
Нейтральная теория молекулярной эволюции кратко
Обновлено: 05.07.2024
Теория была разработана годов. Теория нейтральной эволюции хорошо согласуется с фактом постоянной скорости закрепления мутаций на молекулярном уровне, что позволяет, к примеру, оценивать время расхождения жизни на Земле. Дискуссия ведётся касательно доли мутаций, имеющих приспособительное значение. Большинство биологов признают ряд результатов теории нейтральной эволюции, хотя и не разделяют некоторые сильные утверждения, первоначально высказанные М.Кимурой.
| Это заготовка статьи по биологии. Вы можете помочь проекту, исправив и дополнив её. Это примечание по возможности следует заменить более точным . |
Литература
- Кимура М. Молекулярная эволюция: теория нейтральности. М., Мир, 1985
Материалы сообщества доступны в соответствии с условиями лицензии CC-BY-SA, если не указано иное.
Согласно нейтральной молекулярной теории эволюции, большинство генетических изменений не имеют адаптивного значения, а в небольших популяциях естественный отбор не играет решающей роли. Автор Quanta Magazine просто (и с картинками) объясняет, в чем суть этой теории и как она повлияла на биологию, экологию и другие, далекие от этих наук сферы жизни.
Последний раз голубых тигров видели в 1953 году. Потом они превратились в легенду. И даже единственная сохранившаяся шкура не доказывает, что этот вид когда-либо существовал.
Сразу напрашивается вывод: голубой окрас тигров был обусловлен определенным генетическим изъяном, из-за которого они не смогли конкурировать со своими оранжевыми сородичами. Но, скорее всего, исчезновение голубых тигров никак не связано с их диковинным окрасом, по стечению обстоятельств возникшим в небольшой популяции, численность которой на тот момент снижалась.
Эта теория кажется очень простой, но ее влияние на генетику, экологию и эволюционную биологию трудно переоценить.
Не совсем нейтральная теория
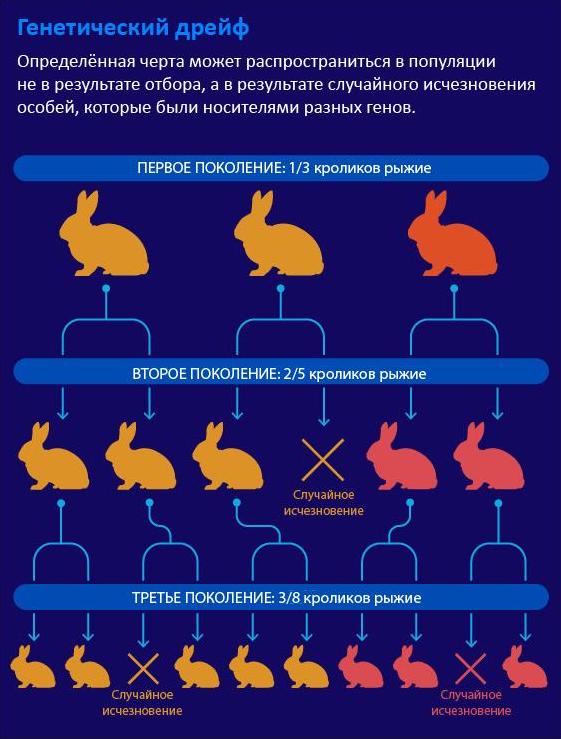
Процесс случайных изменений вариантов генов (аллелей) в популяции называется дрейфом генов . Хотя сегодня дрейф генов считается главной движущей силой эволюции, до 1960-х годов биологи объясняли разнообразие действием отбора: вредоносные гены, которые препятствуют продолжению рода, со временем исчезают ( отрицательный отбор ); а полезные гены, способствующие умножению потомства, сохраняются ( позитивный отбор ) — всё согласно принципу естественного отбора, изложенному Чарлзом Дарвином и Альфредом Расселом Уоллесом.
Затем секвенирование белков показало намного больше наследственных изменений в пределах популяций, чем ожидалось. Некоторые ученые засомневались, что все эти гены одновременно подвергались отбору.
В 1968 году знаменитый генетик Мотоо Кимура предложил иное объяснение, которое позже получило название нейтральной теории молекулярной эволюции. Кимура считал, что большинство генетических изменений не увеличивают и не уменьшают шансы видов на выживание и произведение потомства. Соответственно, разнообразие обусловливается преимущественно не отбором, а волей случая.
Изложенная Кимурой теория вызвала множество споров, так как отрицала ключевую роль отбора в эволюции. Но геномная революция в конце XX века и распространение секвенирования ДНК подтвердили правоту японского исследователя.
С тех пор нейтральная теория молекулярной эволюции стала нулевой гипотезой в генетике.
Размер популяции имеет значение
Представьте себе популяцию из десяти птиц: одной красной, одной зеленой и восьми коричневых. Цвет не является ни преимуществом, ни недостатком, поэтому все птицы имеют равные шансы на произведение потомства. Затем во время торнадо погибают шесть коричневых птиц. Теперь половину популяции составляют коричневые птицы, четверть — красные и еще одну четверть — зеленые. Случайное событие вызвало существенное изменение разнообразия. Это и есть дрейф генов.
Если бы было 98 коричневых птиц, а красных и зеленых только по одной, торнадо мало что бы изменило. Даже если бы уцелело лишь 38 коричневых и две яркие птицы, 95% птиц в популяции по-прежнему были бы коричневыми.
Прелесть нейтральной теории отчасти заключается в ее математической простоте, благодаря которой генетики получили возможность заглянуть в глубь времен: если генетическая изменчивость носит нейтральный характер, то можно вычислить размер популяции в прошлом и определить возраст последнего общего предка.
Но чтобы расчеты были точными, ученые должны учитывать не только случайные изменения, но и отбор. В этом, по словам Йохри, состоит главная трудность, так как в математике с 1970-х годов практически не было новых открытий.
В статье, опубликованной в журнале Genetics в мае, Йохри и ее коллеги предложили новую статистическую модель, учитывающую одновременно нейтральность и отрицательный отбор.
Видеть лес, а не деревья
Несмотря на то, что в популяционной генетике нейтральная теория считается общепринятой, она по-прежнему вызывает споры в других областях — в первую очередь в экологии. Одна из основ экологии гласит, что каждый вид занимает определенную нишу; чем больше ниш — тем больше видов.
Стивен Хаббелл из Калифорнийского университета в Лос-Анджелесе адаптировал теорию Кимуры для экологии.
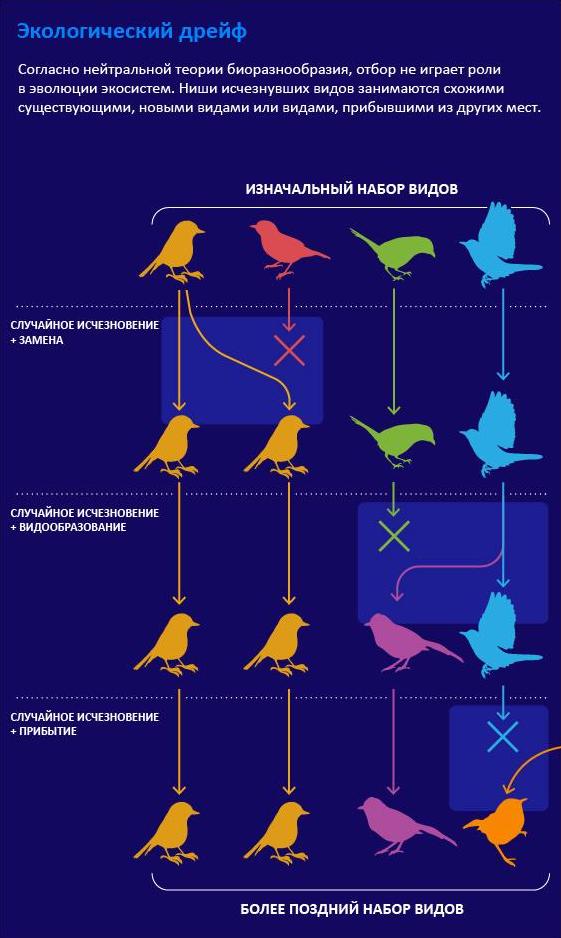
Эта теория противоречит теории Дарвина, и многие биологи подвергают ее сомнению. Согласно нейтралистам, не имеет значения, с какими видами соперничает определенный организм: зарянка конкурирует за червей не только с другими зарянками, но и с дроздами; а каждое дерево конкурирует за солнечный свет со всеми остальными деревьями. В итоге выживание видов зависит от случайных событий.
В экспериментах нейтральная теория далеко не всегда помогает точно предсказать состав экосистем, но многие экологи по-прежнему находят ее полезной в качестве нулевой гипотезы. Другие рассматривают ее как одну сторону медали, так как нейтральные силы и силы отбора всегда действуют вместе.
От генов к детским именам
Влияние нейтральной теории не ограничивается экологией. Она с успехом применяется для объяснения разнообразия во многих других сферах — от криптовалют до детских имен, поскольку представляет собой очень элегантную модель, объясняющую возникновение и исчезновение вариантов даже при отсутствии функциональных различий между ними.
Но влияние нейтральных сил также часто преувеличивается.
В статье, опубликованной в мае в журнале Nature Human Behaviour, Леруа и его коллеги описывают так называемый нейтральный синдром — склонность отвергать влияние сил отбора только потому, что разнообразие можно объяснить нейтральностью.
Авторы утверждают, что нужны более эффективные тесты, чтобы отделить влияние нейтральных сил от действия отбора.
Тем не менее Леруа не собирается полностью отказываться от нейтральной теории.
Нейтральная теория молекулярной эволюции (Нейтрализм Кимуры) — теория эволюции, которая утверждает, что подавляющее большинство мутаций на молекулярном уровне носит нейтральный по отношению к естественному отбору характер. Как следствие, значительная часть внутривидовой изменчивости (особенно в малых популяциях) объясняется не действием отбора, а случайным дрейфом мутантных аллелей, которые нейтральны или почти нейтральны.
Теория была разработана Мото Кимурой в конце 1960-х годов. Теория нейтральной эволюции хорошо согласуется с фактом постоянной скорости закрепления мутаций на молекулярном уровне, что позволяет, например, оценивать время дивергенции видов.
Теория нейтральной эволюции не отрицает решающей роли естественного отбора в эволюции. Дискуссия ведется по части мутаций, имеющих приспособительное значение. Большинство биологов признает ряд результатов теории нейтральной эволюции, хотя и не разделяет некоторые сильные утверждения, сначала высказанные М. Кимурой.
Создание теории
Изучая скорости аминокислотных замен в белках, Кимура обратил внимание на несоответствие данных, полученных им и ранее — Джоном Холдейном: скорость замен на геном на поколение для млекопитающих превышала оценку Холдейна в несколько сотен раз. Получалось, что для поддержания постоянной численности популяции при одновременном сохранении отбором мутантных замен, появляющихся с такой высокой скоростью, каждый родитель должен был оставлять около 22 000 потомков.
Метод электрофореза позволил выявить полиморфизм белков. Для 18 случайно выбранных локусов Drosophila pseudoobscura средняя гетерозиготность, приходящаяся на локус, составила около 12%, а доля полиморфных локусов — 30%. По данным Ф. Аялы, в генофонде природных популяций различных видов организмов полиморфные присутствует в 20-50% локусов.
Для объяснения популяционного полиморфизма Рональдом Фишером была разработана модель балансирующего отбора, основанная на селективной преимуществу гетерозигот. В то же время уровень гетерозиготности большинства организмов оценивался в среднем в 7-15%. А поскольку в популяциях тысячи аллелей, продуцируют полиморфные белки, невозможно утверждать, что все они обладают адаптивной ценностью.
В 1957 году в Холдейн показал математически, что в популяции не может заменяться одновременно более 12 аллелей более приспособленными аллелями без того, чтобы ее репродуктивная численность не упала до нуля.
Все эти соображения и натолкнули Кимуру на мысль, что большинство нуклеотидных замен должно быть селективно нейтральными и фиксироваться дрейфом генов. Соответствующие полиморфные аллели поддерживаются в популяции балансом между мутационным давлением и случайным отбором. Все это было изложено Кимурой в его первой публикации по нейтральной эволюции, названной так из-за нейтральность по отношению к естественному отбору. В дальнейшем появилась целая серия статей Кимуры, в том числе в соавторстве, а также обобщающая монография.
В этих трудах экспериментальные данные молекулярной биологии сочетались со строгими математическими расчетами, осуществленными самим автором на основе разработанного им математического аппарата. В книге, наряду с рассмотрением доводов в пользу новой теории, Кимура останавливается и на возможных оговорках и критики в ее адрес.
В качестве своих предшественников Кимура указывает также на Джеймса Ф. Кроу, с которым он сам активно сотрудничал, и Алана Робертсона. В поддержку теории свидетельствовали полученные позже данные о том, что наиболее распространенными эволюционными изменениями на молекулярном уровне является синонимичные замены, а также нуклеотидные замены в интронов (некодирующих участках ДНК). Все эти публикации породили острую дискуссию.
После интенсивных обсуждений и многочисленных исследований, порожденных нейтралистичною концепцией, интерес к ней в 1990-е годы годы заметно спал. Это произошло, во многом, из-за ограниченного применения теории, поскольку серьезный эволюционный прогресс по нейтралистським сценарию требовал бы огромных промежутков времени и непомерно больших количеств генов. Поэтому успех теории в рамках популяционной генетики и молекулярной биологии не распространился на дисциплины, изучающие макроэволюционные преобразования.
Классическая теория популяционной генетики, содержательно основанная на синтетической концепции эволюции, интенсивно развивалась до 1960-х годов, до тех пор, пока не возникли трудности интерпретации экспериментальных данных молекулярной биологии. В лекции 1 я уже отмечал, в 1950-1960-х годах произошла революция в молекулярной биологии. Была определена структура ДНК, расшифрован генетический код, ученые установили общие принципы работы молекулярно-генетической системы живой клетки.
Интенсивные исследования молекулярной биологии привели к серьезным результатам, касающимся биологической эволюции: была оценена скорость аминокислотных замен в белках, а также получены оценки, характеризующие полиморфизм белков.
Анализируя экспериментальные данные, М.Кимура обнаружил, что когда он пытался объяснить эти эксперименты на основе селекции благоприятных мутаций путем Дарвиновского отбора, то возникли серьезные затруднения. В своей книге [6] Кимура подробно описывает идеи, послужившие основанием для изобретения теории нейтральности. Например, в некоторых своих оценках, основанных на Дарвинском отборе, он получил, что для объяснения экспериментальных данных нужно потребовать, чтобы каждая особь в процессе эволюции давала 22 000 потомков. И для того, чтобы проинтерпретировать данные по молекулярной эволюции белков, Кимура предложил теорию нейтральности [6,9].
Основное предположение этой теории состоит в следующем: на молекулярном уровне мутации (замены аминокислот или нуклеотидов) преимущественно нейтральны или слабо вредны (существенно вредные мутации также возможны, но они элиминируются из популяции селекцией). Это предположение согласуется с экспериментально наблюдаемой скоростью аминокислотных замен и с тем фактом, что скорость замен в менее важных частях белков значительно больше, чем для активных центров макромолекул.
Используя математические методы популяционной генетики, Кимура получил ряд следствий теории, которые находятся в довольно хорошем согласии с данными молекулярной генетики [6].
Математические модели теории нейтральности существенно стохастические, т.е. относительно малая численность популяции играет важную роль в фиксации нейтральных мутаций. См. примеры расчетов, приведенных выше.
Но если молекулярные замены преимущественно нейтральны, как возможна прогрессивная эволюция? Чтобы ответить на этот вопрос, Кимура использует концепцию дупликации генов, развитую С.Оно [10]. Согласно теории Кимуры, дупликация генных участков создает дополнительные, избыточные ДНК-последовательности, которые в свою очередь дрейфуют далее за счет случайных мутаций, предоставляя тем самым сырой материал, из которого могут возникать новые, биологически значимые гены (Рис.1).

Рис. 1. Иллюстрация к механизму прогрессивной эволюции в теории нейтральности. Схема появления нового биологически значимого белка. Показаны участки ДНК ( Ii ) и кодируемые ими белки ( Ei ). a) ген I1 кодирует белок E1, b) дупликация гена I1, новый участок (справа) кодирует тот же белок E1, c) случайный дрейф правого участка, d) возникновение нового биологически значимого белка E2 кодируемого участком ДНК I2 .
Заключая наш сжатый обзор теории нейтральности, процитируем пять принципов этой теории [6]. Первые четыре из них – эмпирические, а пятый установлен теоретическим путем.
Скорость эволюции любого белка, выраженная через число аминокислотных замен на сайт в год, приблизительно постоянна и одинакова в разных филогенетических линиях, если только функция и третичная структура этого белка остаются в основном неизменными. Функционально менее важные молекулы и их части эволюционируют (накапливая мутационные замены) быстрее, чем более важные. Мутационные замены, приводящие к меньшим нарушениям структуры и функции молекулы (консервативные замены), в ходе эволюции происходят чаще тех, которые вызывают более существенное нарушение структуры и функции этой молекулы. Появлению нового в функциональном отношении гена всегда должна предшествовать дупликация гена. Селективная элиминация вредных мутаций и случайная фиксация селективно нейтральных или очень слабо вредных мутаций происходят в ходе эволюции гораздо чаще, чем положительный дарвиновский отбор благоприятных мутаций.
4. Другие модели, характеризующие общие закономерности эволюции
Теория нейтральности – одна из наиболее разработанных общих теорий эволюции. Однако есть ряд моделей и концепций, также характеризующих эволюцию на молекулярном уровне, которые в основном дополняют теорию нейтральности. Отметим наиболее известные из них.
В работах Д.С.Чернавского и Н.М.Чернавской [11,12] сделана оценка вероятности случайного формирования нового биологически значимого белка с учетом того, что в белке есть активный центр, в котором замены аминокислот практически недопустимы, и участки, свойства которых не сильно меняются при многих аминокислотных заменах. Там же сделана оценка количества возникающей в геноме информации при появлении нового белка. Полученная оценка указывает на то, что случайное формирование белка было вполне вероятно в процессе эволюции.
Интересна, хотя, по-видимому, не бесспорна, модель блочно-иерархического эволюционного отбора [13,14], согласно которой новые генетические тексты большой длины сначала случайно составляются из коротких текстов, оптимизированных в предыдущие эволюционные эпохи, а после составления оптимизируются. Модель блочно-иерархического эволюционного отбора критически проанализирована в [15].
Блочно-модульный принцип организации и эволюции молекулярно-генетических систем управления обосновывается В.А.Ратнером [16]. Согласно этому принципу эволюция генов, РНК, белков, геномов и молекулярных систем управления на их основе шла путем комбинирования блоков (модулей) снизу доверху, причем модулями, из которых составлялись вновь возникающие молекулярно-генетические системы, служили уже функционирующие макромолекулярные компоненты. По сравнению с моделью блочно-иерархического отбора блочно-модульный принцип более гибок и более реалистичен.
В модели "генов-мутаторов" [17] предполагается, что уровень мутаций может меняться и наследоваться, в результате чего при попадании популяции в новую среду, когда выгоден активный поиск новых свойств, уровень мутаций возрастает, а при длительном нахождении в постоянной среде, где важно сохранение уже найденных свойств, интенсивность мутаций падает.
Интересно проанализировать, как могли возникать достаточно нетривиальные системы обработки информации. Для простейших организмов (вирусов и бактерий) в качестве таковых можно рассматривать регулирование синтеза белков (функциональных и структурных элементов организма) в соответствии с определенной "программой". Например, в процессе онтогенеза фага Т4 происходит образование сложной пространственной структуры, в формировании которой участвуют несколько десятков белков, синтезируемых в соответствии с программой, закодированной геномом фага [18]. Иллюстративная модель эволюционного возникновения подобных "программ жизнедеятельности" предложена [19]. Согласно модели в процессе эволюционного формирования этих программ в генотип закладывается некоторая избыточность, которая приводит к тому, что при небольшой модификации генома часть блоков программ сохраняется. При введении в модель представлений о "генах-мутаторах" наблюдалось поведение, качественно сходное с явлением каскадного мутагенеза [20] – резким возрастанием интенсивности мутаций после дестабилизации генома.
В чрезвычайно интересном цикле работ С.Кауффмана с сотрудниками [21,22] исследуется эволюция автоматов, состоящих из соединенных между собой логических элементов. Отдельный автомат можно рассматривать как модель молекулярно-генетической системы управления живой клетки, при этом каждый логический элемент интерпретируется как регулятор синтеза определенного фермента. Модели Кауффмана позволяют сделать ряд предсказаний относительно "программ" жизнедеятельности клеток. В частности, продемонстрировано, что для одновременного обеспечения устойчивости и гибкости программы число входов логических элементов должно быть ограничено определенным интервалом, а именно составлять величину примерно равную 2-3. Для моделей Кауффмана разработаны эффективные методы анализа на базе статистической физики, эти модели получили широкую известность и исследовались рядом ученых. Подробнее основные результаты этой модели мы обсудим в следующей лекции.
1) Диплоидный организм: особь, имеющая два набора хромосом в каждой из ее клеток.
2) Аллель: Одна из различных форм гена, который может быть в заданном локусе.
Читайте также: