
Нейронные механизмы памяти кратко
Обновлено: 28.06.2024
Накопление, хранение и обработка информации — это важнейшее свойство нервных сетей. Невозможно переоценить биологическое значение этих процессов для адаптации поведения живого организма к окружающей среде. Без способности к научению и памяти ни отдельная особь, ни вид в целом не могли бы выжить, поскольку в этом случае было бы невозможно планировать свое поведение и преднамеренно избегать ошибок.
БЛАГАЯ ЗАБЫВЧИВОСТЬ
Совершенно ясно, что в нашей памяти откладывается лишь незначительная часть воспринимаемых нами явлений. В этой связи возникают вопросы: каким же образом мозг запоминает, что он запоминает и что забывает.
Без отбора информации и удаления ее из памяти мы были бы буквально затоплены потоком непрерывно поступающих данных. В настоящее время за единицу информации принимается бит. Для хранения 1 бита требуется 10 нейронов. Величина общей информационной емкости мозга равна
3 х 108 бит. Такой емкости хватило бы, для того чтобы хранить около 1% от общего потока информации, протекающей через наше сознание. Если считать, что в среднем информационный поток составляет 20 бит/сек, то за 70 лет при длительности активного дня 16 часов, общее поступление информации составит 3 х 1010 бит. Это в 100 раз больше, чем информационная емкость мозга.
Память есть результат образования новых условных связей, формирующихся в коре головного мозга, с помощью которых фиксируется индивидуальный опыт организма. Например, ребенок, единожды получив ожог от пламени свечи, от кипятка, на всю жизнь запоминает эти ощущения.
Память как процесс связана с восприятием информации, с ее хранением и использованием. Извлечение из хранилища памяти прошлого опыта называется актуализацией памяти.
ВИДЫ ПАМЯТИ
Различают несколько видов памяти.
Моторная память
Эволюционно это самый древний вид. В его основе лежит способность запоминать и производить какую-то программу движений. Этот вид памяти генетически запрограммирован. Например, ходьба, подъем по ступенькам, плаванье и т.д.
Эмоциональная память
Связана с фиксацией ощущений, которыми сопровождались те или иные события, явления жизни. Эмоции — эволюционно более древнее явление, чем ощущения. Эмоции выполняют регуляторную функцию в обеспечении поведения и адаптации организма к окружающей среде. Биологический смысл эмоциональной памяти заключается в том, что во всем диапазоне эмоциональных проявлений чаще всего фиксируются отрицательные эмоции, то есть вырабатывается система предупреждения.
Помимо этого эмоционально окрашенные ощущения фиксируются практически мгновенно и непроизвольно. Эмоциональная память самая прочная, поэтому имеет очень большое значение в процессе обучения.
Образная память
Связана с работой сенсорных систем или органов чувств. Информация запоминается в виде образов определенной модальности. Существуют зрительная, слуховая, тактильная, вкусовая и другие виды образной памяти. Эта память спонтанна, гибка и обеспечивает длительное хранение следа памяти.
Логическая память
Этот вид памяти эволюционно самый новый. Логическая память формируется только на базе второй сигнальной системы в процессе обучения. Вторая сигнальная система, по определению физиолога И.П. Павлова, — это слово. Сложно запомнить материал без понимания, без логического восприятия. Никаких природных готовых программ у этой памяти нет. Логическая память — результат тех интеллектуальных возможностей, которые есть у ребенка. Логическая память у младших и старших школьников различна. Лучше выражена она у старшеклассников.
В ЗАВИСИМОСТИ ОТ ВРЕМЕНИ
СИНАПТИЧЕСКИЙ ЭФФФЕКТ И ДРУГИЕ
ПОИСК НЕ ЗАКОНЧЕН
В онтогенезе в процессе дифференцировки во всех типах клеток, кроме нервных, реализуется от 1 до 12% генетической информации. Это специфическая генетическая информация о тех белках, которые должны синтезироваться в клетках определенного типа. В процессе онтогенеза разные типы клеток начинают использовать разную генетическую информацию.
В системе генома нервных клеток, особенно клеток структур больших полушарий, в зависимости от условий жизнедеятельности, меняется уровень синтеза белков, которые обеспечивают фиксацию жизненного опыта. Нервные клетки реализуют от 15 до 37% генетической информации. Если тренировать память, функциональные способности нервной деятельности повышаются до верхней границы.
В последнее время в самых различных структурах мозга были обнаружены нейроактивные пептиды. Предполагают, что они причастны к процессам памяти. Существует весьма генерализованное влияние пептидов, участвующих в общей активации и мотивационных механизмах.
Например, введение адренокортикотропного гормона или его фрагментов приводит к активации нейронов во многих отделах нервной системы. Ухудшение памяти связывают также с генетическим дефицитом вазопрессина. Окситоцин оказывает противоположное действие. Эндорфины и энкефалины регулируют память посредством взаимодействия с медиатором и уже через них оказывают влияние на метаболизм макромолекул. Нейропептиды могут либо усилить, либо ослабить действие медиатора.
Все ныне существующие представления и гипотезы о нейрофизиологических основах памяти не являются до конца изученными и доказанными. В этой связи и на сегодняшний день эта проблема интригующе интересна как для физиологов, так и для психологов.
С развитием микроэлектродной техники появилась возможность изучения электрофизиологических процессов, лежащих в основе памяти на уровне нервной клетки.
Наиболее эффективным оказался метод внутриклеточного отведения электрической активности отдельного нейрона. С его помощью можно анализировать роль синаптических процессов в изменении активности нейрона. В частности, на этой основе были установлены нейронные механизмы простой формы обучения — привыкания
Изучение нейронных основ памяти сопряжено с поиском структур, нейроны которых обнаруживают пластические изменения при обучении. Экспериментальным путем такие нейроны обнаружены у животных в гиппокампе, ретикулярной формации и некоторых зонах коры.
По его представлениям Е. Н. Соколова, информация закодирована в нейронных структурах мозга в виде особых векторов памяти, которые создаются набором постсинаптических локусов на теле нейрона-детектора, имеющих разную электрическую проводимость. Этот вектор определяется как единица структурного кода памяти. Вектор восприятия состоит из набора постсинаптических потенциалов разнообразной амплитуды. Размерности всех векторов восприятия и всех векторов памяти одинаковы. Если узор потенциалов полностью совпадает с узором проводимостей, то это соответствует идентификации воспринимаемого сигнала.
Психофизиологический подход к проблеме сознания.
В психофизиологии сознание – особое состояние мозга, при котором только и возможна реализация высших психических функций.
Х. Дельгадо представлял сознание как организованную группу процессов в нервной ткани, возникающих на предшествующие интрапсихические (вызванные внутренними причинами) или экстрапсихические (вызванные внешними причинами) процессы. т.е. сознание, воспринимает, классифицирует, трансформирует и координирует вызвавшие его процессы с целью начать действие на основе предвидения его последствий и в зависимости от наличной информации.
В других определениях подчеркиваются системность сознания, комплексность выполняемых им функций, связь с памятью (прошлым и будущим человека), привязанность к мозговому субстрату. П.В. Симонов (1987), например, особо выделяет коммуникативный аспект сознания, определяя его как оперирование знанием, способность к направленной передаче информации от одного лица к другому.
Сознание и уровни бодрствования.
В качестве самостоятельной характеристики предлагается выделять содержание сознания.
Существует 2 уровня внимания:
1. Зона ясного виденья.
2. Зона смутного виденья
В зависимости от характера деятельности зона повышенной активации может перемещаться по коре больших полушарий.
Структуры мозга, участвующие в процессе восприятия - это
1.Корковые проекции органов чувств.
2. Приток активирующих воздействий из РФ
3. Связь лимбической системы с верховным центром регуляторных функций (бессознательные рефлекторные реакции)
4. Связь эндокринной гормональной системы (бессознательные рефлекторные реакции)
Поводом написания данной статьи послужила публикация материала американских неврологов на тему измерения емкости памяти головного мозга человека, и представленная на GeekTimes днем ранее.
В подготовленном материале постараюсь объяснить механизмы, особенности, функциональность, структурные взаимодействия и особенности в работе памяти. Так же, почему нельзя проводить аналогии с компьютерами в работе мозга и вести исчисления в единицах измерения машинного языка. В статье используются материалы взятые из трудов людей, посвятившим жизнь не легкому труду в изучении цитоархитектоники и морфогенетике, подтвержденный на практике и имеющие результаты в доказательной медицине. В частности используются данные Савельева С.В. учёного, эволюциониста, палеоневролога, доктора биологических наук, профессора, заведующего лабораторией развития нервной системы Института морфологии человека РАН.
Прежде, чем преступить к рассмотрению вопроса и проблемы в целом, мы сформулируем базовые представления о мозге и сделаем ряд пояснений, позволяющих в полной мере оценить представленную точку зрения.
Первое что вы должны знать: мозг человека — самый изменчивый орган, он различается у мужчин и женщин, расовому признаку и этническим группам, изменчивость носит как количественный (масса мозга) так и качественный (организация борозд и извилин) характер, в различных вариациях эта разница оказывается более чем двукратной.
Второе: мозг самый энергозатратный орган в человеческом организме. При весе 1/50 от массы тела он потребляет 9% энергии всего организма в спокойном состоянии, например, когда вы лежите на диване и 25% энергии всего организма, когда вы активно начинаете думать, огромные затраты.
Третье: в силу большой энергозатраты мозг хитер и избирателен, любой энергозависимый процесс невыгоден организму, это значит, что без крайней биологической необходимости такой процесс поддерживаться не будет и мозг любыми способами старается экономить ресурсы организма.
Вот, пожалуй, три основных момента из далеко не полного списка особенностей мозга, которые понадобится при анализе механизмов и процессов памяти человека.
Что же такое память? Память – это функция нервных клеток. У памяти нет отдельной, пассивной эноргонезатратной локализации, что является излюбленной темой физиологов и психологов, сторонников идеи нематериальных форм памяти, что опровергается печальным опытом клинической смерти, когда мозг перестает получать необходимое кровоснабжение и примерно через 6 минут после клинической смерти начинаются необратимые процессы и безвозвратно исчезают воспоминания. Если бы у памяти был энергонезависимый источник она могла бы восстановиться, но этого не происходит, что означает динамичность памяти и постоянные энергозатраты на ее поддержание.
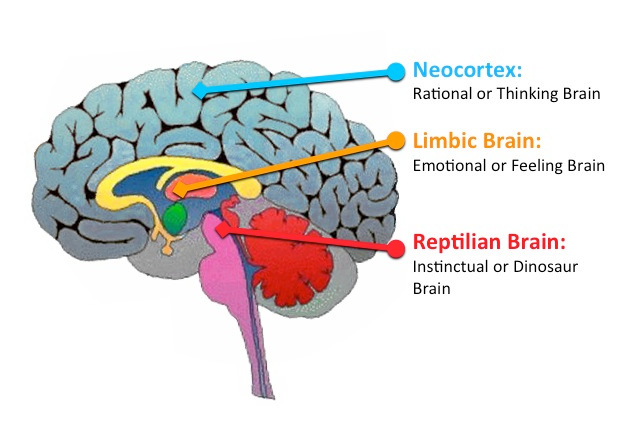
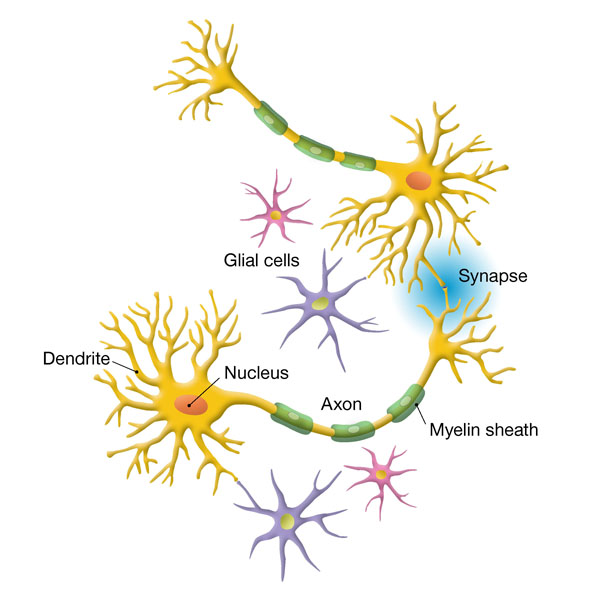
Важно знать, что нейроны, определяющие память человека, находятся преимущественно в неокортоксе. Неокортекс содержит порядка 11млрд. нейронов и в разы больше глии. (Глия – тип клеток нервной системы. Глия является средой для нейронов глиальные клетки служат опорным и защитным аппаратом для нейронов. Метаболизм глиальных клеток тесно связан с метаболизмом нейронов, которые они окружают.
Глии, связи нейронов:
Хорошо известно, что в памяти информация хранится разное время, существуют такие понятия как долговременная и кратковременная память. События и явления быстро забываются, если не обновляются и не повторяются, что очередное подтверждение динамичности памяти. Информация определенным образом удерживается, но в отсутствии востребованности исчезает.
Как говорилось ранее, память – энергозависимый процесс. Нет энергии – нет памяти. Следствием энергозависимости памяти является нестабильность ее содержательной части. Воспоминания о прошедших событиях фальсифицируются во времени вплоть до полной неадекватности. Счета времени у памяти нет, но его заменяет скорость забывания. Память о любом событии уменьшается обратно пропорционально времени. Через час забывается ½ от всего попавшего в память, через сутки – 2/3, через месяц – 4/5.
Память мозга – вынужденная компенсаторная реакция нервной системы. Любая информация переходит во временное хранение. Поддержка стабильности кратковременной памяти и восприятия сигналов от внешнего энергетически крайне затратна, к тем же клеткам приходят новые возбуждающие сигналы и, накапливаются ошибки передачи и происходит перерасход энергетических ресурсов. Однако ситуация не так плоха, как выглядит. Нервная система обладает долговременной памятью. Зачастую она так трансформирует реальность, что делает исходные объекты неузнаваемыми. Степень модификации хранимого в памяти объекта зависит от времени хранения. Память сохраняет воспоминания, но изменяет их так, как хочется обладателю. В основе долговременной памяти лежат простые и случайные процессы. Дело в том, что нейроны всю жизнь формируют и разрушают свои связи. Синапсы постоянно образуются и исчезают. Довольно приблизительные данные говорят о том, что этот процесс спонтанного образования одного нейронного синапса может происходить у млекопитающих примерно 3-4 раза в 2-5 дней. Несколько реже происходит ветвление коллатералей, содержащих сотни различных синапсов. Новая полисинаптическая коллатераль формируется за 40-45 дней. Поскольку эти процессы происходят в каждом нейроне, вполне можно оценить ежедневную емкость долговременной памяти для любого из животных. Можно ожидать, что в коре мозга человека ежедневно будет образовываться около 800 млн. новых связей между клетками и примерно столько же будет разрушено. Долговременным запоминанием является включение в новообразованную сеть участков с совершенно не использованными, новообразованными контактами между клетками. Чем больше новых синаптических контактов участвует в сети первичной (кратковременной) памяти, тем больше у этой сети шансов сохраниться надолго.
Из выше сказанного ясно, что мозг динамическая структура, постоянно перестраивается и имеет определенные физиологические пределы, так же мозг чрезмерно энергозатратный орган. Мозг не физиологичен, а морфогенетичен, потому его активности некорректно и неправильно измерять в системах, используемых и применимых в информационных технологиях. Из за индивидуальной изменчивости мозга не представляется возможным делать какие либо выводы обобщающие различные функциональные показатели мозга человека. Математические методы так же не применимы в расчете структурного взаимодействия в работе мозга человека, из за постоянного изменения, взаимодействия и перестраивания нервных клеток и связей между ними, что в свою очередь доводит до абсурда работу американских ученых в исследовании емкости памяти головного мозга человека.


В этой работе соединились две большие области исследований: механизмы памяти и амилоиды. Предыстория открытия интересна и содержит немало важных деталей.
Амилоиды (см. amyloid) известны в первую очередь как патологические белковые структуры, вызывающие обширную группу амилоидных заболеваний, в частности болезни Альцгеймера и Паркинсона, а также прионные болезни. Они представляют собой фибриллярные полимеры некоторых в норме растворимых клеточных белков. Амилоиды катализируют структурную перестройку и присоединение к себе мономеров того же белка, и за счет этого растут. Они намного прочнее штатных клеточных полимеров, составляющих цитоскелет, поскольку фактически представляют собой единый мультимолекулярный бета-слой, в котором отдельные молекулы соединены множеством водородных связей. Поэтому амилоиды обладают высокой устойчивостью к протеазам и накапливаются, вызывая болезнь.
Амилоиды могут быть инфекционными, и тогда они называются прионами. У человека и животных прионы связаны лишь с одним белком, PrP, и вызывают коровье бешенство, скрейпи овец и болезнь Крейцфельдта–Якоба у людей. Инфекционность прионов связана с общим свойством амилоидов катализировать структурную перестройку. Различие же с прочими амилоидами по инфекционности определяется частными деталями: расположением прионного белка PrP на внешней клеточной мембране и, вероятно, какими-то механизмами, дробящими полимеры PrP на множество мелких, более подвижных частиц.
Явление, аналогичное прионам, было обнаружено у дрожжей Saccharomyces cerevisiae. У дрожжей, однако, прионы проявляются не как болезнь, а как фенотипы с нестандартным, неменделевским способом наследования. Прионы дрожжей (см. fungal prions) возникают спонтанно, но достаточно редко. Затем они могут стабильно сохраняться в ряду поколений, а при скрещивании и мейотической сегрегации передаваться всем потомкам. Таким образом, прионогенный белок может стабильно находиться в двух состояниях: прионном (полимеризующемся) или нормальном. А значит, клетку с прионогенным белком можно представить, как однобитную ячейку памяти.
А теперь перейдем к устройству памяти. Большая доля знания о работе нейронов была получена при изучении нейронов моллюска Aplysia californica. Этот крупный моллюск имеет большие и удобные для изучения нейроны. Ключевым механизмом памяти считается способность нейронов изменять силу своих синапсов, или синаптическая пластичность. Существует два вида памяти — кратковременная и долговременная. Кратковременная память опосредуется фосфорилированием некоторых уже существующих белков и укреплением имеющихся синаптических связей. Долговременная требует синтеза новых мРНК и белков и часто сопровождается установлением новых синаптических связей. Поскольку синтез мРНК происходит в ядре и затрагивает весь нейрон, возник вопрос: происходит ли событие запоминания одновременно во всех синапсах нейрона или же оно специфично для каждого синапса? На модели изолированного нейрона было показано, что единичный импульс нейромедиатора серотонина вызывает кратковременное синапс-специфическое запоминание, а два и более — долговременное. При этом, если первый импульс прикладывали к одному синапсу, а второй — к другому, то долговременное запоминание происходило во втором синапсе и только в нём.
И действительно, оказалось, что CPEB необходим для долговременной, но не кратковременной памяти, и его синтез намного увеличивается при стимуляции нейрона нейромедиатором серотонином. Далее обнаружилась удивительная вещь: по своей структуре CPEB оказался похож на дрожжевые прионные белки. Эти белки довольно несхожи между собой, но каждый из них имеет две части: функциональный домен и прионный домен, способный полимеризоваться. Функциональные домены совершенно различны, а прионные обладают общим свойством: они не структурированы и сильно обогащены аминокислотными остатками глутамином и аспарагином. Это свойство позволяет прионным доменам полимеризоваться в амилоидные фибриллы, и именно такой домен был обнаружен в белке CPEB. Свойства CPEB проверили в дрожжевой модели, и оказалось, что он ведет себя, как полноценный дрожжевой прион, то есть может переходить в стабильно наследуемое полимерное состояние. Правда, в отличие от дрожжевых белков, у которых прионное состояние функционально неактивно, у CPEB прионное состояние отличалось повышенной активностью. Всё это позволило предположить, что переход CPEB в полимерное состояние является ключевым событием в формировании долговременной памяти.
Однако доказательство этого тезиса оказалось нелегким, и, несмотря на интенсивные усилия, следующее продвижение в этой теме произошло лишь через семь лет. Видимо, это говорит о том, насколько сложнее манипулировать нервными клетками аплизии в сравнении с клетками дрожжей: трудно набрать достаточное количество клеток для биохимического анализа, сложнее манипуляции с генами. На этом этапе в тело нейрона аплизии инъецировали гены, кодирующие различные гибриды белка CPEB, сшитые с зеленым флуоресцентным белком. Такие гибриды — классический инструмент в изучении прионов: если белок растворим, клетка светится равномерно, а если он перешел в амилоидное состояние — свечение концентрируется в яркие точки. Гибридные белки, синтезированные нейроном, образовали характерные зеленые точки, а контрольный белок без глутамин-богатого домена давал диффузное свечение. Амилоидное состояние СРЕВ-GFP в точках было подтверждено окрашиванием амилоид-специфичным флуоресцентным красителем тиофлавином S. Также было показано, что переход СРЕВ-GFP в амилоидное состояние усиливался при стимуляции нервных клеток нейромедиатором серотонином. Прогресс небольшой, да и получен он был при искусственно завышенном уровне синтеза СРЕВ.

Рис. 2. Структура кодирующей Orb2 мРНК и двух белков Orb. Изображение из обсуждаемой статьи Amitabha Majumdar et al. в Cell, с изменениями
Но недавняя работа, сделанная уже на дрозофиле, расставила все точки над i. В отличие от аплизии, у дрозофилы есть два варианта белка СРЕВ: Orb2A и Orb2B, которые получаются из одной мРНК в результате альтернативного сплайсинга. Orb2B синтезируется постоянно (конститутивно), а Orb2A — лишь в ответ на стимуляцию нейрона. Они одинаковы в карбокси-концевой части, содержащей глутамин-богатый прионо-подобный домен и РНК-связывающий домен, но отличаются в амино-концевой части, имеющей 8 аминокислот в Orb2A и 162 аминокислоты в Orb2B (рис. 2). Было показано, что оба белка способны полимеризоваться, однако Orb2B может стабильно пребывать в растворимом состоянии, а Orb2A, напротив, с высокой вероятностью начинает полимеризацию. В частности, это наблюдали при флуоресцентной микроскопии гибридных белков Orb2-GFP (рис. 3). Оказалось, что способность Orb2A начинать полимеризацию связана с его уникальным N‑концевым фрагментом из 8 аминокислот.

Рис. 3. При экспрессии в нейроне Orb2A-GFP образует амилоидные полимеры, видные как яркие точки, а Orb2B-GFP не полимеризуется, давая диффузное свечение. Длина масштабной линейки 10 мкм. Изображение из обсуждаемой статьи Amitabha Majumdar et al. в Cell, с изменениями
Эти наблюдения позволили предложить простую схему: при стимуляции синапса синтезируется Orb2A, он переходит в полимерное состояние и увлекает за собой Orb2B. Далее процесс полимеризации поддерживается молекулами Orb2B и может продолжаться сколь угодно долго, что соответствует фиксации события в долговременной памяти.
Для подтверждения этой модели провели мутагенез Orb2A и получили мутации, нарушающие его способность инициировать полимеризацию. Половина мутаций попала в уникальный N‑концевой фрагмент Orb2A, содержащий лишь 8 аминокислот. Наиболее эффективная из этих мутаций была изучена подробнее. Это оказалась замена в пятой позиции фенилаланина на тирозин.
Важным достижением данной работы является и то, что она впервые показала, как клетка может управлять переходом в амилоидное состояние. Однако существование механизма, способного вывести синапс из этого состояния, представляется маловероятным. Скорее всего, в компьютерной терминологии, мозг является устройством с однократной записью, подобным CD, а не винчестеру.
Несомненно, эта работа устанавливает один из принципов устройства живых организмов. Однако она заслуживает не только похвал. На всякого мудреца довольно простоты, и в этой работе есть существенный прокол. Он не отменяет главного вывода, хотя и ослабляет его и бросает плотную тень на научно-издательский процесс. Мы привыкли думать, что наиболее авторитетные научные журналы, к которым относится Cell, не могут публиковать откровенных ошибок. Ан нет. Среди представленных доказательств существенную роль имеет подтверждение существования полимеров СРЕВ методом электрофореза, выполненным совершенно некорректно. Авторы тщательно кипятят образцы в присутствии детергента, а в этих условиях любой амилоид должен раствориться. Удивительным образом растворяется не совсем всё, но всё равно такое безграмотное и неколичественное использование столь сильных средств недопустимо. В частной переписке авторы признали этот просчет. А ведь двумя годами раньше у них была еще одна статья в Cell с той же ошибкой. И это один из лучших и самых строгих журналов!
Как бы то ни было, но теперь мы понимаем основу устройства памяти. Интересно, для чего еще могут быть использованы амилоиды? Есть мнение, что это может быть процесс клеточной дифференциации.
Есть способы классификации памяти, основанные на ее функциональном значении. Оперативная память связана с хранением информации, необходимой для решения конкретной задачи, в течение времени, которое требуется для ее решения. Тем не менее она отличается от кратковременной памяти, поскольку к оперативной памяти относятся только те следы, которые были воспроизведены, тогда как в кратковременной памяти хранится и воспроизведенная, и невоспроизведенная информация. В оперативной памяти характерно сочетание сиюминутного сознания и мгновенного воспоминания о накопленной ранее информации (Гольдман-Ракич, 1992). Она имеет дело с фактами и цифрами и обеспечивает срочную активацию и кратковременное хранение символической информации, а также возможность
Примером оперативной памяти может служить операция переноса при вычислении в уме, требующая временного запоминания ряда чисел и удержания результата одного сложения, пока подсчитывается вторая сумма. Другими примерами могут быть планирование шахматных ходов, построение предложения в разговорной речи. Кратковременная память имеет решающее значение в процессе продукции и понимания речи, при обучении и мышлении. Информация из оперативной может стать долговременной, если позднее будет прочно зафиксирована, или может исчезнуть, не оставив следа.
Дети, не достигшие восьмимесячного возраста, у которых нейрональная организация коры еще не имеет характеристик, свойственных взрослому человеку, плохо справляются с такими заданиями и продолжают выбирать ту коробку, на которую указывали раньше, но в которой уже нет игрушки. Так же плохо справляются с ними обезьяны, у которых хирургически удаляли пре-фронтальную кору. Оперированные обезьяны просто повторяли предыдущий ответ, не меняя его в связи с вновь поступившей информацией.
Ж.П.Буржуа и П. Гольдман-Ракич изучали скорость, с которой формируются связи в префронтальной коре детенышей обезьян. Они обнаружили, что быстрее всего образуются синапсы в префронтальной коре в возрасте двух-четырех месяцев, когда у обезьян появляется способность к выполнению тестов с отсроченным воспроизведением (Гольдман-Ракич, 1992).
Активность отдельных нейронов в префронтальной коре подробно изучена Дж. Фастером, К. Кубота и X. Ники. Обезьянам вводили в префронтальную область коры тонкие электроды, регистрирующие нейронную активность, и обучали выполнять простые тесты с отсроченным ответом. Выявилась четкая специфичность в ответах нейронов. Электрическая активность одних клеток усиливалась при предъявлении информации, других — только во время отсрочки, когда животные ее припоминали. Нейроны третьей группы реагировали тогда, когда начиналась двигательная реакция (Гольдман-Ракич, 1992).
Специфическая активность нейронов в процессе отсрочки показана в эксперименте, где регистрировали электрическую активность отдельных нейронов префронтальной коры в тесте с отсроченным ответом. Предварительно обезьяну обучали фиксировать взор на маленьком пятне в центре телевизионного экрана. Затем в одном из восьми участков экрана на короткое время появлялся раздражитель (обычно это небольшой квадрат). В конце отсрочки продолжительностью от трех до шести секунд центральное пятно, на котором фиксировался взор, выключалось, что служило животному сигналом о необходимости перевести взгляд в ту точку экрана, где перед отсрочкой появлялся раздражитель. В случае правильного ответа животное получало сок. Поскольку взор животного был фиксирован на центральном пятне, каждый из зрительных раздражителей активировал определенную группу клеток сетчатки. Эти клетки приводили в действие лишь определенные от- " делы зрительных проводящих путей (рис. 15.1) (Гольдман-Ракич, 1992).
С помощью этого опыта удалось показать, что некоторые нейроны префронтальной коры обладают своего рода полями памяти: когда из поля зрения животного исчезает цель, имеющая для него особую значимость, некоторые нейроны префронтальной коры начинают генерировать электрические сигналы вдвое чаще. Они остаются возбужденными до конца отсрочки, т. е. до того момента, когда животное переводит взгляд в нужную точку экрана. Каждый нейрон кодирует своей активностью определенное положение зрительного раздражителя.
Нейроны, способные хранить информацию о зрительном и пространственном расположении сигнала (отвечающие за пространственную кратковременную память), по-видимому, организованы в префронтальной коре в специфическую область. Если во время отсрочки их работа нарушается, возможны ошибки при воспоминании.
Поскольку префронтальная кора функционирует как посредник между памятью и действием, можно предположить, что повреждение этой области мозга не наносит ущерба знаниям животного о внешнем мире, но нарушает его способность осознавать эти знания и пользоваться ими. Действительно, обезьяны с поврежденной префронтальной корой, как и многие люди с аналогичными травмами, без труда обучаются выполнять тесты на
Рис. 15.1. Тест с отсроченным ответом для изучения оперативной памяти у обезьян. Когда обезьяна фиксирует взор на центральном пятне, на экране (слева) вспыхивает и затем исчезает цель. Во время длящейся несколько секунд отсрочки обезьяна хранит об этой цели “мысленную” память (в центре). Когда центральное пятно исчезает, животное переводит взгляд туда, где появлялась цель (справа). Как показывает регистрация электрической активности, некоторые нейроны префронтальной коры реагируют на появление цели, другие сохраняют о ней “мысленную” память, а третьи разряжаются, подготавливая двигательный ответ (Гольдман-Ракич, 1992).
сенсорное различение. Все формы ассоциативной, долговременной, памяти сохраняются до тех пор, пока испытуемый воспринимает знакомые внешние раздражители, ассоциированные с определенными последствиями и ожиданиями (Гольдман-Ракич, 1992).
Префронталъная кора является частью сети реципрокных связей между центральной бороздой и основными сенсорными, лимбическими и премоторны-ми зонами коры. Эта сеть отвечает за переработку пространственной информации. Нейроны в центральной борозде чувствительны также к специфическому расположению зрительных раздражителей, а значит, имеют доступ к зрительной и пространственной информации, перерабатываемой в других отделах мозга. Центральная борозда получает сигналы из задней теменной коры, где идет анализ информации, связанной с пространственным зрением.
Кратковременная память зависит от доступности информации, хранящейся в долговременной памяти. Эффективность долговременной памяти определяется сохранностью гиппокампа. Метод авторадиографии выявил, что гиппокамп и область центральной борозды часто активируются одновременно во время выполнения тестов с отсроченным ответом. П.С. Гольдман-Ракич полагает, что главная функция гиппокампа связана с консолидацией следов памяти, а префронтальной коры — с извлечением результатов такого обучения из долговременного хранилища в других отделах мозга и их непосредственным включением в оперативную деятельность.
Однако функционирование рабочей памяти зависит и от других структур мозга. Запись электрической активности в процессе запоминания у человека обнаружила появление медленной позитивной волны в лобно-цен-тральных областях на слова, которые впоследствии были узнаны (Friedman, 1990). Существует достаточно данных о том, что эффективность мнемичес-кой деятельности связана с развитием таламокортикальных сетей и частотой альфа-ритма (Khmesh e. a , 1990).
В префронтальной коре выявлено высокое содержание катехоламинов, прежде всего дофамина, регулирующего клеточную активность, связанную с кратковременной памятью. Нейроны некоторых слоев мозговой коры богаты определенными рецепторами дофамина — D1. Именно эти нейроны проецируются в таламус, где происходит переключение потока информации в кору.
Префронтальная кора и области, с которыми она активно связана (гиппо-камп, нижняя часть теменной коры, таламус), характеризуются высокой метаболической активностью при выполнении теста с отсроченным воспроизведением. Метаболическая активность мозга имеет разный характер — в зависимости оттого, требует ли задание от испытуемого памяти о местоположении предметов или об их признаках. При необходимости запоминать ряд слов или произносить глагол, ассоциативно связанный с высвеченным на экране существительным, отмечалось усиление работы нейронов префронтальной коры.
П.С. Гольдман-Ракич полагает, что префронтальная кора разделена на множество участков памяти, каждый из которых специализируется на кодировании особого рода информации, например, о местоположении, отдельных признаках объектов, таких как цвет, размеры, форма, а у людей — еще и семантических и математических знаний. Эти нейроны также выполняют функцию возбуждения или торможения других структур мозга. Кле1 -ки каждого слоя коры образуют в мозге строго специфические системы связей (рис. 15.2) Клетки одного типа, находящиеся в пятом слое коры, проецируются в хвостатое ядро, скорлупу и верхние бугры четверохолмия. Нейроны шестого слоя проецируются в таламус, через который к коре проходят сенсорные сигналы с периферии мозга
Префронтальная кора не может самостоятельно инициировать двигатель-!!
ные ответы, однако она контролирует движение, программируя, облегча^
или подавляя команды тем мозговым структурам, которые управляют сокра.д
щением или расслаблением мышц. , ^
Читайте также: