе подчревных нервов достигают гладкой мускулатуры мочевого пузыря. Импульсы от симпатических волокон расслабляют детрузор и сокращают внутренний сфинктер. В результате этого происходит задержка мочи.
Роль симпатической и парасимпатической нервной системы
Сарклиник считает, что в регуляции рефлекторной деятельности мочевого пузыря большую роль играет парасимпатическая нервная система и парасимпатическая иннервация. Симпатическая нервная система осуществляет преимущественно регуляцию просвета сосудов мочевого пузыря, а также иннервирует мышцы пузырного треугольника. Кроме нисходящих эфферентных вегетативных импульсов на гладкую мускулатуру мочевого пузыря существуют также восходящие афферентные воздействия на вегетативные спинальные центры и супрасегментарные отделы нервной системы на уровне ствола и подкорковых структур. Восходящая цепочка импульсов идет от интерорецепторов мочевого пузыря, идет в восходящем направлении, достигает сначала вегетативных центров спинного мозга, а далее гипоталамолимбических структур.
Произвольная иннервация при акте мочеиспускания
Произвольная иннервация при акте мочеиспускания осуществляется по следующему механизму. От рецепторов наружного сфинктера мочевого пузыря через половой нерв чувствительные волокна направляются в спинной мозг ко 2 – 4 крестцовым сегментам (S2 – S4), далее по задним и боковым канатикам достигают сводчатой извилины. Сводчатая извилина является сенсорной областью мочеиспускания. Далее по ассоциативным волокнам импульсы поступают в двигательную корковую зону мочевого пузыря, она располагается в парацентральной дольке рядом с проекцией стопы. Специалисты частной медицинской практики Сарклиник считают, что корковый центр мочеиспускания также располагается в предцентральной извилине около проекции мышц бедра и тормозит произвольное мочеиспускание. Нисходящие волокна от корковых центров мочеиспускания по системе пирамидного пути направляются к клеткам передних рогов спинного мозга на уровне 2 – 4 крестцовых сегментов (S2 – S4), далее идут вместе с передними корешками и достигают наружного сфинктера мочевого пузыря, обеспечивая позыв на мочеиспускание. Рецепторы наружного сфинктера мочевого пузыря имеют двустороннее корковое представительство, поэтому при одностороннем спинальном поражении выше вегетативных непроизвольно-рецепторных центров произвольное мочеиспускание чаще не нарушено.
Механизм возникновения недержания мочи, почему происходит, бывает недержание мочи?
Механизмы возникновения разнообразных видов недержания мочи основываются на физиологических механизмах рефлекторного выделения мочи в раннем послеродовом периоде. В течение первых 6 месяцев жизни ребенка мочеиспускание осуществляется автоматически по типу безусловного рефлекса с частотой от 15 до 20 раз в сутки. С 6 месяцев до 12 месяцев в дневное время дети перед мочеиспусканием начинают проявлять определенное беспокойство. Они потягиваются, кряхтят, произносят различные звуки, елозят. Во втором полугодии жизни возникает субъективное ощущение позыва на мочеиспускание и начинает вырабатываться условный рефлекс. В ночное время дети в этом возрасте продолжают мочиться в постель, что свидетельствует о функционировании только спинальных механизмов. Постепенно потребность к мочеиспусканию на короткий период времени может подавляться, и формируется навык опрятности сначала в дневное время, а позже и в ночное время. В возрасте 2 года здоровый ребенок обычно уже не мочится непроизвольно ни ночью, ни днем. При тяжелых соматических заболеваниях может наступать временная утрата произвольного мочеиспускания.
Недержание мочи может быть связано с изменениями в самом мочеиспускательном канале, а также сфинктерном аппарате, что приводит к нарушению функции замыкательного аппарата. Недержание мочи может быть связано с ослаблением и дислокацией связочного аппарата уретровезикального сегмента и неизмененного мочеиспускательного канала.
Лечение недержания мочи в Саратове
Сарклиник (Саратов, Россия) проводит лечение недержания мочи в Саратове. Если у Вас частое затрудненное мочеиспускание, боль после или во время мочеиспускания, частое мочеиспускание, жжение, рези, частые позывы, кровь, болезненное учащенное мочеиспускание, проблемы с мочеиспусканием, ночной энурез, обращайтесь в Сарклиник.
Высшие центры контроля мочеиспускания: цикл мочеиспускания, его нарушения
Нижние мочевыводящие пути разделены на два отдела—вместилище мочи (мочевой пузырь), а также собственно выводящие пути—уретру и наружный сфинктер уретры, действующих скоординировано для хранения и выведения мочи. На уровне спинного мозга парасимпатические нейроны крестцовых сегментов вызывают сокращение мочевого пузыря (детрузора). Симпатические нейроны поясничных сегментов угнетают парасимпатические влияния и позволяют мочевому пузырю наполниться.
Двигательные нейроны крестцовых сегментов, иннервирующие наружный сфинктер уретры, вносят вклад в функционирование обеих систем: при их возбуждении облегчается накопление мочи, при их торможении—облегчается ее выведение. Взаимодействие между этими системами происходит на различных уровнях ЦНС.
Мостовой центр контроля мочеиспускания (ядро Баррингтона) находится в парамедианной мостовой ретикулярной формации с каждой стороны, соединяясь по срединной линии. Отсюда отходят волокна крупноклеточных нейронов к отвечающим за мочеиспускание парасимпатическим нейронам в сегментах S2-S4 спинного мозга. Возбуждение центра контроля мочеиспускания приводит к акту мочеиспускания посредством повышения внутрипузырного давления (в результате сокращения гладких мышц стенки мочевого пузыря) и одновременного расслабления поперечно-полосатых мышц наружного уретрального сфинктера (вследствие возбуждения ГАМК-ергических вставочных нейронов ядра Онуфа, образующих синапсы с крестцовыми сегментами спинного мозга).
Аксоны этих двигательных нейронов проходят в составе половых нервов, иннервирующих наружный уретральный сфинктер. (В более латеральном отделе ретикулярной формации моста расположена L-зона, имеющаяся у млекопитающих, от которой волокна направляются к ядру Онуфа и вызывают сокращение наружного сфинктера уретры. Мостовой центр контроля мочеиспускания обозначают как М-зону, однако его взаимодействие с L-зоной вызывает сомнения.)
На более высоких уровнях к клеткам латерального отдела правого ОВСВ подходят восходящие волокна от задних рогов крестцовых сегментов, а от них—возбуждающие волокна к островку, который генерирует чувство наполненности мочевого пузыря и передает возбуждение на медиальные отделы лобной коры. Возбуждение латерального отдела ОВСВ осуществляет также правый гипоталамус. Часть спиноретикулярных волокон от крестцовых сегментов возбуждает L-зону. Другие волокна переключаются через таламус на клетки правой передней поясной коры (ППК), возбуждающиеся в ситуациях, требующих концентрации внимания.
Такое доминирование правых отделов может быть связано с эмоциональными аспектами мочеиспускания. При функциональном исследовании мозга было установлено, что дальнейшее заполнение жидкостью уже полного мочевого пузыря приводит к активации островка и дорсальной передней поясной коры/дополнительной моторной области. Когда мочевой пузырь не наполнен, заполнение жидкостью приводит к активации среднего мозга и парагиппокампальной области, осуществляющих бессознательный контроль афферентных сигналов от мочевого пузыря.
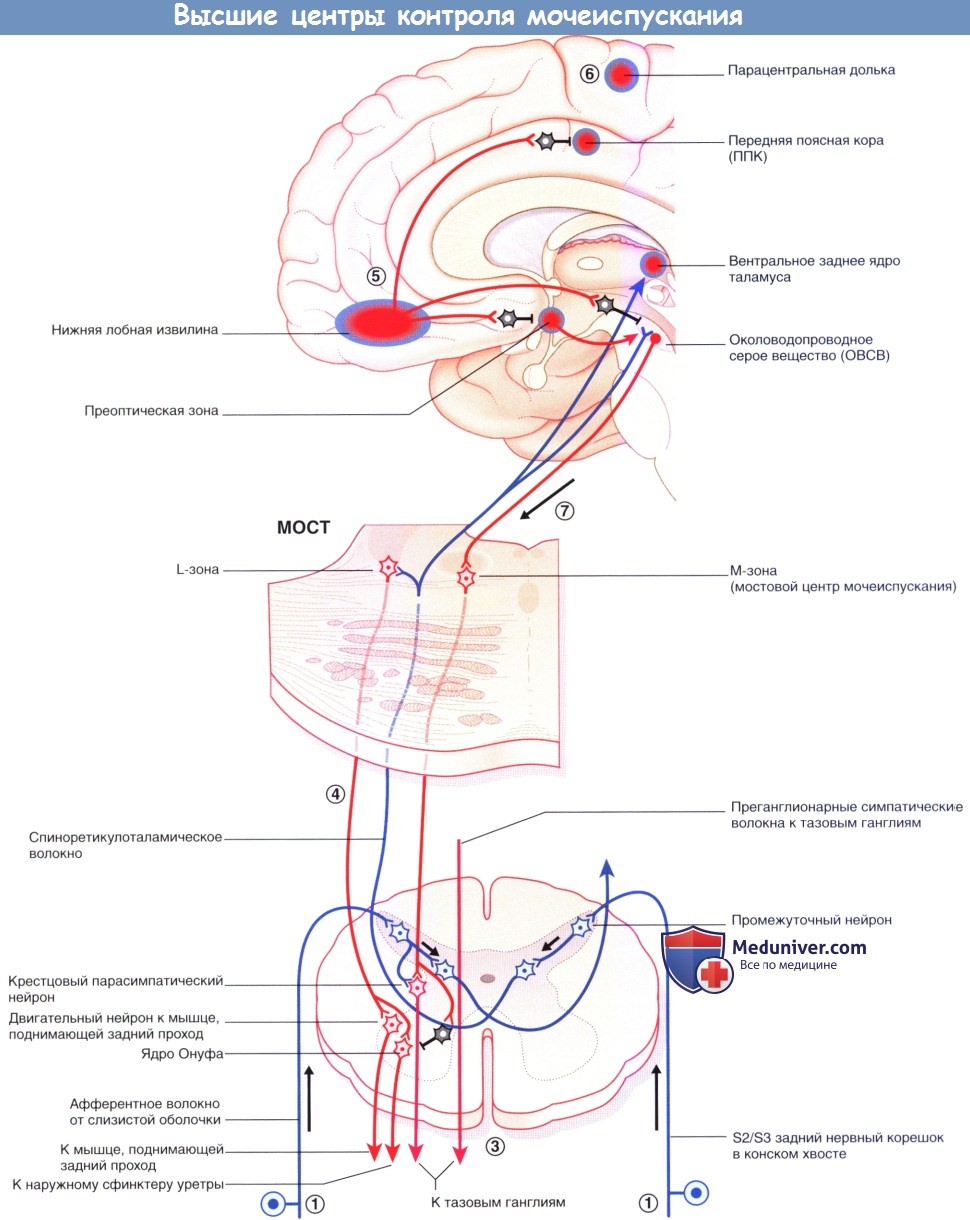
а) Цикл мочеиспускания:
1. Когда мочевой пузырь заполнен наполовину, восходящие волокна от рецепторов растяжения в детрузоре и слизистой оболочке в мочепузырном треугольнике передают эту информацию по спиноретикулярным волокнам через таламус к мосту, среднему мозгу и мозжечку.
2. От коры островка информация поступает в центр принятия решений в медиальной лобной коре, которая получает данные о степени наполнения мочевого пузыря.
3. Активация симпатической системы увеличивается постепенно, позволяя увеличить растяжимость мочевого пузыря (через β2-адренорецепторы). При взаимодействии с α2-адренорецепторами происходит торможение парасимпатических нейронов.
4. Спиноретикулярные волокна, образующие синапсы с L-зоной моста, возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая, таким образом, тонус наружного сфинктера уретры.
5. При полном заполнении возникает ощущение непреодолимого желания. При неподходящем времени или месте активируется отдел медиальной лобной извилины. Эта область тормозит ППК, уменьшая уровень ее активности путем возбуждения ее тормозных вставочных нейронов. Аналогичным образом через переключение в гипоталамусе и среднем мозге происходит торможение пре-оптической области и ОВСВ посредством возбуждения тормозных нейронов.
6. Последняя, но кратковременная мера —произвольное сокращение мышц тазовогодна. Команду на сокращение посылает лобная кора к промежностному отделу в медиальной области моторной коры парацентральной дольки.
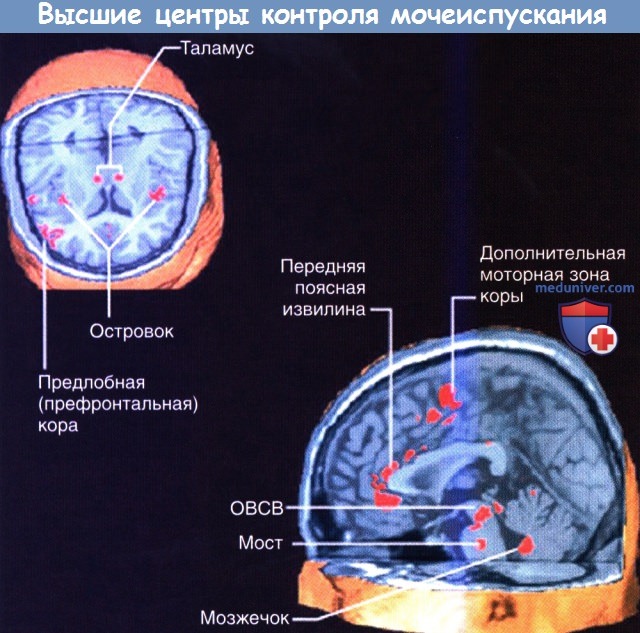
Показаны высокоактивные области при функциональной магнитно-резонансной томографии (фМРТ) в фазу наполнения мочевого пузыря в цикле мочеиспускания.
ОВСВ — околоводопроводное серое вещество. Мост — подразумевается мостовой центр контроля мочеиспускания.
б) Роль моноаминов. К двигательным и чувствительным ядрам спинного мозга, иннервирующим мочевой пузырь, подходит огромное количество серотонинергических нисходящих волокон от большого ядра шва (БЯШ) продолговатого мозга. Доказано, что растяжение мочевого пузыря приводит к стимуляции БЯШ (путем спиноретикулярной активации ОВСВ). Кратко излагая, контроль мочеиспускания на низшем уровне означает, что БЯШ создает тонус мочевого пузыря, направленный, прежде всего, на его наполнение.
Норадренергические нисходящие волокна из голубого пятна к передним рогам спинного мозга усиливают действие глутамата, локально вырабатываемого из клеток ядра Онуфа, увеличивая, тем самым, тонус наружного сфинктера в фазу наполнения.
G-белковые мускариновые рецепторы, активируемые постганглионарными волокнами тазовых ганглиев, в большом количестве присутствуют в детрузоре мочевого пузыря. Соответственно, препаратами выбора в данном случае служат блокаторы м-холинорецепторов. Однако появление антихолинергических побочных эффектов, таких как сухость во рту и запор, может потребовать отмены терапии.
В последние годы для лечения гиперактивности детрузора все чаще используют ботулотоксин. Известно, что он нарушает взаимодействие между холинергическими синаптическими везикулами и мышечными волокнами (как гладкими, так и поперечно-полосатыми), делая синаптическую передачу неэффективной. Гибкий цистоскоп проводят через уретру и выполняют многочисленные небольшие инъекции ботокса в стенку мочевого пузыря. Для получения долговременного результата процедуру повторяют 2 раза в год.

Хранение мочи и периодическое опорожнение мочевого пузыря (МП) зависят от координированной работы гладких и поперечнополосатых мышц, условно разделенных на два структурно функциональных отдела: резервуара (мочевой пузырь) и выпускной системы, состоящей из шейки МП, уретры и наружного сфинктера мочеиспускательного канала. За координированную работу вышеперечисленных структур отвечают сложные нейронные системы управления, расположенные в периферических ганглиях, спинном и головном мозге.
Периферическая иннервация нижних мочевыводящих путей (НМП)
Обеспечение сознательного контроля мочеиспускания требует сложных взаимодействий между вегетативным (симпатика и парасимпатика) и соматическим отделами нервной системы.
Симпатические нервные волокна берут свое начало из боковых рогов грудопоясничного отдела (спинномозговой центр Якобсона) и ганглиях нижнего брыжеечного сплетения и достигают МП в составе подчревного нерва. Симпатические постганглионарные нервы выделяют норадреналин, который активирует β-адренергические рецепторы, ингибирующие сокращение детрузорной мышцы, и α-адренергические рецепторы, возбуждающие мускулатуру уретры и шейки МП. Симпатическое воздействие приводит к расслаблению мышцы, изгоняющей мочу, сокращению мускулатуры уретры и шейки МП и ингибированию интрамуральных ганглиев МП.
Центральный отдел парасимпатической иннервации МП располагается в промежуточных ядрах крестцовых сегментов. Холинергические преганглионарные волокна из промежуточных ядер посылают свои аксоны через тазовые нервы к ганглиозным клеткам тазового сплетения и интрамуральным нейронам в стенке мочевого пузыря. Ганглиозные клетки в свою очередь возбуждают детрузор мочевого пузыря, что приводит к его сокращению с последующим опорожнением. Основными медиаторами парасимпатической нервной системы тут являются ацетилхолин и другие нехолинергические медиаторы. Ацетилхолин действует опосредованно, возбуждая M3-холинорецепторы клеток детрузора. Парасимпатические нервные окончания в нервно-мышечных синапсах и в парасимпатических ганглиях тоже имеют холинорецепторы, возбуждение этих рецепторов на нервных окончаниях может усиливать (через рецепторы М1) или подавлять (через рецепторы М4) высвобождение медиаторов в зависимости от интенсивности нейронного возбуждения. Основной нехолинергический медиатор это АТФ, который активирует внутриклеточную пуринергическую систему через возбуждение P2X рецепторов и тоже способствует сокращению детрузора. Парасимпатические волокна вызывают расслабление ГМК уретры путем высвобождения оксида азота (NO).
Аксоны соматических двигательных нейронов передних рогов крестцовых сегментов S2–S4 (ядро Онуфа) проходят в срамном нерве и иннервируют поперечно-полосатые мышцы наружного уретрального сфинктера. Нейроны более медиально расположенного моторного ядра на том же уровне позвоночника иннервируют мускулатуру тазового дна.
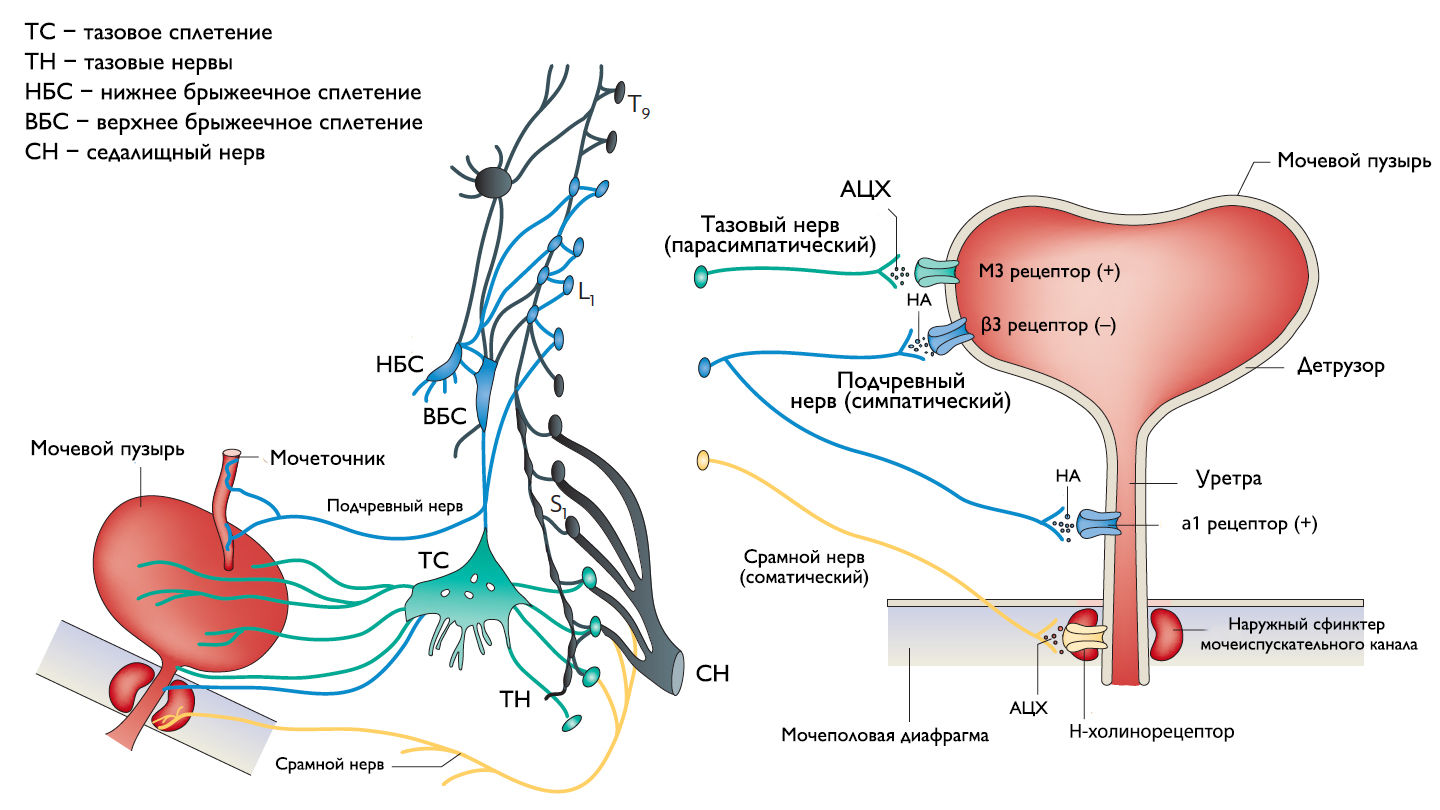
Рисунок 1 | Иннервация нижних мочевыводящих путей.
Центральные нервные пути и центры, регулирующие мочеиспускание
Специфический и неспецифический восходящий супраспинальный сенсорный путь
Одни спинномозговые промежуточные нейроны посылают восходящие волокна к определенным областям моста и среднего мозга, участвующим в мочеиспускании. Другие промежуточные нейроны передают информацию из нижних мочевыводящих путей в структуры переднего мозга, включая таламус и гипоталамус. Спиноталамический и спиногипоталамический тракты хотя и не играют главную роль в мочеиспускании, но могут включаться в сознательный контроль полноты мочевого пузыря. Чувствительные зоны коры ГМ через спиноталамический тракт информируются о состоянии наполнения МП.
Мостовой центр мочеиспускания (МЦМ) и его нисходящие спинномозговые двигательные пути
Впервые центр управления мочеиспусканием был открыт в дорсальной части моста Баррингтоном в 1925 году и с тех пор называется мостовым центром мочеиспускания или ядром Баррингтона. МЦМ располагается в области покрышки моста. Нейроны МЦМ имеют нисходящие возбуждающие синаптические контакты с клетками парасимпатических преганглионарных мотонейронов, иннервирующих постганглионарные клетки мочевого пузыря. Электрическая и химическая стимуляция МЦМ у крыс и кошек инициирует сокращение мочевого пузыря и расслабляет сфинктер уретры имитируя нормальное мочеиспускание. Цикл рефлекса мочеиспускания состоит из трех фаз, контролируемых различными центральными механизмами: фаза реализации безопасной среды — для начала мочеиспускания человеку необходимо осознание, что окружающие обстановка комфортна; фаза релаксации наружного уретрального сфинктера; и фаза сокращения мышцы, выталкивающей мочу. Процесс нормального мочеиспускания невозможен без какой-либо из этих фаз. МЦМ является командным центром мочеиспускания, который контролирует последовательное переключение фазы расслабления наружного уретрального сфинктера на фазу сокращения детрузорной мышцы.
Мостовой центр удержания мочи (МЦУ) и его нисходящие спинномозговые двигательные пути
Его роль заключается в расслаблении детрузора и сокращении наружного уретрального сфинктера. МЦУ располагается вентролатеральней МЦМ. Синапсы волокон МЦУ возбуждают ядро Онуфа в крестцовых сегментах спинного мозга, повышая таким образом тонус наружного сфинктера уретры. Стимуляция области МЦУ останавливает мочеиспускание, возбуждает мышцы тазового дна и сокращает уретральный сфинктер. Наоборот, двусторонние поражения МЦУ вызывают недержание мочи, чрезмерную детрузорную активность, невозможность хранения мочи и снижение тонуса уретрального сфинктера. На сегодняшний день нет анатомических доказательств связи между МЦУ и МЦМ, и было высказано предположение, что эти центры функционально независимы.
Роль кортикальных областей
Наиболее частые симптомы поражения кортикальных областей ГМ это поллакиурия и ургентное недержание мочи. Поэтому Andrew и Nathan выдвинули гипотезу, что отсоединение лобной или передней поясной извилины от гипоталамуса приводит к непроизвольному началу мочеиспускания [3]. Действительно, префронтальная кора головного мозга человека и передняя поясная извилина активируются во время мочеиспускания [4].
Мозжечок и базальные ганглии
Существует ряд исследований о том, что мозжечок и базальные ганглии оказывают в основном ингибирующее действие на мочевой пузырь. Мозжечковая патология приводит к увеличению частоты мочеиспускания и ургентному недержанию мочи. Симптомы гиперактивного мочевого пузыря также встречаются при болезни Паркинсона. Поскольку нет прямых связей этих областей с МЦМ, ингибирующее влияние, вероятно, косвенное через структуры переднего и среднего мозга.
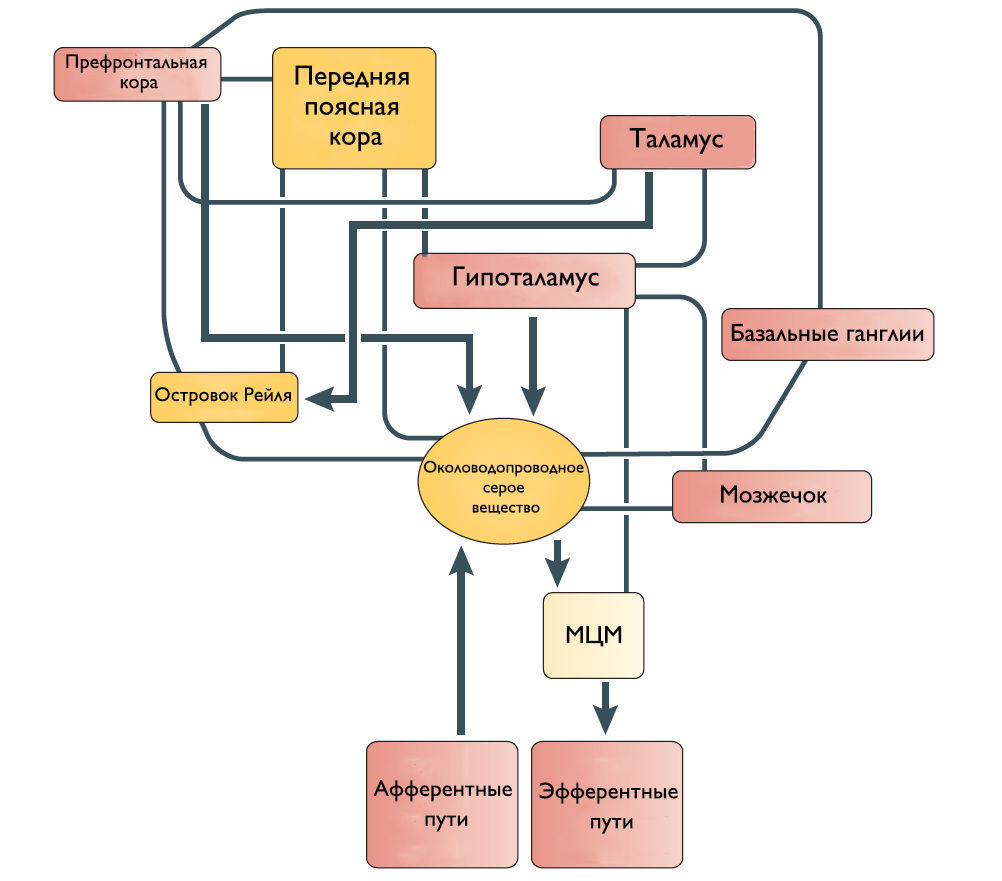
Рисунок 2 | Предположительное схематическое изображение связей между различными структурами переднего мозга и ствола мозга, которые участвуют в контроле мочеиспускания.
Читайте также: