
Межклеточные контакты гистология кратко
Обновлено: 06.07.2024
Плазмолемма многоклеточных животных организмов принимает активное участие в образовании специальных структур — межклеточных соединений (junctiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 7).
Простое межклеточное соединение, (junctio intercellularis simplex) — сближение плазмолемм соседних клеток на расстояние 15—20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток. Разновидностью простого соединения является "пальцевидное", или соединение по типу замка.
Плотное соединение (запирающая зона) (zonula occludens) — зона, где слои двух плазмолемм максимально сближены, здесь происходит как бы слияние участков плазмолемм двух соседних клеток. Роль плотного замыкающего соединения заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды.
Часто встречается, особенно в эпителии, особый тип соединения — пятно сцепления, или десмосома (desmosoma). Эта структура представляет собой небольшую площадку, иногда имеющую слоистый вид, диаметром до 0,5 мкм, где между мембранами располагается зона с высокой электронной плотностью. К плазмолемме в зоне десмосомы со стороны цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под этим утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Функциональная роль десмосом заключается главным образом в механической связи между клетками.
Щелевидное соединение, или нексус (nexus), представляет собой область протяженностью 0,5—3 мкм, где плазмолеммы разделены промежутком в 2—3 нм. Со стороны цитоплазмы никаких специальных примембранных структур в данной области не обнаруживается, но в структуре плазмолемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют как бы каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевидного соединения заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус.
Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы). Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому.
Строение синапса также рассматривается в теме нервные окончания.
Плазмолемма многоклеточных животных организмов принимает активное участие в образовании специальных структур — межклеточных соединений (junctiones intercellulares), обеспечивающих межклеточные взаимодействия. Различают несколько типов таких структур (рис. 7).
Простое межклеточное соединение, (junctio intercellularis simplex) — сближение плазмолемм соседних клеток на расстояние 15—20 нм. При этом происходит взаимодействие слоев гликокаликса соседних клеток. Разновидностью простого соединения является "пальцевидное", или соединение по типу замка.
Плотное соединение (запирающая зона) (zonula occludens) — зона, где слои двух плазмолемм максимально сближены, здесь происходит как бы слияние участков плазмолемм двух соседних клеток. Роль плотного замыкающего соединения заключается в механическом соединении клеток друг с другом. Эта область непроницаема для макромолекул и ионов и, следовательно, она запирает, отграничивает межклеточные щели (и вместе с ними собственно внутреннюю среду организма) от внешней среды.
Часто встречается, особенно в эпителии, особый тип соединения — пятно сцепления, или десмосома (desmosoma). Эта структура представляет собой небольшую площадку, иногда имеющую слоистый вид, диаметром до 0,5 мкм, где между мембранами располагается зона с высокой электронной плотностью. К плазмолемме в зоне десмосомы со стороны цитоплазмы прилегает участок электронноплотного вещества, так что внутренний слой мембраны кажется утолщенным. Под этим утолщением находится область тонких фибрилл, которые могут быть погружены в относительно плотный матрикс. Функциональная роль десмосом заключается главным образом в механической связи между клетками.

Щелевидное соединение, или нексус (nexus), представляет собой область протяженностью 0,5—3 мкм, где плазмолеммы разделены промежутком в 2—3 нм. Со стороны цитоплазмы никаких специальных примембранных структур в данной области не обнаруживается, но в структуре плазмолемм соседних клеток друг против друга располагаются специальные белковые комплексы (коннексоны), которые образуют как бы каналы из одной клетки в другую. Этот тип соединения встречается во всех группах тканей. Функциональная роль щелевидного соединения заключается в переносе ионов и мелких молекул от клетки к клетке. Так, в сердечной мышце возбуждение, в основе которого лежит процесс изменения ионной проницаемости, передается от клетки к клетке через нексус.
Синаптические соединения, или синапсы (synapsis). Этот тип соединений характерен для нервной ткани и встречается в специализированных участках контакта как между двумя нейронами, так и между нейроном и каким-либо иным элементом, входящим в состав рецептора или эффектора (например, нервно-мышечные, нервно-эпителиальные синапсы). Синапсы — участки контактов двух клеток, специализированных для односторонней передачи возбуждения или торможения от одного элемента к другому.
ЦИТОЛОГИЯ
МЕЖКЛЕТОЧНЫЕ КОНТАКТЫ
Простой контакт — соединение клеток за счет пальцевидных впячиваний и выпячиваний цитомембран соседних клеток. Специфических структур, формирующих контакт, нет.
Плотный замыкающий контакт — соприкасаются билипидные слои мембран соседних клеток. В области зоны плотных контактов между клетками не проходят практически никакие вещества.
Межклеточные адгезионные соединения:
Точечные — контакт образуется на небольшом по площади участке цитомембран соседних клеток.
- В области контакта в цитомембрану встроены специальные трансмембранные белки — кадгерины, которые соединяются с кадгеринами другой клетки.
- Для соединения кадгеринов нужны ионы кальция.
- Со стороны цитоплазмы к кадгеринам присоединяются белки ,бета-катенин, альфа-катенин, гамма-катенин, PP-120, EB-1, и к ним присоединяются актиновые микрофиламенты.
- Контакт образуется на небольшом по площади участке.
- В месте контакта в цитомембрану встроены трансмембранные белки десмоглеин и десмоколин, которые соединяются с такими же белками другой клетки.
- Для соединения десмоколинов и десмоглеинов нужны ионы кальция.
- Со стороны цитоплазмы к десмоколину и десмоглеину присоединяются промежуточные белки — десмоплакин и плактоглобин, к которыем присоединяются промежуточные филаменты.
Все типы адгезионных контактов, кроме механического скрепления клеток между собой и с внеклеточным матриксом, каким-то непонятным в настоящее время образом передают информацию о состоянии окружающей среды внутрь клетки (в цитоплазму и ядро). При этом ход различных биологических процессов в клетке может меняться.
- Контакт образуется на небольшом по площади участке.
- В месте контакта в цитомембрану встроены трансмембранные белки коннексины, которые соединяются между собой и образуют водный канал в толще мембраны — конексон.
- Коннексоны контактирующих клеток соединяются (или сопоставляются), в результате чего между соседними клетками образуется канал, с помощью которого из одной клетки в другую (в обоих направлениях) свободно проходит вода, малые молекулы и ионы, а также электрический ток.
Синапсы — см. раздел по нервной ткани. Обеспечивают передачу потенциала действия (нервного импульса) с нервной клетки на другую нервную или иную клетку.
- Адгезионные межклеточные контакты. Этот тип межклеточных контактов механически скрепляет клетки между собой. Связь клеток с внеклеточным матриксом при помощи подобных контактов рассмотрена в III А 3. К адгезионным относятся промежуточный контакт (опоясывающая десмосома, zonula adherens), десмосома (macula adherens), полудесмосома.
- Замыкающий контакт. Фактически это плотный контакт (zonula occludens).
- Проводящие (коммуникационные) контакты передают химические и электрические сигналы от клетки к клетке. К ним относятся щелевые контакты и синапсы (см. главу 8.1 III Г).
- Адгезионные контакты
- Структура. Мембраны соседних клеток разделены промежутком шириной 10-20 нм, заполненным аморфным или фибриллярным материалом. Электроноплотная пластинка на цитоплазматической стороне клеточной мембраны в пределах контакта содержит белки плакоглобин, винкулин, а-актинин и радицин. В пластинку вплетены концы актинсодержащих микрофиламентов. В образовании контакта участвуют трансмембранные белки адгезии из семейства кадгерина.
- Цепь молекулярных взаимодействий выглядит следующим образом: актин (примембранные микрофиламенты) — а-актинин — винкулин — Е-кадгерин (трансмембранный гликопротеин адгезии). Е-кадгерин в присутствии Ca2+ прочно связывает мембраны соседних клеток. Цепь молекулярных взаимодействий в промежуточном контакте, начиная с актина микрофиламентов и до винкулина включительно, аналогична таковой в точечном (фокальном) адгезионном контакте (III А 3, рис. 4-2). Если винкулин фокального контакта взаимодействует с трансмембранными бел- ками-рецепторами (интегрины), то в промежуточном контакте он связывается с Е-кадгерином.
- Функция. Промежуточный контакт скрепляет не только мембраны соседних клеток, но и стабилизирует их цитоскелет, объединяя клетки с их содержимым в единую жёсткую систему.
- Примеры
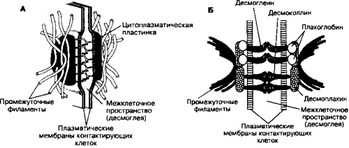
- Структура. Участки клеточных мембран, входящие в состав десмосомы, разделены слоем десмоглеи толщиной 20-30 нм. С внутренней стороны к плазматической мембране примыкает цитоплазматическая пластинка толщиной 10-40 нм с вплетёнными в неё промежуточными филаментами. В десмосомах всех клеточных типов присутствуют следующие белки: плакоглобин, десмоплакины, десмоглеины, десмоколлины.
- Цепь молекулярных взаимодействий (рис. 4-3). Молекулы образуют две линии связи:
- Функция. Десмосомы поддерживает структурную целостность ткани, скрепляя клетки между собой. Десмосомы в комплексе с промежуточными филаментами придают ткани упругость и поддерживают в ней усилие натяжения.
Рис. 4-3. Строение (А) и организация (Б) десмосомы. Плазматические мембраны клеток разделены промежутком 20-30 нм, в котором находятся внемембранные части Са2*-связывающихбелков десмоглеина и десмоколлина. К внутренней (цитоплазматической) поверхности плазматической мембраны прилегает цитоплазматическая пластинка с вплетёнными в неё промежуточными филаментами. В состав этой пластинки входят десмоплакины, плакоглобин и часть молекулы десмоглеина [из Alberts В et al, 1989, Gumbiner BM, 1993]
- Примеры. Десмосомы скрепляют клетки одного типа и различных типов.
- Патология. При аутоиммунном кожно-слизистом пемфигусе (паранеопластичес- кий процесс) в организме больного вырабатываются аутоантитела против десмоп- лакина I и пемфигоидного Ar (IV Б I в (I) (а)).
- Структура. Полудесмосома, как и десмосома, содержит цитоплазматическую пластинку с вплетёнными в неё промежуточными филаментами. Особенность состава цитоплазматической пластинки полудесмосомы — пемфигоидный антиген. Этого Ar нет в цитоплазматической пластинке десмосом.
- Примеры
- Плотный контакт формирует в слое клеток барьер проницаемости, разделяющий различные по химическому составу среды (например, внутреннюю и внешнюю). Барьер проницаемости, сформированный плотным контактом, задерживает даже малые молекулы.
- Каёмчатые клетки эпителия кишки (см. главу 12 IX Б I а (I) (б))
- Коммуникационные (проводящие) контакты. Контакты этого типа — щелевые и синапсы — передают сигналы от клетки к клетке.
- Структура. Плазматические мембраны клеток, образующих щелевой контакт, разделены щелью шириной 2~4 нм. Коннексон — трансмембранный белок цилиндрической конфигурации; состоит из 6 CE коннексина. Два коннексона соседних клеток соединяются в межмембранном пространстве и образуют канал между клетками (рис. 4-5). Канал коннексона диаметром 1,5 нм пропускает ионы и молекулы с Mr до 1,5 кД.
- Коннексин-43 экспрессируют кардиомиоциты, особенно важен при развитии миокарда.
- Коннексин-32 входит в состав щелевых контактов между гепатоцитами, найден в составе миелиновых волокон периферического нерва (см. главу 8.1 IV Б 4 в (5) (а)).
- Функция. Щелевой контакт контролирует проницаемость между взаимодействующими клетками. В некоторых клетках (например, глиальные) подобный механизм имеет важное значение в регуляции уровня внутриклеточного Ca2*. Через щелевые контакты проходят низкомолекулярные вещества, регулирующие рост и развитие клеток. Щелевые контакты обеспечивают распространение возбуждения — переход ионов между мышечными клетками миокарда и между ГМК.
- Примеры
- Структура. В синапсе различают пресинаптическую часть, постсинаптическую часть и расположенную между клетками синаптическую щель.
- Нейромедиаторы — низкомолекулярные вещества, поступают из синаптичес
Межклеточные контакты — соединения между клетками, образованные при помощи белков. Межклеточные контакты обеспечивают непосредственную связь между клетками. Кроме того, клетки взаимодействуют друг с другом на расстоянии с помощью сигналов (главным образом — сигнальных веществ), передаваемых через межклеточное вещество.

Функции межклеточных соединений
Межклеточные соединения возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов (межклеточное взаимодействие), а также для механического скрепления клеток друг с другом.
Через щелевые контакты могут передаваться электрические сигналы. Клетки органов и тканей вырабатывают ряд химических веществ, действующих на другие клетки (в том числе через межклеточные контакты) и вызывающих изменения в работе цитоскелета, в интенсивности обмена веществ и процессе синтеза клеткой белков.
Типы межклеточных соединений
Простое межклеточное соединение
Плотное соединение (запирающая зона)
В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания.

Плотные контакты состоят из тонких лент, пересекающихся между собой, которые полностью опоясывают клетку и контактируют с аналогичными лентами на соседних клетках. На электронных микрофотографиях заметно, что в участках плотных контактов мембраны соприкасаются одна с другой или даже сливаются. Комбинация метода замораживания-скалывания с электронной микроскопией с высоким разрешением позволила установить, что плёнки плотных контактов построены из белковых частиц диаметром 3-4 нм, которые выступают с обеих поверхностей мембраны. Также в пользу того, что в образовании плотных контактов ключевую роль играют белки, свидетельствует деление клеток под действием протеолитического фермента трипсина.
Всего в состав тесных контактов входит около 40 различных белков, как мембранных, так и цитоплазматических. Последние необходимы для прикрепления актиновых филаментов, регуляции и сигнализирования.
Мембранные белки плотных контактов можно разделить на две группы: те, которые пересекают мембрану 4 раза, и те, которые пересекают её только раз. Первая группа значительно распространена, в неё входят белки клаудины, окклюдины и трицеллюлин. Они имеют общие черты строения, в частности в них имеются четыре α-спиральных трасмембранных домена, N- и С-концы обращены к цитозолю, а домены, выступающие в межклеточное пространство, участвуют в гомо- или гетерофильных взаимодействиях с подобными белками на соседней клетке.
Основными белками плотных контактов являются клаудины (лат. claudo). Их роль была продемонстрирована на примере мышей с отсутствующим геном клаудин-1, — в эпидермисе таких животных не формируются плотные контакты и они погибают в течение дня после рождения из-за обезвоживания вследствие интенсивного испарения[1]. Клаудины также участвуют в формировании селективных каналов для транспорта ионов. В геноме человека есть гены по крайней мере 24 различных клаудинов, экспрессия которых происходит тканеспецифически.
Второе место по распространенности в плотных контактах занимают белки окклюдины (от лат. occludo — закрывать), они регулируют транспорт маленьких гидрофильных молекул и прохождение нейтрофилов через эпителий. Наибольшие концентрации третьего белка — трицеллюлина, наблюдаются в местах контакта трех клеток.
К белкам плотных контактов, пересекающим мембрану один раз, относятся JAM-A,-B,-C и-D (англ. junctional adhesion molecules) и родственные им CAR (англ. coxsackievirus and adenovirus receptor), CLMP (англ. CAR-like membrane protein) и ESAM (англ. endothelial-cell selective adhesion molecule), имеющие по два иммуноглобулинных домена, а также белки CRB3 (англ. Crumbs homologue 3) и Bves
Цитоплазматическая пластинка плотных контактов необходима для их присоединения к актиновым филаментам, регуляции сцепления клеток и параклеточного транспорта, а также для передачи сигналов от поверхности внутрь клетки. В её состав входят адаптерные, каркасные и цитоскелетные белки, а также элементы сигнальных путей (киназы, фосфатазы). Наиболее изучен белок цитоплазматической пластинки — ZO-1, он имеет несколько доменов белок-белкового взаимодействия, каждый из которых обеспечивает контакт с другими компонентами, в том числе три PDZ-домена (англ. PSD95–DlgA–ZO-1) — с клаудинами и другими адаптерными белками — ZO-2 и ZO-3, GUK-домен (англ. guanylate kinase homology) — с окклюдинами, а SH3-домен — с сигнальными белками.
С цитоплазматической стороной плотных контактов также ассоциированы комплексы белков PAR3/PAR6 и Pals1/PATJ, необходимые для установления полярности клеток и эпителиального морфогенеза.
Функции Первые исследования функций плотных контактов привели к представлению, что это статические непроницаемые структуры, необходимые для того, чтобы ограничить диффузию веществ между клетками. Впоследствии было выяснено, что они избирательно проницаемы, к тому же их пропускная способность отличается в различных тканях и может регулироваться. Также установлена ещё одна функция плотных контактов: роль в поддержании полярности клеток путем ограничения диффузии липидов и белков во внешнем слое плазматической мембраны. В первом десятилетии 21 века также накоплены данные, свидетельствующие об участии этих структур в сигнальных путях, в частности, регулирующих пролиферацию и полярность.
Регулирование парацеллюлярного транспорта
Непроницаемость плотных контактов в большинстве водорастворимых соединений может быть продемонстрирована в опыте по введению гидроксида лантана (электронно плотный коллоидный раствор) в кровеносные сосуды поджелудочной железы. Через несколько минут после инъекции ацинарные клетки фиксируются, и из них готовятся препараты для микроскопии. В таком случае можно наблюдать, что гидроксид лантана диффундирует из крови в пространство между латеральными поверхностями клеток, но не может проникнуть через плотные контакты в их верхней части. Другие опыты показали, что плотные контакты также непроницаемы для солей. Например при выращивании почек собаки MDCK (англ. Madin-Darby canine kidney) в среде с очень низкой концентрацией кальция, они формируют монослой, однако не сочетаются между собой плотными контактами. Через такой монослой могут свободно двигаться соли и жидкости. Если культуре добавить кальция, то за час формируются плотные контакты, и слой становится непроницаемым для жидкостей.
Однако не во всех тканях плотные контакты полностью непроницаемы, существуют так называемые неплотные эпителии (англ. leaky epithelia). Например, эпителий тонкого кишечника пропускает в 1000 раз больше ионов Na +, чем эпителий канальцев почек. Ионы проникают через параклеточные поры диаметром 4 Å, селективные по заряду и размеру частиц, которые формируются белками клаудинами. Поскольку эпителии различных органов эксрессируют различные наборы клаудинов, то отличается и их проницаемость для ионов. Например, специфический клаудин, присутствуюий только в почках, позволяет проходить ионам магния в процессе реабсорбции.
Межклеточное пространство эпителия может быть проницаемым и для больших частиц, например, при повторении упомянутого опыта с гидроксидом лантана на ткани эпителия тонкого кишечника кролика можно наблюдать прохождение коллоидных частиц между клетками. Крупные молекулы транспортируются через специальные пути утечки (англ. leak pathway) диаметром более 60 Å. Это важно, например, для процессов всасывания аминокислот и моносахаридов, концентрация которых в тонком кишечнике возрастает после еды достаточно для их пассивного транспорта.
Поддержание различия между апикальной и базолатеральной мембранами
Если в среду, контактирующую с апикальной частью монослоя MDCK-клеток, добавить липосомы, содержащие флуоресцентно меченые гликопротеины, некоторые из них спонтанно сливаются с клеточными мембранами. После этого флуоресценцию можно обнаружить в апикальной, но не в базолатеральной части клеток при условии целостности плотных контактов. Если же их разрушить, удалив из среды кальций, флуоресцентные белки диффундируют и равномерно распределяются по всей поверхности клетки.
Цитозольный слой мембраны имеет одинаковый липидный состав, как в апикальном, так и в базолатеральном участках, эти липиды могут свободно диффундировать. С другой стороны, липиды внеклеточного слоя двух частей клетки существенно различаются, и обмену между ними препятствуют плотные контакты. Например, все гликолипиды, как и белки заякоренные гликозилфосфатидилинозитолом, в мембранах MDCK клеток расположены исключительно в внеклеточном слое апикальной части, а фосфатидилхолин — почти исключительно в базолатеральной части.
Болезни, связанные с плотными контактами
С нарушением формирования тесных контактов связаны некоторые наследственные расстройства человека, например мутации в генах клаудина-16 и клаудина-19, которые приводят к гипомагниемии, вследствие чрезмерной потери магния с мочой. Мутации в гене клаудина-13 и трицеллюлина вызывают наследственную глухоту. Дисрегуляция некоторых белков плотных контактов связана с онкологическими заболеваниями, например экспрессия ZO-1 и ZO-2 снижается во многих типах рака. Компоненты тесных контактов также могут быть мишенями для онкогенных вирусов.
Некоторые вирусы используют мембранные белки плотных контактов для проникновения в клетку, в частности клаудин-1 является корецептором для вируса гепатита C. Другие вирусы присоединяются к белкам плотных контактов, чтобы разрушить барьер, отделяющий их от настоящих рецепторов на базолатеральной слое эпителиальных клеток, или неэпителиальных клетках.
Плотные контакты могут быть мишенью и для бактериальных патогенов, например Clostridium perfringens — возбудитель газовой гангрены, выделяет энтеротоксин (англ.), действующий на внеклеточные домены мембранных клаудинов и окклюдинов, и вызывает протечки эпителия. Helicobacter pylori — возбудитель гастрита — вводит в клетки белок CagA, взаимодействующий с комплексом ZO-1-JAM-A, считается, что это помогает бактерии преодолеть защитный барьер желудочного эпителия.
Адгезионные контакты

Адгезионные контакты (англ. adherens junctions, AJ) — якорные межклеточные контакты, ассоциированные с микрофиламентами, обеспечивающие целостность и механическую прочность ткани, в частности противостояние растяжению, придающие клеткам возможность координированно использовать актиновый цитоскелет. Адгезионные контакты относятся к гомофильным, то есть соединяют клетки одинакового типа. В их формировании принимают участие белки кадгерины и катенины.
Морфологически адгезионные контакты являются относительно простыми, в отличие от десмосом, плотных и щелевых контактов, они не имеют высокоспециализированных ультраструктур, кроме скопления актиновых филаментов. От других типов соединений клеток они отличаются относительной гибкостью и изменчивостью.
Чаще всего адгезионные контакты встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки поясок, который называют зоной прилипания (англ. zonula adherens). Такие зоны в эпителии позвоночных животных преимущественно размещаются базальнее участка плотных контактов (англ. zonula occludens) и апикальнее десмосом (англ. macula adherens).
В зоне адгезионных контактов мембраны соседних клеток удалены друг от друга на расстояние 10-20 нм. В состав адгезионных контактов входят три основных элемента:
Схематическое изображение процесса формирования трубки из слоя эпителиальных клеток Одной из основных функций адгезионных контактов является физическое соединение клеток в единую ткань, их ослабление часто приводит к диссоциации клеток. Такого эффекта можно достичь, обработав ткань или монослойную культуру хелатирующим агентами, такими как ЭДТА, связывающими ионы кальция, вследствие чего взаимодействие между кадгеринами нарушается. Однако, хелатирующих агентов обычно недостаточно для полного разделения клеток, поскольку между ними существуют другие — кальций независимые — контакты.
Адгезионные контакты обеспечивают образование широкой межклеточной сети из пучков сократительных актиновых филаментов, расположенных параллельно мембранам клеток и соединенных между собой с помощью белков катенинов и кадгеринов. Такая организация позволяет не только противостоять механическому напряжению, но и согласовывать поведение клеток во время процессов морфогенеза. Например, координированное сокращение колец актиновых филаментов соседних клеток является необходимым для формирования трубок из слоя эпителия, в частности во время закладки нервной трубки. Одним из примеров является Shroom3-зависимое сокращение зоны прилипания, при этом актин-связывающий белок Shroom3 привлекает к участку адгезионные контакты Rho-киназы и активирует миозин-II, в результате чего и происходит сокращение.
Адгезионные контакты также задействованы в межклеточной передаче сигналов, об этом свидетельствует локализация в зрелых контактах рецепторной тирозинфосфатазы μ и белка RACK1, который взаимодействует с ней. При снижении экспрессии α-катенина наблюдается значительное увеличение темпов пролиферации эпителиальных клеток, было показано, что за этот эффект отвечает сигнальный путь инсулин / MAPK.
Читайте также: