
Летальные гены это кратко
Обновлено: 05.07.2024
§ 32. Взаимодействие аллельных генов. Анализирующее скрещивание
Взаимодействие аллельных генов. Вы знаете, что гены, контролирующие альтернативные формы проявления определенного признака, называются аллельными. Известно несколько типов взаимодействия аллельных генов.
Полное доминирование — взаимодействие, при котором доминантный аллель полностью подавляет проявление рецессивного аллеля. Вследствие этого доминантный признак проявляется как у гомозигот АА, так и у гетерозигот Аа. Полное доминирование — наиболее распространенный тип взаимодействия аллельных генов. Например, у гороха желтый цвет семян полностью доминирует над зеленым, а гладкая поверхность — над морщинистой. У человека карий цвет глаз полностью доминирует над голубым, резус-положительность — над резус-отрицательностью, праворукость — над леворукостью, шестипалость — над пятипалостью (нормальным количеством пальцев рук) и т. п.
Некоторые аллельные гены взаимодействуют по типу неполного доминирования. В этом случае доминантный признак проявляется только при наличии двух доминантных аллелей в генотипе (АА). Если же организм гетерозиготен (Аа), т. е. имеет лишь один доминантный аллель, в фенотипе проявляется признак, промежуточный между доминантным и рецессивным.
Неполное доминирование наблюдается, например, при наследовании окраски плодов 1 земляники. У этого растения гомозиготные особи имеют либо красные (АА), либо белые (аа) плоды (рис. 32.1). Если скрестить такие растения, у всех гибридов первого поколения проявится промежуточный признак — розовая окраска плодов.
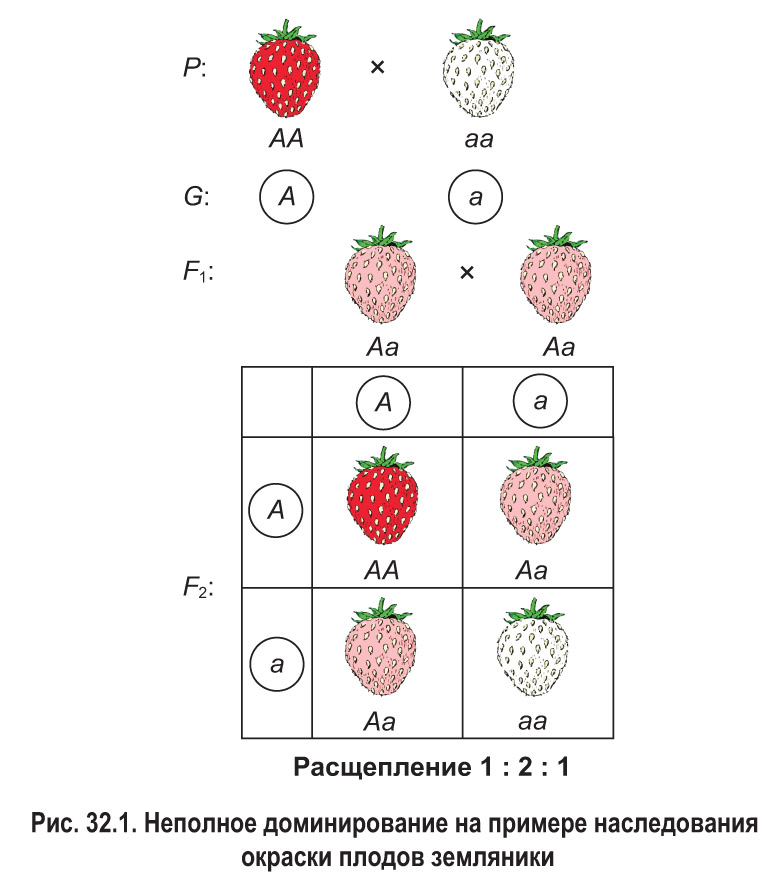
При неполном доминировании признак гетерозиготных особей отличается от признаков обеих гомозигот. Из-за этого во втором поколении наблюдается расщепление по фенотипу не 3 : 1, как при полном доминировании, а 1 : 2 : 1. Таким образом, в случае неполного доминирования расщепление по фенотипу совпадает с расщеплением по генотипу. В нашем примере одна часть гибридов второго поколения наследует красный цвет плодов (АА), две части — розовый (Аа) и одна часть — белый (аа).
По типу неполного доминирования наследуются и другие признаки живых организмов. Некоторые из них представлены в таблице 32.1 (не для запоминания).
Таблица 32.1. Некоторые признаки, при наследовании которых наблюдается неполное доминирование
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2.
Разная жизнеспособность зигот может быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими половой зрелости, называются летальными, более 50 % —сублетальными (полулетальными) и менее 50 % — субвитальными.
Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур.
Почти половина голых цыплят гибнет в последние 2—3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32—35 °С.
Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4—5 месяцев у голых цыплят вырастает редкое оперение, и они уже в состоянии переносить довольно низкие температуры. В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть:
В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей.
Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной.
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии. Такие летальные гены оказывают отрицательное воздействие в гомозиготе (рецессивной), в гетерозиготе они безвредны.
Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние.
В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2—4-й день после рождения от уродства прямой кишки — отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца.
Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина — гетерозиготной (LI), несущей рецессивный задаток (0 летального гена.
При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли.
Летальные гены примеры
Летальные гены – мутационные гены, вызывающие гибель особи до достижения ею половозрелого возраста. Бывают доминантными, рецессивными, сцепленные с полом.
Обычно проявляют своё действие в гомозиготном состоянии, в гетерозиготном состоянии понижают жизнеспособность. Пенетрантность – способность гена проявиться фенотипически, выражается в % и бывает полный и неполный.
Полный – у всех особей популяции, имеющих данный ген, он проявляется в виде признака. Неполный – у некоторых особей ген имеется, но внешне себя не проявляет. Экспрессивность – степень проявления признака, т.е. один и тот же признак у разных особей выражен с разной интенсивностью.
При наличии разных летальных генов организмы гибнут на различных стадиях развития.
Как правило, летальное действие таких генов рецессивно, т.е. проявляется лишь тогда, когда они находятся в гомозиготном состоянии.
При возникновении мутаций с доминирующим летальным действием организм погибает, не давая потомства.
Однако отмечены случаи, когда летальные гены, вызывая в гетерозиготном состоянии видимые изменения, становятся даже полезными с хозяйственной точки зрения. Так, среди овец каракульской породы встречаются животные с красивой серебристо-серой окраской шкурки, оцениваемой дороже, чем обычный черный каракуль, При скрещивании серых овец с серыми баранами оказалось, что они всегда гетерозиготны.
При изучении причин этого явления было установлено, что среди полученных в результате такого скрещивания серых ягнят примерно 1/3, или около 25% всего приплода, заболевала хроническим тимпанитом и погибала. Причиной заболевания служили нарушения в деятельности парасимпатической нервной системы. Когда же скрещивали серых баранов с черными овцами или серых овец с черными баранами, в потомстве рождалось 50% серых и 50% черных ягнят, причем серые ягнята не заболевали.
Оказалось, что в гомозиготном состоянии ген, вызывавший развитие серой окраски, обладал рецессивным летальным действием.
У лисиц встречается ценная окраска меха — платиновая, обусловленная доминантным геном, вызывающим в гомозиготном состоянии гибель зародышей на ранней стадии развития.
В таких случаях соотношение фенотипов во втором поколении меняется, так как из трех доминантных форм одна (гомозиготная по летальному гену) погибает, в результате чего соотношение фенотипов становится 2:1.
Аналогичное расщепление отмечено и у зеркального карпа, у которого наблюдалось наследование недоразвития чешуи: она сохранилась только на средней линии тела, остальная часть его голая, поэтому его называют линейным.
При скрещивании между собой линейных карпов потомство всегда расщепляется в соотношении 2 линейных: 1 с нормальной чешуей, так как зародыши, гомозиготные по гену, вызывающему недоразвитие чешуи, погибают на ранних стадиях развития.
У таких кур теплоизоляция недостаточна, и они страдают от охлаждения. Для компенсации потери тепла у них появляется ряд структурных и физиологических адаптаций, но эти адаптации малоэффектны и среди таких кур высока смертность.
Воздействие летального гена ясно видно на примере наследования окраски шерсти у мышей. У диких мышей шерсть обычно серая, типа агути; но у некоторых мышей шерсть желтая. При скрещиваниях между желтыми мышами в потомстве получаются как желтые мыши, так и агути в отношении 2:1.
Единственное возможное объяснение таких результатов состоит в том, что желтая окраска шерсти доминирует над агути и что все желтые мыши гетерозиготны. Атипичное менделевское отношение объясняется гибелью гомозиготных желтых мышей до рождения. При вскрытии беременных желтых мышей, скрещенных с желтыми же мышами, в их матках были обнаружены мертвые желтые мышата. Если же скрещивались желтые мыши и агути, то в матках беременных самок не оказывалось мертвых желтых мышат, поскольку при таком скрещивании не может быть потомства, гомозиготного по гену желтой шерсти.
Летальные гены и характер их наследования
Лета́льные алле́ли (также летали) — аллели, как правило, рецессивные, фенотипический эффект которых вызывает гибель организма при определённых условиях или на определённых этапах развития. Чаще всего гибель из-за леталей происходит на эмбриональных стадиях развития, но существуют летали, вызывающие гибель например, при окукливании личинки дрозофилы.
Летальные аллели возникают в результате т. н. летальныхмутаций — летальность таких мутаций говорит о том, что данный ген ответственен за какую-либо жизненно необходимую функцию[1].
Летальными называются аллели, носители которых погибают из-за нарушений развития или заболеваний, связанных с работой данного гена.
Между летальными аллелями и аллелями, вызывающими наследственные болезни, есть все переходы. Например, больные хореей Хантингтона (аутосомно-доминантный признак) обычно умирают в течение 15—20 лет после начала заболевания от осложнений, и в некоторых источниках предлагается считать этот ген летальным.
Сублетальными, или полулетальными называются аллели, эффект гибельности которых част, но не обязателен (то есть переходные между летальными аллелями и аллелями, вызывающими наследственные болезни), условно летальными называют мутации, при которых организм, несущий такие мутации, может жить в предельно узком диапазоне условий, например мутации ауксотрофности у микроорганизмов (неспособность расти на питательных средах без определённых жизненно необходимых веществ из-за утраты способности их синтезировать), субстратнозависимые мутации (неспособность использовать некоторые вещества в качестве источника углерода и энергии) и температурнозависимые мутации (способность жить только в узком диапазоне температур — например некоторые мутанты дрозофилы не способны жить при температуре выше 25 оС)
Рассмотрение гибридологического и генеалогического методов генетических исследований. Изучение признаков, детерминируемых летальными генами у разных видов сельскохозяйственных животных. Описание пенетрантности и экспрессивности при наследовании аномалий.
| Рубрика | Сельское, лесное хозяйство и землепользование |
| Вид | контрольная работа |
| Язык | русский |
| Дата добавления | 30.04.2014 |
| Размер файла | 95,3 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
МИНИСТЕРСТВО СЕЛЬСКОГО ХОЗЯЙСТВА РФ
Федеральное государственное бюджетное образовательное учреждение высшего профессионального образования
Пермская государственная сельскохозяйственная академия имени академика Д.Н. Прянишникова
Кафедра: кормление и разведения сельскохозяйственных животных
по дисциплине: генетика
Выполнила студентка 1 курса
Каменских Ксения Алексеевна
Проверил: старший преподаватель
Пьянкова Светлана Юрьевна
1. Введение
гибридологический генетический аномалия летальный
Генетика представляет собой одну из основных, наиболее увлекательных и вместе с тем сложных дисциплин современного естествознания. Место генетики среди биологических наук и особый интерес к ней определяются тем, что она изучает основные свойства организмов, а именно наследственность и изменчивость.
В результате многочисленных - блестящих по своему замыслу и тончайших по исполнению - экспериментов в области молекулярной генетики современная биология обогатилась двумя фундаментальными открытиями, которые уже нашли широкое отражение в генетике человека, а частично и выполнены на клетках человека.
Это показывает неразрывную связь успехов генетики человека с успехами современной биологии, которая все больше и больше становится связана с генетикой.
Первое - это возможность работать с изолированными генами. Она получена благодаря выделению гена в чистом виде и синтезу его. Значение этого открытия трудно переоценить. Важно подчеркнуть, что для синтеза гена применяют разные методы, т.е. уже имеется выбор, когда речь пойдет о таком сложном механизме как человек.
Второе достижение - это доказательство включения чужеродной информации в геном, а также функционирования его в клетках высших животных и человека. Материалы для этого открытия накапливались из разных экспериментальных подходов.
Прежде всего, это многочисленные исследования в области вирусогенетической теории возникновения злокачественных опухолей, включая обнаружение синтеза ДНК на РНКматрице. Кроме того, стимулированные идеей генетической инженерии опыты с профаговой трансдукцией подтвердили возможность функционирования генов простых организмов в клетках млекопитающих, включая клетки человека. Без преувеличения можно сказать, что, наряду с молекулярной генетикой, генетика человека относится к наиболее прогрессирующим разделам генетики в целом. Ее исследования простираются от биохимического до популяционного, с включением клеточного и организменного уровней.
2. Методы генетических исследований: гибридологический, генеалогический, популяционный, феногенетический, рекомбинационный, мутационный, цитогенетический, статистический
Для познания закономерностей наследования признаков и их изменчивости генетика использует ряд методов. Основным методом является гибридологический. При этом методе для выявления закономерностей наследования того или иного признака проводится скрещивание особей, различающихся по этому признаку, и изучается полученное потомство в первом и последующих поколениях. Гибридологический метод впервые в своих исследованиях удачно применил Г. Мендель.
Генеалогический метод является одним из вариантов гибридологического. Наследование признака при этом изучают путем анализа передачи его потомству в целых семьях или родственных группах животных, для чего составляют родословные на несколько поколений предков, отдельных особей и целых семей. Генеалогический метод имеет большое значение при изучении наследственности человека и медленно плодящихся животных, к которым обычный гибридологический метод или не применим, или требует продолжительного времени для получения результатов опыта.
Частным случаем гибридологического метода является рекомбинационный метод, основанный на явлении кроссинговера. Его широко используют для составления генетических карт, создания рекомбинантных молекул ДНК, содержащих генетические системы различных организмов.
Цитологический метод применяют при изучении наследственности на уровне клетки и хромосом. Установлено, что многие дефекты и нарушения в организме связаны с изменениями в числе и структуре хромосом. Поэтому при диагностике некоторых наследственных заболеваний человека и животных широко используется цитологический метод.
Биохимический метод используется в генетике для более глубокого анализа нарушений в обмене веществ и их строении. Этот метод используется при манипуляциях на уровне ДНК в генной инженерии.
Популяционно-статистический метод используется для изучения явлений наследственности в популяциях. Этот метод дает возможность установить частоту доминантных и рецессивных аллелей, определяющих тот или иной признак, частоту доминантных и рецессивных гомозигот и гетерозигот, динамику генетической структуры популяций под влиянием мутаций, изоляции и отбора. Широко используется в современной селекции животных.
Феногенетический метод применяется для установления степени влияния генов и факторов внешней среды на развитие признаков организма. При использовании этого метода изучаются особи с разной наследственностью или находящиеся в разных условиях среды.
Мутационный метод (мутагенез) позволяет установить характер влияния мутагенных факторов на генетический аппарат клетки, ДНК, хромосомы, на изменения признаков или свойств. Мутагенез используют в селекции сельскохозяйственных растений, в микробиологии для создания новых штаммов бактерий. Он нашел применение в селекции тутового шелкопряда.
3. Летальные гены и их действие. Назовите признаки, детерминируемые летальными генами у разных видов сельскохозяйственных животных и покажите их на схемах скрещиваний особенности их наследования
Изменение расщепления по фенотипу в отношении 3:1 во втором поколении моногибридного скрещивания бывает связано с разной жизнеспособностью зигот Ф2. Разная жизнеспособность зигот может быть обусловлена наличием летальных генов. Летальным называется ген, вызывающий нарушения в развитии организма, что приводит его к гибели или уродству.
Изучение врожденных аномалий показало, что при разных летальных генах гибель особей бывает различной и может происходить на разных стадиях развития.
По классификации, предложенной Розенбауэром (1969), гены, вызывающие гибель 100 % особей до достижения ими пЪловой зрелости, называются летальными, более 50 % --сублетальными (полулетальными) и менее 50 % -- субвитальными. Однако следует отметить, что разделение это до некоторой степени условно и иногда не имеет четких границ. Примером может служить сцепленная с полом голость у кур. Почти половина голых цыплят гибнет в последние 2--3 дня инкубации. Из числа вылупившихся около половины цыплят гибнет до 6-недельного возраста, если их выращивают при температуре 32--35 "С. Но если температура в брудерах будет повышена на 5,5 °С, то погибнет значительно меньше голых цыплят. В 4--5 мес у голых цыплят вырастает редкое оперение и они уже в состоянии переносить довольно низкие температуры.
В естественных условиях эта мутация, по-видимому, будет летальной и приведет к 100%-ной гибели птиц. Приведенный пример показывает, что характер проявления полулетального гена в значительной мере может зависеть от условий среды.
Летальные гены могут быть доминантными и рецессивными. В числе первых летальных факторов был открыт аллель, обусловливающий желтую окраску мышей. Ген желтой окраски является доминантным (Y). Его рецессивный аллель (у) в гомозиготном состоянии обусловливает появление черной окраски. Скрещивание желтых мышей между собой давало в потомстве две части особей желтых и одну часть черных, т. е. получалось расщепление 2:1, а не 3:1, как следовало из правила Менделя. Оказалось, что все взрослые мыши гетерозиготны (Yy). При скрещивании между собой они должны были дать одну часть гомозиготного потомства по желтой окраске (IT), но оно погибает еще в эмбриональном периоде, две части гетерозигот (Yy) будут желтыми и одна часть гомозигот по рецессивному признаку (уу) будет черной. Схема скрещивания выглядит так:
Таким же образом наследуется серая окраска шерсти у каракульских овец (сокольских, малич и др.), платиновая окраска у лисиц, распределение чешуи у линейных карпов и т. д.
Летальные гены в большинстве случаев рецессивны и поэтому долгое время могут находиться в скрытом состоянии.
Совершенно здоровое и нормальное по фенотипу животное может быть носителем летального гена, действие которого обнаруживается только при переходе в гомозиготное состояние. В гомозиготное состояние летальные гены чаще всего переходят при родственном спаривании. В практике животноводства при разведении лошадей был случай гибели 25 жеребят на 2--4-й день после рождения от уродства прямой кишки -- отсутствия заднепроходного отверстия (Atresia ani). Оказалось, что все жеребцы и кобылы, от которых рождались такие ненормальные жеребята, происходили от одного жеребца. Он был гетерозиготен по летальному гену (LI). Вначале этот жеребец при скрещивании с нормальными кобылами (LL) дал потомство, по фенотипу нормальное, но по генотипу половина потомков была благополучной (LL), a половина -- гетерозиготной (LI), несущей рецессивный задаток (0 летального гена. При родственном спаривании гетерозиготных животных (Ы х Ы) появилась часть жеребят, гомозиготных по летальному гену (II), с уродством прямой кишки. Они все погибли.
4. Пенетрантность и экспрессивность при наследовании аномалий
Пенетрантность стерильности колеблется от 0,42 до 0,62.
Как уже известно, существует понятие экспрессивности генов. В качестве примеров разной экспрессивности генов можно привести следующие. Так, адактилия у крупного рогатого скота варьирует от частичного до полного отсутствия фаланг конечностей. При исследовании пупочных грыж у телят мы обнаружили значительные различия в диаметре грыжевого отверстия. При изучении нарушения координации движений у телят черно-пестрой и симментальской пород мы наблюдали заметную разницу во внешнем проявлении аномалии -- от дрожания головы и покачивающейся ходьбы до резко выраженной несогласованности движений головы и конечностей при попытке животных встать. Патологоанатомическими и гистологическими исследованиями установлена различная степень гипоплазии мозжечка у телят, что сочеталось со степенью фенотипического проявления аномалии.
У сельскохозяйственных животных известны десятки аномалий, возникновение которых связано с рецессивными или доминантными мутациями генов. Эти аномалии встречаются в отдельных популяциях с разной частотой, что зависит от скорости мутационного процесса, системы разведения животных и др. Знание конкретных форм врожденных аномалий у животных каждого вида, а также частоты их проявления в отдельных породах необходимо ветеринарным специалистам для селекционной профилактики распространения генетической патологии.
Аномалии у крупного рогатого скота. Биологические особенности данного вида животных -- малоплодие и относительная позднеспелость. Корова обычно приносит одного теленка, который достигает половой и физиологической зрелости только к 1,5 года, так что период между первым отелом матери и дочери в среднем составляет 5 лет. Вследствие этого появление в стаде аномального приплода может существенно снижать уровень воспроизводства и интенсивность племенного отбора поголовья. У крупного рогатого скота изучен широкий спектр врожденных аномалий, детерминированных летальными, полулетальными и субвитальными генами. 46 аномалий включено в Международный список летальных дефектов под шифром А (табл.).
Разумное собаководство осуществляется при комбинировании двух основных типов разведения — ИНБРИДИНГА и ЛАЙНБРИДИНГА, но время от времени необходим и разумный АУТКРОССИНГ. В среднем, потомство при всех типах разведения будет практически равным, но там, где применялся инбридинг, особенно в чрезмерной степени, потомство будет или значительно лучше, или много хуже.
Следует также помнить, что даже самый выдающийся племенной кобель не будет в генетическом смысле одинаково хорошо подходить для всех сук.
Гены старения
Гены старения Возраст — мерзкая вещь, и с каждым годом она становится все хуже. Диана Купер Люди не хотят жить вечно. Люди просто не хотят умирать. Станислав Лем — Бессмертные клетки существуют — Запрограммированная гибель — Часовой механизм старения — Болезни
Гены и поведение
Гены и поведение Факты, добытые нейробиологами, говорят о материальной, нейрологической природе психики. Но для того, чтобы душа во всех ее проявлениях могла эволюционировать, этого, строго говоря, еще недостаточно. Эволюционировать могут не все признаки, а только
Новые гены?
Новые гены? Активность генов может меняться в ходе эволюции не только путем изменений сайтов связывания ТФ, работы самих ТФ или регуляторных РНК, но и в результате дупликации генов. При прочих равных два одинаковых гена произведут больше продукта (то есть информационной
Рецессивные гены
Рецессивные гены РЕЦЕССИВНЫЙ ГЕН (т.е. признак, им определяемый) МОЖЕТ НЕ ПРОЯВЛЯТЬСЯ У ОДНОГО ИЛИ МНОГИХ ПОКОЛЕНИЙ пока не встретятся два идентичных рецессивных гена от каждого из родителей (внезапное проявление такого признака у потомков не следует путать с
Доминантные гены
Доминантные гены Присутствие доминантного гена всегда явно и внешне проявляется соответствующим признаком. Поэтому доминантные гены, несущие нежелательный признак, представляют для селекционера значительно меньшую опасность, чем рецессивные, так как их присутствие
Рецессивные гены
Рецессивные гены РЕЦЕССИВНЫЙ ГЕН (т. е. признак, им определяемый) МОЖЕТ НЕ ПРОЯВЛЯТЬСЯ У ОДНОГО ИЛИ МНОГИХ ПОКОЛЕНИЙ пока не встретятся два идентичных рецессивных гена от каждого из родителей (внезапное проявление такого признака у потомков не следует путать с
Доминантные гены
Доминантные гены Присутствие доминантного гена всегда явно и внешне проявляется соответствующим признаком. Поэтому доминантные гены, несущие нежелательный признак, представляют для селекционера значительно меньшую опасность, чем рецессивные, так как их присутствие
Летальные гены
Гены спортсмена
Гены спортсмена Женщины любят его за его внешность, а мужчинам нравится его умение забивать мячи. Женщины восхищаются его прической и соблазнительной улыбкой, мужчины уважают его за броски с фланга. Речь идет о двух персонах. После всего, что мы узнали о генетическом
Гены Моцарта
Гены Моцарта О детстве Моцарта не так много точных сведений, куда больше – предположений. Как все происходило у этого маленького гения? Сестра Наннерль, старше его на пять лет, захотела узнать побольше о своем младшем брате. Какой он был на самом деле в раннем детстве? В
ГЕНиальные гены
ГЕНиальные гены А теперь совсем другой вопрос: почему это так интересно? Что можно такого узнать, исследовав череп Моцарта? Ну, допустим, его взвесили и измерили. По результатам анализа костей можно выяснить, что ел владелец этого черепа. И это тоже уже сделали. Этот череп
Гены-преступники
Гены-преступники После того как в гангстерской саге Фрэнсиса Форда Копполы умирает крестный отец Дон Вито Корлеоне (Марлон Брандо) у мужчин клана появляется новый крестный отец – его сын Майкл Корлеоне (Аль Пачино). И почему сын должен быть таким же гениальным
Гены Открытие Арендта подводит нас еще к одному вопросу. Одно дело, что у глаз разных животных есть общие части, но как получилось, что такие непохожие друг на друга глаза, как у червей, мух и мышей, родственны друг другу? Чтобы ответить на этот вопрос, обратимся к
Читайте также: