
Механизм транспорта веществ через мембрану нейрона кратко
Обновлено: 07.07.2024
Транспортные структуры мембраны
Транспортные структуры мембраны - это специальные белковые структуры, встроенные в мембрану и обеспечивающие трансмембранный транспорт, т.е. перенос веществ через мембрану.
Мы уже сказали в предыдущем разделе 1_1 Строение клеточной мембраны, что многие вещества могут проходить сквозь клеточную мембрану самостоятельно, за счёт простой диффузии. Они "автоматически" перемещаются из зоны своей повышенной концентрации в зону пониженной концентрации, т. к. стремятся уравнять свою концентрацию по обе стороны мембраны. Так обычно ведут себя все вещества в растворах: они стремятся равномерно распределиться по всему объёму жидкости, пытаясь преодолеть разделяющую раствор на отсеки преграду. В принципе, любая молекула может пройти через липидный бислой клеточной мембраны, потому что составляющие его молекулы липидов сохраняют некоторую подвижность относительно друг друга и могут временами раздвигаться в стороны, пропуская различные вещества. Однако скорость такой пассивной диффузии, т.е. перехода вещества через мембрану из области с большей концентрацией в область с меньшей, может сильно различаться для разных веществ. Для многих веществ диффузия занимает столь длительное время, что можно говорить о практической непроницаемости для них мембраны. Скорость диффузии различных веществ через мембрану зависит главным образом от размера их молекул, электрического заряда (полярности) и их относительной растворимости в жирах.
Легче и лучше всего через мембрану пассивно проникают жирорастворимые неполярные мелкие молекулы.
Для транспорта в клетку веществ, слабо способных к диффузии через липидный слой мембраны, необходимы специальные транспортные структуры.
Виды транспортных структур мембраны:
1. Ионные каналы - это специальные молекулярные трубочки с порами (дырочками) в мембране, образованные канальными белками, позволяющие ионам проходить через мембрану в обоих направлениях: как внутрь, так и наружу. Ионные каналы могут открываться при определённых условиях, в этом случае они являются управляемыми этими условиями. Через открытую пору ионного канала ионы могут свободно диффундировать, не вызывая конформационных изменений в белке канала. При токе через отдельный канал в 1 пА по нему проходят 6×10 6 одновалентных ионов в секунду.
2. Транслоказы, - специальные мембранные белки, облегчающие переход вещества через мембрану за счёт своего временного связывания с диффундирующим веществом. Не требуют энергии, работают в обоих направлениях в зависимости от концентрации переносимого вещества. Раньше их представляли себе как некие "белки-переносчики" в виде "челноков", но когда стали известны аминокислотные последовательности транспортных белков, то стало понятно, что все они обычно содержат многочисленные трансмембранные домены и образуют сквозной путь через мембрану. Транспортёр, связываясь с субстратом по одну сторону мембраны, изменяет свою конформацию для переноса субстрата через мембрану. При ко-транспорте, в дополнение к сказанному, повышается аффинность транспортёра в новой конформации ко второму субстрату, и субстраты переносятся через мембрану сопряженно, совместно.
3. Транспортёры - белковые структуры, насильно протаскивающие определённые вещества сквозь клеточную мембрану в определённом направлении с затратами энергии. Ионные насосы - это транспортёры ионов. По способу использования энергии для своей работы транспортёры можно разделить на "симпортные" и "антипортные". Симпортные транспортёры используют совместный транспорт в одном направлении двух веществ: одно из них должно иметь большую потенциальную энергию для движения через мембрану. Например, симпорт в клетку с помощью ионов натрия глюкозы, или симпорт ионов кальция с помощью ионов натрия. Антипортные транспортёры (обменники) используют встречный транспорт двух веществ с разной потенциальной энергией диффузии. Так работает, например, натрий-калиевый ионный насос.
Итак, перенос веществ через клеточную мембрану происходит различными путями.
Механизмы транспорта веществ через мембрану
1. Простая диффузия жирорастворимых (гидрофобных) веществ через жировой слой мембраны. Это пассивный процесс под действием градиента (перепада) концентрации вещества по разные стороны мембраны. (Смотрите видео 1 : пассивный транспорт через мембрану).
2. Неуправляемая диффузия (неуправляемый пассивный перенос) водорастворимых веществ через постоянно открытые ионные каналы мембраны.
3. Управляемая диффузия (управляемый пассивный перенос) водорастворимых веществ через управляемые ионные каналы мембраны. (Смотри: Ионные каналы мембраны)
4. Активный транспорт водорастворимых веществ с помощью специальных белковых транспортных структур (транспортёров) за счёт использования энергии расщепления АТФ. (Смотрите видео 2 : активный транспорт через мембрану).
5. Эндоцитоз крупных частиц за счёт образования мембранных пузырьков.
Видео 3 : Эндоцитоз и экзоцитоз
Видео 4 : Фагоцитоз (эндоцитоз) бактерий макрофагом
Как это происходит? Вот этот вопрос мы сейчас и рассмотрим.
Видеоанимация 5 : Пассивный мембранный транспорт
Видео 6 : Мембрана и транспорт веществ
Видео 7 : Мембранный транспорт
1. Транспортёры мембраны - ферменты по имени АТФазы
Одна из самых главных транспортных структур мембраны — это фермент АТФаза (произносится как "а\тэ\эф\аза"). АТФазы разных видов транспортируют через мембрану ионы. Они переносят их как внутрь клетки, так и, наоборот, наружу. Поэтому их называют "ионными насосами" или "ионными помпами". Электрическая проводимость ионных насосов очень мала: Na/K-АТФаза переносит всего около 300 Na + и 200 K + в секунду.
Название АТФаза означает, что это фермент, нацеленный на расщепление АТФ, его полное название - аденозинтрифосфатаза.
Только не надо думать, что АТФаза существует в единственном варианте! В настоящее время уже обраружено множество различных видов транспортных АТФаз. Они схожи между собой по строению и механизму действия, но имеют разную специализацию, т.е. каждый их вид перетаскивает через мембрану что-то своё. В настоящее время достаточно хорошо изучены Na + /K + -АТФаза, Ca 2+ -АТФаза, H + -АТФаза, H + ,K + -АТФаза, Mg 2+ -АТФаза, которые обеспечивают перемещение соответственно ионов Na + , K + , Ca 2+ , H + , Mg 2+ изолированно или сопряжённо: например, Na + сопряжённо с К + ; Н + сопряжённо с К + .
В чём принцип работы АТФазы?
Самой главной мембранной АТФазой по праву можно считать Na,K-АТФазу (натрий-калиевую аденозинтрифосфатазу).

По своей структуре она является представителем гетеродимерных АТФаз Р-типа.
При этом следует помнить, что, как истинный фермент, Na,K-АТФаза параллельно расщепляет АТФ, получая от этого энергию на свою транспортную деятельность.
Далее цикл повторяется.
Гипотеза работы Na,K-АТФазы рассмотрена подробнее здесь: Механизм натрий-калиевого насоса
Какие есть ещё АТФазы?
Н + ,К + -АТФазы обеспечивают секрецию соляной кислоты париетальными клетками желудка. Они перемещают на наружную сторону мембраны ионы водорода, которые создают кислую среду в желудке. Этот транспорт тоже работает по принципу обменника, т.к. меняет внутриклеточные ионы водорода на внеклеточные ионы калия. Кстати, в мембрану боковой и базальной поверхности этих клеток встроен хлорно-бикарбонатный анионообменни, через который анионы: Cl - вводятся в клетку в обмен на HCO3 -
Н + -АТФаза растений обеспечивает поглощение из почвы солей корнями растений. Принцип действия тот же: обмен одних ионов на другие за счёт энергии, полученной из АТФ. Из клеток корня в почву выделяются ионы водорода Н+, а на их место в клетку переносятся ионы солей.
Протонная АТФаза грибка Neurospora crassa состоит из 920 аминокислот (источник: Hager et al. 1986).
Са 2+ -АТФаза саркоплазматического ретикулюма в мышечных клетках обеспечивает транспорт кальция из цитоплазмы мышечных клеток во внутриклеточные цистерны для депонирования (запасания) кальция.

Выводы
Специальные транспортные ферменты АТФазы, встроенные в клеточную мембрану, работают как транспортёры для различных веществ. Они насильно переносят вещества в клетку и из клетки. При этом АТФазы получают на свой активный транспорт энергию за счёт расщепления АТФ.
Активный транспорт веществ через мембрану клетки - это насильственный перенос вещества с затратой энергии. Он возможен даже против градиента концентрации вещества, т.е. из зоны пониженной концентрации в зону повышенной концентрации.
На рисунке справа - структура кальциевой АТФазы по Toyoshima et al. Nature 405 (2000) 647-655 PDB ID: 1EUL.pdb
2. Транспортёры глюкозы
Глюкозные транспортёры - это белки, переносящие глюкозу через мембрану. Их называют белками-переносчиками, а также рецепторами глюкозы. Эти белки образуют гидрофильные трансмембранные каналы.
Глюкозные трнанспортёры делятся на две группы.
1. Na + -глюкозные ко-транспортёры (симпортёры). Эти транспортёры занимаются активным транспортом глюкозы с помощью ионов Na + и их работа зависит от градиента концентрации натрия. Они работают только в почечных канальцах и кишечнике, обеспечивая всасывание глюкозы против градиента её концентрации.
2. Транспортные белки семейства ГЛЮТ. Они отличаются от сходных по функции белков, транспортирующих глюкозу через мембрану в кишечнике и почках, и обеспечивают облегчённую диффузию, а не активный транспорт. Белки ГЛЮТ обнаружены во всех тканях и их существует несколько разновидностей.
Все 5 типов ГЛЮТ имеют сходную первичную структуру и доменную организацию.
- ГЛЮТ-1 (эритроцитарный тип) обеспечивает стабильный поток глюкозы в глиальные клетки мозга.
- ГЛЮТ-2 (печёночный тип) обнаружен в клетках органов, выделяющих глюкозу в кровь. Именно при участии ГЛЮТ-2 глюкоза переходит в кровь из энтероцитов и печени. ГЛЮТ-2 участвует в транспорте глюкозы в β-клетки поджелудочной железы. В то же время ГЛЮТ-2 обеспечивает проникновение глюкозы из крови в клетки печени (гепатоциты) по механизму облегчённой диффузии. Там глюкоза превращается в активное вещество глюкозо-6-фосфат, участвующее в обмене углеводов, жиров и в энергообмене.
- ГЛЮТ-3 (мозговой тип) обладает большим, чем ГЛЮТ-1, сродством к глюкозе. Он также обеспечивает постоянный приток глюкозы к клеткам нервной и других тканей.
- ГЛЮТ-4 (мышечно-жировой тип) - главный переносчик глюкозы в клетки мышц и жировой ткани. Это единственный переносчик, регулируемый инсулином, поэтому мышцы и жировую ткань называют инсулинзависимыми тканями.
- ГЛЮТ-5 (кишечный тип) встречается, главным образом, в клетках тонкого кишечника.
Влияние инсулина на транспортёры ГЛЮТ
Все типы ГЛЮТ могут находиться как в наружной мембране клетки, так и в цитозольных везикулах. ГЛЮТ-4 (и в меньшей мере ГЛЮТ-1) почти полностью находятся в цитоплазме клеток. Влияние инсулина на такие клетки приводит к перемещению везикул, содержащих ГЛЮТ, к плазматической мембране, слиянию с ней и встраиванию этих белков-транспортёров в мембрану. После этого становится возможным облегчённый транспорт глюкозы в эти клетки. Скорость потребления глюкозы возрастает в 30-40 раз. После снижения концентрации инсулина в крови транспортёры глюкозы снова перемещаются в цитоплазму, и поступление глюкозы в клетку прекращается.
Видео 8 : Транспорт веществ в клетку
Видео 9 : Виды клеточного транспорта (англ.яз.)
Строение и физиология нейрона. Внутриклеточный транспорт нейрона
Миллиарды нейронов формируют поверхностный слой — кору— полушарий головного мозга и полушарий мозжечка. Кроме того, в толще белого вещества нейроны образуют скопления—ядра.
Практически все нейроны ЦНС мультиполярны: сома (тело) нейронов характеризуется наличием нескольких полюсов (вершин). От каждого полюса, за исключением одного, отходят отростки — дендриты, которые образуют многочисленные разветвления. Дендритные стволы могут быть гладкими или образовывать многочисленные шипики. Дендриты образуют синапсы с другими нейронами в области шипиков или ствола дендритного дерева.
От оставшегося полюса сомы отходит отросток, проводящий нервные импульсы,— аксон. Большинство аксонов формирует коллатеральные ветви. Концевые ветви образуют синапсы с нейронами-мишенями.
Нейроны образуют два основных типа синаптических контактов: аксодендритические и аксосоматические. Аксодендритические синапсы в большинстве случаев передают возбуждающие импульсы, а аксосоматические — тормозящие.
Формы нейронов мозга.
(1) Пирамидальные нейроны коры полушарий.
(2) Нейроэндокринные нейроны гипоталамуса.
(3) Шипиковые нейроны полосатого тела.
(4) Корзинчатые нейроны мозжечка. Дендриты нейронов 1 и 3 образуют шипики.
А — аксон; Д — дендрит; КА — коллатерали аксона. Дендритные шипики.
Срез мозжечка, на котором имеются дендриты гигантских клеток Пуркинье, образующие шипики.
В поле зрения различимы три шипика (Ш), образующие синаптические контакты с булавовидными расширениями аксонов (А).
Четвертый аксон (слева вверху) образует синапс с дендритным стволом. (А) Двигательный нейрон переднего рога серого вещества спинного мозга.
(Б) Увеличенное изображение (А). Миелиновые оболочки участков 1 и 2, располагающихся в белом веществе ЦНС, образованы олигодендроцитами.
Возвратная коллатеральная ветвь аксона начинается от немиелинизированного участка.
Миелиновые оболочки участков 3 и 4, относящихся к периферической части нервной системы, образованы шванновскими клетками.
Утолщение аксона в области вхождения в спинной мозг (переходного участка) соприкасается с одной стороны с олигодендроцитом, а с другой—со шванновской клеткой.
(В) Нейрофибриллы, состоящие из нейрофиламентов, видны после окрашивания солями серебра.
(Г) Тельца Ниссля (глыбки гранулярной эндоплазматической сети) видны при окрашивании катионными красителями (например, тионином).
Внутреннее строение нейронов
Цитоскелет всех структур нейрона образован микротрубочками и нейрофиламентами. Тело нейрона содержит ядро и окружающую его цитоплазму— перикарион (греч.peri— вокруг и karyon—ядро). В перикарионе расположены цистерны гранулярной (шероховатой) эндоплазматической сети — тельца Ниссля, а также комплекс Гольджи, свободные рибосомы, митохондрии и агранулярная (гладкая) эндоплазматическая сеть.
1. Внутриклеточный транспорт. В нейронах происходит обмен веществ между мембранными структурами и компонентами цитоскелета: непрерывно синтезируемые в соме новые клеточные компоненты перемещаются в аксоны и дендриты путем антероградного транспорта, а продукты метаболизма поступают путем ретроградного транспорта в сому, где происходит их лизосомальное разрушение (распознавание клеток-мишеней).
Выделяют быстрый и медленный антероградный транспорт. Быстрый транспорт (300-400 мм в сутки) осуществляют свободные клеточные элементы: синаптические пузырьки, медиаторы (или их предшественники), митохондрии, а также липидные и белковые молекулы (в том числе и белки-рецепторы), погруженные в плазматическую мембрану клетки. Медленный транспорт (5-10 мм в сутки) обеспечивают компоненты цнто-скелета и растворимые белки, в том числе и некоторые белки, задействованные в процессе высвобождения медиаторов в нервных окончаниях.
Первым среди нейротрофинов был изучен фактор роста нервов, выполняющий особенно важные функции в развитии периферической чувствительной и вегетативной нервной системы. В соме нейронов зрелого мозга синтезируется фактор роста, выделенный из головного мозга (BDNF), который транспортируется антероградно в их нервные окончания. Согласно данным, полученным в результате исследований на животных, фактор роста, выделенный из головного мозга, обеспечивает жизнедеятельность нейронов, принимая участие в обмене веществ, проведении импульсов и синаптической передаче.
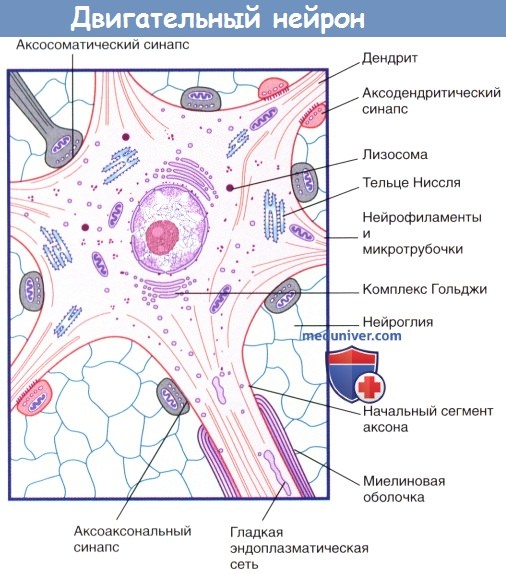
Внутреннее строение двигательного нейрона.
Изображены пять дендритных стволов, три возбуждающих синапса (выделены красным цветом) и пять тормозных синапсов.
2. Механизмы транспорта. В процессе нейронального транспорта роль поддерживающих структур выполняют микротрубочки. Связанные с микротрубочками белки перемещают органеллы и молекулы вдоль внешней поверхности миктротрубочек за счет энергии АТФ. Антероградный и ретроградный транспорт обеспечивают разные виды АТФаз. Ретроградный транспорт осуществляется за счет динеиновых АТФаз. Нарушение функционирования динеинов приводит к болезни двигательного нейрона.
Ниже описано клиническое значение нейронального транспорта.
Столбняк. При загрязнении раны почвой возможно заражение столбнячной палочкой (Clostridium tetani). Этот микроорганизм продуцирует токсин, который связывается с плазматическими мембранами нервных окончаний, проникает путем эндоцитоза в клетки и посредством ретроградного транспорта попадает в нейроны спинного мозга. Нейроны, расположенные на более высоких уровнях, также захватывают этот токсин путем эндоцитоза. Среди этих клеток необходимо особенно отметить клетки Реншоу, которые в норме оказывают тормозное действие на двигательные нейроны путем выделения тормозного медиатора—глицина.
При поглощении клетками токсина выделение глицина нарушается, вследствие чего прекращаются тормозные влияния на нейроны, осуществляющие двигательную иннервацию мышц лица, челюстей и позвоночника. Клинически это проявляется длительными и изнурительными спазмами этих мышц и в половине случаев заканчивается гибелью пациентов от истощения в течение нескольких дней. Предотвратить столбняк возможно, проведя своевременную иммунизацию в должном объеме.
Вирусы и токсичные металлы. Считают, что за счет ретроградного аксонального транспорта происходит распространение вирусов (например, вируса простого герпеса) из носоглотки в ЦНС, а также перенос токсичных металлов—алюминия и свинца. В частности, распространение вирусов по структурам мозга осуществляется за счет ретроградного межнейронального переноса.
Периферические нейропатии. Нарушение антероградного транспорта — одна из причин дистальных аксональных нейропатий, при которых развивается прогрессирующая атрофия дистальных участков длинных периферических нервов.
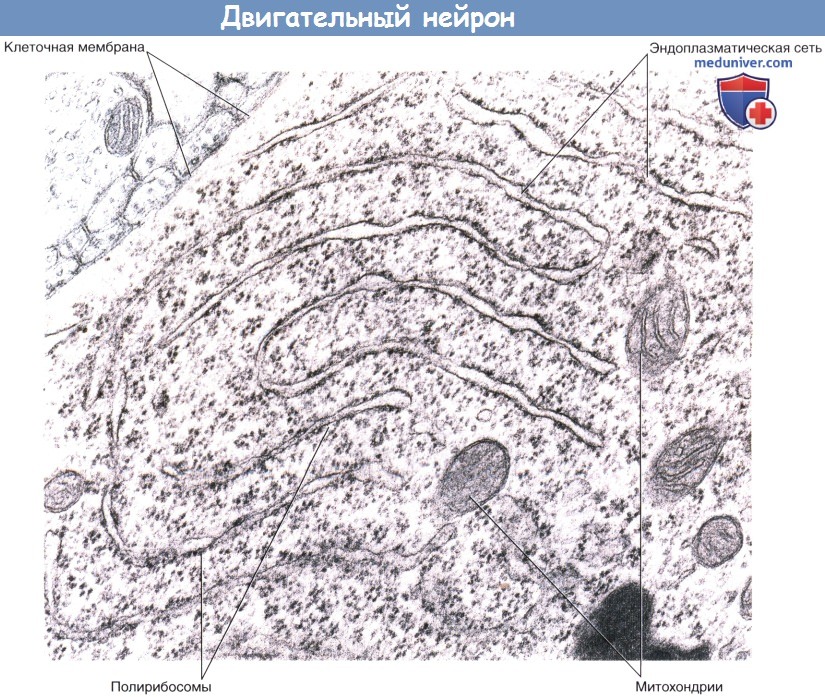
Тельце Ниссля в соме двигательного нейрона.
Эндоплазматическая сеть имеет многоуровневую структуру. Полирибосомы образуют выросты на внешних поверхностях цистерн или свободно лежат в цитоплазме.
(Примечание: для лучшей визуализации структуры слабо окрашены).
Читайте также: