
Механизм шагательного рефлекса физиология кратко
Обновлено: 12.05.2024
Б. Особая роль проприорецепторов в осуществлении шагательного рефлекса определяется их расположением. Мышечные веретена (мышечные рецепторы) расположены параллельно скелетной мышце - своими концами они крепятся к соединительнотканной оболочке (перимизий) пучка экстрафузальных (fusus - веретено) мышечных волокон при помощи напоминающих сухожилия полосок соединительной ткани длиной 0,5-1,0 мм (рис. 5.6). Поэтому, когда мышца расслабляется (удлиняется), растягиваются и мышечные рецепторы, что и ведет к их возбуждению (см. рис. 5.6, а).
Мышечный рецептор состоит из нескольких поперечнополосатых интрафузальных мышечных волокон, окруженных соединительнотканной капсулой. Длина волокон равна 4-7 мм, толщина составляет 15-30 мкм.
Длина экстрафузальных мышечных волокон - от нескольких миллиметров до многих сантиметров, толщина составляет 50-100 мкм. Вокруг средней части мышечного веретена несколько раз обвивается окончание одного афферентного волокна. Многие мышечные веретена иннервируются и другим - более тонким - афферентным волокном, которое обвивает в виде спирали периферические участки мышечного рецептора. Импульсы от мышечных рецепторов возбуждают нейроны своего центра (гомонимные нейроны) и тормозят нейроны центра-антагониста. Ближе к концам мышечного веретена подходят двигательные нервные окончания, являющиеся аксонами ?-мотонейронов спинного мозга. Их импульсация вызывает сокращение интрафузальных мышечных волокон (мышечных волокон рецептора), что ведет к возбуждению рецепторов.

Рис. 5.6. Схема, отражающая процессы возбуждения ?-мотонейронов спинного мозга и их торможения с помощью вставочных тормозных нейронов во время сокращения и расслабления скелетной мышцы. 1 - мышечные рецепторы (мышечные веретена); 2 - сухожилия мышцы и рецепторы (рецепторы Гольджи); 3 - полусегмент спинного мозга; а - мышца растянута и расслаблена - возбуждаются мышечные рецепторы (1); б - мышца сокращена, напряжена и укорочена - возбуждаются сухожильные рецепторы (2);
сплошные линии - импульсация выражена; пунктирные линии - импульсация отсутствует
Сухожильные рецепторы (рецепторы Гольджи) заключены в соединительнотканную капсулу и локализуются в сухожилиях скелетных мышц вблизи сухожильно-мышечного соединения. Рецепторы представляют собой безмиелиновые окончания толстого миелинового афферентного волокна (подойдя к капсуле рецепторов Гольджи, это волокно теряет миелиновую оболочку и делится на несколько окончаний). Сухожильные рецепторы крепятся к скелетной мышце последовательно, что обеспечивает их раздражение при натяжении сухожилия (см. рис. 5.6,б). Поэтому сухожильные рецепторы посылают в мозг информацию о том, что мышца сокращена (напряжено и сухожилие), а мышечные рецепторы - о том, что мышца расслаблена и удлинена. Импульсы от сухожильных рецепторов тормозят нейроны своего центра и возбуждают нейроны центра-антагониста (у мышц-сгибателей это возбуждение выражено слабее).
В. Элемент шагательного рефлекса - поочередное сокращение и расслабление скелетной мышцы под влиянием импульсов от проприорецепторов, поступающих в свой центр.
Когда мышца (сгибатель или разгибатель) расслаблена и удлинена, возбуждаются мышечные веретена, импульсы от которых поступают к своим ?-мотонейронам спинного мозга и возбуждают их (рис. 5.6, а). Далее ?-мотонейроны посылают импульсы к этой же скелетной мышце, вызывая ее сокращение. Как только мышца сократилась (рис. 5.6, б), возбуждение мышечных веретен прекращается или сильно ослабляется (они уже не растянуты), но начинают возбуждаться сухожильные рецепторы. Импульсы от рецепторов также поступают в первую очередь в свой центр спинного мозга, но к тормозным клеткам Реншоу. Возбуждение тормозных клеток вызывает торможение ?-мотонейронов этой же скелетной мышцы, вследствие чего она расслабляется. Однако ее расслабление (удлинение) снова ведет к возбуждению мышечных веретен и ?-мотонейронов - мышца вновь сокращается. Вследствие ее сокращения возбуждаются сухожильные рецепторы и тормозные клетки в спинном мозге, что вновь ведет к расслаблению скелетной мышцы и т.д. Мышца поочередно сокращается и расслабляется в результате поступления к ее мотонейронам импульсов от собственных рецепторов. Описанные процессы в равной степени относятся и к мышце-сгибателю, и к мышце-разгибателю. При этом расслабление скелетной мышцы запускает механизмы ее сокращения, а сокращение скелетной мышцы активирует механизмы, расслабляющие мышцу.

Рис. 5.7. Рефлекторные дуги шагательного рефлекса. А - конечность согнута; Б - контралатеральная конечность разогнута;
В - коленные суставы с мышцами; Г - полусегменты спинного мозга;
1 - сухожилия и их рецепторы (рецепторы Гольджи); 2 - мышечные рецепторы (мышечные веретена); ? афферентные пути от проприорецепторов;
Шагание на месте. Чесательный рефлекс и рефлексы мышечного спазма
а) Диагональное шагание всех четырех конечностей - рефлекс шагания на месте. Если спинальное животное (выхоженное после высокой перерезки спинного мозга на уровне шеи над областью иннервации верхних конечностей) удерживать над полом так, чтобы его лапы свободно свисали вниз (для облегчения понимания просим вас изучить рисунок ниже), растяжение конечностей иногда возбуждает шагательные рефлексы, вовлекающие все четыре конечности.

Диагональные шагательные движения, выполняемые спинальным животным
В целом, шагательные движения передних и задних конечностей осуществляются по диагонали. Эта диагональная реакция является еще одним проявлением реципрокной иннервации, на этот раз осуществляемой вдоль всей длины спинного мозга в направлении вверх и вниз между передними и задними конечностями. Такую организацию шагания называют рефлексом шагания на месте.
б) Рефлекс галопирования. Другим типом рефлекса у спинальных животных является рефлекс галопирования, при котором обе передние конечности движутся вместе назад, тогда как обе задние конечности движутся вперед. Это происходит, когда почти равные стимулы растяжения или давления действуют одновременно на конечности с обеих сторон тела; неравная стимуляция вызывает диагональный шагательный рефлекс. Такое представление поддерживается особенностями шагания и галопа в нормальных условиях. При шагании одновременно стимулируются только одна передняя и одна задняя конечности, что способствует продолжению шагания.
Во время галопа все конечности стимулируются примерно в равной степени, и животное сразу ударяет о землю обеими передними и обеими задними конечностями.
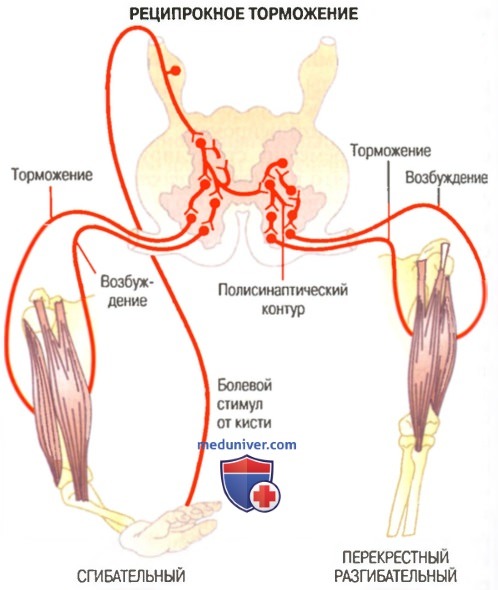
Сгибательный рефлекс, перекрестный разгибательный рефлекс и реципрокное торможение
Чесательный рефлекс
Особенно важным спинальным рефлексом у некоторых животных является чесательный рефлекс, который запускается ощущениями зуда или щекотки. Этот рефлекс включает две функции: (1) ощущение положения, которое позволяет лапе точно найти место раздражения на поверхности тела; (2) чесателъные движения вперед-назад.
Ощущение положения чесательного рефлекса является высокоразвитой функцией. Так, если блоха заползает на плечо спинального животного, его задняя лапа может определить ее положение, несмотря на то, что для приведения лапы к месту, где ползает блоха, должны одновременно и весьма координированно сокращаться 19 мышц конечности. На еще большую сложность рефлекса указывает факт, что при переходе блохи на другую сторону первая лапа прекращает чесание, а противоположная лапа начинает движения вперед-назад и в итоге находит блоху.
Движения вперед-назад, как и шагательные локомоторные движения, вовлекают контуры с реципрокной иннервацией, которые вызывают осцилляцию.
Спинальные рефлексы, вызывающие мышечный спазм
У людей часто наблюдается локальный мышечный спазм. Во многих, если не в большинстве случаев, причиной такого спазма является локальная боль.
а) Мышечный спазм в результате перелома кости. Один тип клинически важного спазма происходит в мышцах, окружающих сломанную кость. Этот спазм является результатом болевых импульсов, возникающих в сломанных концах кости, что заставляет мышцы, окружающие эту область, сокращаться тонически. Уменьшение боли при введении локального анестетика в область перелома снижает спазм; глубокий общий наркоз, например эфирный, также уменьшает спазм. Часто предварительное использование одного из этих видов обезболивания необходимо для успешного преодоления спазма в процессе приведения концов переломанной кости в правильное положение.
б) Спазм брюшных мышц при перитоните. Другим типом локального спазма, вызываемого спинальными рефлексами, является спазм мышц передней брюшной стенки в результате раздражения пристеночной брюшины при перитоните. В этом случае облегчение боли позволяет спастически сокращенным мышцам расслабиться. Тот же тип спазма часто происходит во время хирургической операции. Например, при операции на брюшной полости болевые импульсы от пристеночной брюшины часто заставляют брюшные мышцы сильно сокращаться, иногда выдавливая кишечник через хирургический разрез. По этой причине для проведения операций внутри брюшной полости обычно требуется глубокая анестезия.
в) Судороги мышц. Другим типом локального спазма является типичная мышечная судорога. Электромиографические исследования указывают, что причина некоторых мышечных судорог следующая. Любой местный раздражающий фактор или нарушение метаболизма в мышце, например сильный холод, прекращение кровоснабжения или перенапряжение, может вызвать проведение болевых или других сенсорных сигналов от мышцы к спинному мозгу, что, в свою очередь, вызывает рефлекторное сокращение мышцы по механизму обратной связи. Полагают, что сокращение еще больше стимулирует сенсорные рецепторы, заставляя спинной мозг усилить интенсивность сокращения.
Таким образом, благодаря развитию положительной обратной связи даже небольшое первичное раздражение вызывает все более сильное сокращение, вплоть до развития выраженного судорожного сокращения мышцы.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021

Вполне логично, что первые теоретики психологии развития (А.Л. Гезелл, 1977 и др.) предполагали, что хорошо известные этапы моторного развития, фиксирующие достижения ребенка в управлении движениями головы, головы и туловища, в сидении, стоянии, ползании и ходьбе, обеспечиваются процессами созревания. Прогресс в развитии настолько универсален и неизбежен, что представлялся автоматическим следствием биологического роста.
Однако в настоящее время известно, что созревание этих моторных навыков отнюдь не является жестко запрограммированным. Напротив, овладение каким-либо навыком создает условия для обучения другим, более сложным навыкам. На моторное развитие оказывают влияние также и культурные факторы (Д. Баттерворт, М. Харрис, 2000).
В настоящее время накоплен уже достаточно большой фактический материал, свидетельствующий о том, что положительные эмоции у младенцев возникают и развиваются лишь под влиянием удовлетворения потребности во внешних впечатлениях (Э.Л. Фрухт, 1996; К.Л. Печора, 2000; Г.В. Пантюхина, 2001). Наблюдения показывают, что ребенок при удовлетворении потребности испытывает не только состояние довольства и успокоения, но при каких-то, пока еще не достаточно изученных условиях, и сильные положительные эмоции (Н.М. Щелованов, 1975; М.Ю. Кистяковская, 1978).
Известно также, что полноценное развитие органа, а тем более такого сложного, как полушария головного мозга, возможно лишь в результате его функционирования. Поэтому мозг нуждается в раздражителях, вызывающих его деятельность и тем самым обеспечивающих его морфологическое и функциональное развитие. Иначе говоря, движущей силой выступает не слабость ребенка перед окружающей средой, не стремление лишь приспособиться к ней, а напротив, стремление познать действительность и овладеть ею (Л.И. Божович, Д.И. Фельдштейн, 1995).
После рождения ребенок последовательно наращивает эффективность управления моторикой, типичный ход развития которой одним из первых описал М. Макгро (Д. Баттерворт, М. Харрис, 2000).
Сначала дети приобретают способность поднимать голову, потом голову и грудь, затем они приподнимают голову и туловище с упором на локти и запястья и, наконец, они поднимают себя на всех четырех конечностях. К 9-ти месяцам дети обычно ползают на четвереньках, хотя некоторые ползают на животе. Вскоре они могут встать прямо, а в 12 месяцев большинство детей делает свои первые шаги (М.М. Безруких с соавт., 2005).
Ранее высказывалось предположение о том, что может существовать преемственность между шагательным рефлексом новорожденных и последующими движениями при ползании и ходьбе (А. Пайпер, 1968).
Однако Ф. Зелазо (F. Zelaso, 1984) было установлено, что тренировка шагательного рефлекса предотвращает его исчезновение в более позднем возрасте и ведет к чуть более раннему началу ходьбы по сравнению с контрольной группой, не имевшей подобной тренировки (Д. Баттерворт, М. Харрис, 2000).
Существуют также наблюдения, относящиеся к некоторым африканским племенам таким, как кипсигам из Нигерии, которые помогают своим младенцам упражняться в принятии различных поз, что ведет, например, к ускорению на срок до 5-ти недель времени самостоятельного принятия сидячей позы. Подобные культурные обычаи ускоряют моторное развитие, однако еще до конца не ясно, происходит это благодаря накоплению специфического опыта или же за счет усиления общего мышечного тонуса и увеличения силы так, что дети становятся способны поддерживать свой вес в более раннем возрасте.
Необходимым условием достижения такого динамического равновесия является овладение статической позой тела. Поэтому младенцы сначала научаются поддерживать устойчивую позу головы, а потом сидячую и стоячую позы.
Нормальное развитие всех этих навыков требует участия зрения, в пользу чего говорят факты замедленного развития локомоции у детей с врожденной слепотой (S. Fraiberg, 1974).
Дж. Баттерворт и Д. Сичетти (1982) в своих исследованиях показали, что дети, которые только что научились сидеть без поддержки, должны учитывать поток зрительной информации для успешного контроля сидячей позы (Дж. Баттерворт, М. Харрис, 2000). Они заметно качаются, если их посадить в комнату с движущимися стенами, искажающую зрительную обратную связь.
После рождения, благодаря морфологическим и анатомическим особенностям головного мозга, тип развития движений у детей отличается от развития моторики у детенышей животных. Сравнение обоих типов позволяет считать, что чем большую роль играет кора головного мозга в развитии движений, тем неорганизованнее моторика новорожденных, тем продолжительнее период ее развития и тем сложнее и разнообразнее движения взрослого организма. Наконец, соотношение во времени развития функции рецепторов и моторики оказывается у ребенка иным, чем у детенышей животных (Л.О. Бадалян, 2001). Так, кора головного мозга ребенка становится деятельной около середины первого месяца жизни ребенка, о чем свидетельствуют условные рефлексы, которые можно получить в это время. Однако движения даже 2-месячного ребенка еще крайне несовершенны.
У большинства млекопитающих последовательность в развитии движений и деятельности рецепторов совершенно иная: их движения или уже организованы к моменту рождения, или формируются прежде, чем образуются условные рефлексы с высших анализаторов – слухового и зрительного. Таким образом, у ребенка вначале начинают функционировать высшие анализаторы и лишь затем развиваются сложные локомоторные акты, требующие сложной координации. Эта закономерность имеет важное практическое значение и свидетельствует о необходимости воспитания движений в определенной последовательности (Л.О. Бадалян, 2001).
К рождению формируются подкорковые образования двигательного анализатора, интегрирующие деятельность экстрапирамидной системы. Этот уровень, по Н.А. Бернштейну, называется таламопаллидарным. Движения новорожденного хаотичны, генерализованны, носят атетозоподобный характер, нецеленаправленны, наблюдается мышечная гипертония с преобладанием сгибателей (Н.А. Бернштейн, 1966).
Координация движений у детей начинает развиваться после рождения. Этот уровень организации движений назван пирамидно-стриарным. Вначале формируется координация мышц глаз, что проявляется у ребёнка на 2–3-й неделе в виде фиксации взора на ярком предмете, затем ребенок следит за движущейся высоко поднятой игрушкой, поворачивая голову, что уже свидетельствует о начале развития координации движений шейных мышц. К 1–2-му месяцу ребенок начинает держать голову. Затем начинают развиваться координированные движения рук – это приближение рук к глазам и носу, потирание их, а несколько позднее поднимание рук над лицом и разглядывание их (Д.А. Фарбер, 2005).
С 3–3,5 месяцев ребенок начинает ощупывать свои руки, перебирать пальцами одеяло и край пеленки. Теперь необходимы уже игрушки более мелкие, и их подвешивают невысоко над кроватью. В этот период начинают формироваться целенаправленные движения. Вначале ребенок начинает удерживать игрушку двумя руками, а затем постепенно начинает делать попытки и активно захватывать ее руками (с 12–13-й недели). На 4–5-м месяце развивается координация движений мышц спины, что проявляется вначале переворачиванием со спины на живот, а затем (в 5–6 месяцев) – с живота на спину. Лишь только с 5-го месяца протягивание руки и хватание предмета начинает напоминать аналогичные движения взрослого человека. Однако и здесь имеется ряд особенностей, прежде всего, это обилие сопутствующих нерациональных движений. Хватательные движения этого периода сопровождаются параллельными движениями второй руки, в связи, с чем можно говорить о двуручном хватании. Наконец, во время хватания движения возникают в ногах и в туловище, нередко происходит и открывание рта. Хватающая рука совершает много лишних ищущих движений; схватывание осуществляется исключительно ладонью, т.е. пальцы сгибаются так, чтобы прижать игрушку к ладони.
На 6-м месяце начинает формироваться навык самостоятельного сидения, что уже свидетельствует о развитии координации движений мышц ног. В последующем совершенствуется взаимодействие двигательного и зрительного анализаторов, что приводит к 7–8 месяцу к большей прицельности движения хватающей руки. С 9–10 месяца возникает ножницеобразное хватание посредством смыкания большого и II–III пальцев по всей длине. С 12–13-го месяца возникает клещеобразное хватание с использованием концевых фаланг большого и указательного пальцев. В течение всего периода детства постепенно угасают различные содружественные нерациональные движения (А.Б. Пальчик, 2002).
Читайте также: