
История открытия хромосом кратко
Обновлено: 26.07.2024
Только благодаря этим организмам впервые было показано, что дезоксирибонуклеиновая кислота, рибонуклеиновая кислота и белок - универсальные детерминанты генетического поведения. Стремительность дальнейшего прогресса в этой области и убедительность полученных результатов стали реальными благодаря особым биологическим свойствам микроорганизмов, которые позволяли проводить манипуляции, необходимые для анализа генетических структур. Аналогичные аналитические исследования более сложных генетических систем тогда были невозможны, поэтому на животных и растения этот прогресс не распространялся.
Содержание работы
Введение…………………………. 3
История открытия хромосом………………………………………………4
Хромосомы, их строение, видовая специфичность, кариотип………….5
Участки хромосом…………………………………………………………6
Хромосомные аномалии…………………………………………………. 8
Денверская классификация хромосом……………………………………9
Заключение………………………………………………………………..11
Список литературы……………………………………………………….12
Файлы: 1 файл
строение и функци хромосом 2.docx
- Введение…………………………. . . 3
- История открытия хромосом………………………………………………4
- Хромосомы, их строение, видовая специфичность, кариотип………….5
- Участки хромосом………………………………………………………… 6
- Хромосомные аномалии…………………………………………………. 8
- Денверская классификация хромосом……………………………………9
- Заключение…………………………………………………… …………..11
- Список литературы…………………………………………………… ….12
Систематическое изучение наследственности начиналось со сложных в генетическом отношении объектов - растений и животных. Благодаря этим ранним исследованиям была сформулирована концепция неделимого гена как функциональной единицы наследственности и принято положение, что перенос генов от одного поколения к другому подвержен действию разных случайных факторов. Однако до понимания химической природы генов и механизма их функционирования было еще далеко. Исследование генетических молекул и тонких механизмов регуляции наследственности стало возможным лишь тогда, когда в качестве экспериментальных моделей начали использоваться бактерии и вирусы, о существовании которых первые генетики даже не подозревали. Только благодаря этим организмам впервые было показано, что дезоксирибонуклеиновая кислота, рибонуклеиновая кислота и белок - универсальные детерминанты генетического поведения. Стремительность дальнейшего прогресса в этой области и убедительность полученных результатов стали реальными благодаря особым биологическим свойствам микроорганизмов, которые позволяли проводить манипуляции, необходимые для анализа генетических структур. Аналогичные аналитические исследования более сложных генетических систем тогда были невозможны, поэтому на животных и растения этот прогресс не распространялся. Развитие технологии рекомбинантных ДНК разрушило труднопреодолимые технические и концептуальные барьеры на пути расшифровки и понимания сложных генетических систем. Неудивительно, что наши взгляды на структуру и функцию генов значительно изменились, а новое мышление в свою очередь радикально изменило перспективы биологии.
Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидные структуры в ядре эукариотической клетки, в которых сосредоточена большая часть наследственной информации и которые предназначены для её хранения, реализации и передачи. Хромосомы чётко различимы в световом микроскопе только в период митотического или мейотического деления клетки. Набор всех хромосом клетки, называемый кариотипом, является видоспецифичным признаком, для которого характерен относительно низкий уровень индивидуальной изменчивости.
История открытия хромосом
Хромосомы, их строение, видовая специфичность, кариотип
Хромосомы - наиболее важные компоненты ядра. Они играют ведущую роль в явлениях наследственности. Хромосомы хорошо видны под микроскопом в момент деления клетки. Хромосомы ядра неделящейся клетки не видны, поскольку они деконденсации хромосом, тем активнее протекают метаболические процессы в самом ядре.
Морфологические хромосомы растений чаще всего имеют нитевидную или палочкообразную форму. Большинство хромосом разделено первичной перетяжкой на два плеча. Под микроскопом первичная перетяжка представлена светлой (неокрашенной) зоной, получившее название центромеры, которые играют основную роль в перемещении хромосом строго определении ядра. Центромера занимает на каждой из хромосом строго определенного место. По положению центромеры хромосомы делят на метацентрические (приблизительно равноплечие), субметацентрические (неравноплечие) и акроцентрические (головчатые), у которых центромера сдвинута к одному из концов.
У некоторых хромосом имеется и вторичная перетяжка. Она, как правило, располагается у дистального конца хромосомы и отделяет небольшой ее участок, носящий название спутника. Вторичная перетяжка не участвует в движении хромосом при деление ядра. Она получила название ядрышкового организатора, поскольку в месте ее локализация происходит образование ядрышка. Концевые участки хромосомы называют теломерными. Они препятствуют ее соединению с другими хромосомами.
Формы хромосом на стадии метафазы (схема):1,5 - равноплечие; 2,3 - неравноплечие; 4 – головчатые
Каждому из населяющих нашу планету видов растений и животных свойственно строгое число хромосом, обозначаемое 2n (диплоидный набор). В половых клетках число хромосом в два раза меньше и равно n (гаплоидный набор). В соматических клетках организма каждая хромосома имеет пару, идентичную как морфологически, так и генетически (гомологичные хромосомы).
Исключение из этого правила составляют половые хромосомы у
гетерогаметных особей. Специфический для определенного вида по числу и структуре набор хромосом получил название кариотипа.
Графическое изображение кариотипа, показывающие его структурные особенности, называется идиограммой. В последние годы получил распространение метод дифференциального окрашивание хромосом. При этом на каждого из хромосом прокрашиваются специфические, характерные для нее полосы (бэнды), что значительно облегчает идентификацию отдельных хромосом кариотипа.
Хромосомы, определяющие пол особи, называют половыми хромосомами, а все остальные - аутосомами. Внутренне строение хромосом чрезвычайно сложно. По химическому составу они на 40% состоят из ДНК и на 60% из белков, в среднем около 60% из которых приходится гистоны. Строение метафазной хромосомы при исследовании с помощью светового микроскопа представляет следующим образом. Каждая хромосома состоит из двух хроматид, спирально закрученными и располагающихся параллельно оси хромосомы. Для прокрашивающихся в интерфазном яде участков хромосом используют термин "хромонема" - красящая нить. Утолщения на хромонемах получили название хромомер. Особенность вышеописанного строения хромосом зависит от уровня меняется при переходе от интерфазного состояния хромосом к метафазному.
Первый, получивший название нуклеосомного, определяет скручивание ДНК по поверхности гистоновой сердцевины. Второй - объединение нескольких нуклеосом (до 10) в бусину - называется нуклеомерный. Третий уровень - объединение скрепками из негистоновых белков фибрилл дезоксирибонуклеопротеида в петлевой домен, называемый хромомером. Четвертый - образование хромонем. Далее, по-видимому, хромонема укладывается в виде спирали в хроматиде, хотя весьма вероятно, что это еще один уровень - "петлистых структур".
Центромера (первичная перетяжка)- это место соединения двух хроматид; к центромере присоединяются нити веретена деления. По сторонам от центромеры лежат плечи хромосомы.
В зависимости от места расположения центромеры хромосомы делят на:
палочковидные (акроцентрические) – имеется только одно плечо.
Вторичная перетяжка – ядрышковый организатор, содержит гены рРНК, имеется у одной – двух хромосом в геноме.
Теломеры – концевые участки хромосом, содержащие до 10 тысяч пар нуклеотидов с повторяющейся последовательностью ТТАГГГ. Теломеры не содержат генов, они защищают концы хромосом он действия нуклеаз – ферментов, разрушающих ДНК. Обеспечивают прикрепление концов хромосом изнутри к ядерной оболочке. Защищают гены от концевой недорепликации.
человека(серые) и их
Схема расположения теломер на хромосоме.
Анеуплоидия. При анеуплоидии происходит потеря или приобретение хромосомы одной пары. В случае утраты хромосомы мутантов называют моносомиками, в случае приобретения хромосомы — трисомиками. Анеуплоидия у людей часто не совместима с жизнью или приводит к тяжёлым патологиям. Например, трисомия по хромосоме 21 у человека приводит к развитию синдрома Дауна.
Полиплоидия. Изменение числа хромосом, кратное гаплоидному набору хромосом (n), называется полиплоидией. Полиплоидия широко и неравномерно распространена в природе. Известны полиплоидные эукариотические микроорганизмы — грибы и водоросли, часто встречаются полиплоиды среди цветковых, но не среди голосемянных растений. Полиплоидия клеток всего организма у многоклеточных животных редка, хотя у них часто встречается эндополиплоидия некоторых дифференцированных тканей, например, печени у млекопитающих, а также тканей кишечника, слюнных желёз, мальпигиевых сосудов ряда насекомых.
Хромосомные перестройки (хромосомные аберрации) — это мутации, нарушающие структуру хромосом. Они могут возникнуть в соматических и зародышевых клетках спонтанно или в результате внешних воздействий (ионизирующее излучение, химические мутагены, вирусная инфекция и др.). В результате хромосомной перестройки может быть утрачен или, наоборот, удвоен фрагмент хромосомы (делеция и дупликация, соответственно); участок хромосомы может быть перенесён на другую хромосому (транслокация) или он может изменить свою ориентацию в составе хромосомы на 180° (инверсия). Существуют и другие хромосомные перестройки.
Денверская классификация хромосом
Классификация и номенклатура равномерно окрашенных хромосом человека впервые были приняты на международном совещании в 1960 году в г. Денвере, в дальнейшем несколько измененные и дополненные(Лондон, 1963 и Чикаго,1966).Согласно Денверовской классификации все хромосомы человека разделены на 7 групп, расположенных в порядке уменьшения их длины и с учетом центриольного индекса(отношение длины короткого плеча к длине всей хромосомы, выраженное в процентах). Группы обозначаются буквами английского алфавита от А до G. Все пары хромосом принято нумеровать арабскими цифрами. Характеристика групп представлена в табл.4
Хромосомы представляют собой нуклеопротеидные структуры эукариотической клетки, в которых хранится большая часть наследственной информации. Благодаря своей способности к самовоспроизведению, именно хромосомы обеспечивают генетическую связь поколений. Хромосомы образуются из длинной молекулы ДНК, в которой содержится линейная группа множества генов, и вся генетическая информация будь-то о человеке, животном, растении или любом другом живом существе.
Морфология хромосом связана с уровнем их спирализации. Так, если во время стадии интерфазы хромосомы максимально развернуты, то с началом деления хромосомы активно спирализуются и укорачиваются. Своего максимального укорочения и спирализации они достигают во время стадии метафазы, когда происходит формирование новых структур. Эта фаза наиболее удобна для изучения свойств хромосом, их морфологических характеристик.
История открытия
Хромосомная теория наследственности
Разумеется, история изучения хромосом не закончилась на их открытии, так в 1901-1902 годах американские ученые Уилсон и Сатон, причем независимо друг от друга, обратили внимание на сходство в поведении хромосом и менделевских факторов наследственности – генов. В результате ученые пришли к заключению, что гены находятся в хромосомах и именно посредством их из поколения в поколения, от родителей к детям передается генетическая информация.
В 1915-1920 годам участие хромосом в передаче генов было доказано на практике в целой серии опытов, сделанных американским ученым Морганом и сотрудниками его лаборатории. Им удалось локализировать в хромосомах мухи-дрозофилы несколько сот наследственных генов и создать генетические карты хромосом. На основе этих данных была создана хромосомная теория наследственности.
Строение
Строение хромосом разнится в зависимости от вида, так метафазная хромосома (образующаяся в стадии метафазе при митозном делении клетки) состоит из двух продольных нитей – хроматид, которые соединяются в точке, именуемой центромерой. Центромера – это участок хромосомы, который отвечает за расхождение сестринских хроматид в дочерние клетки. Она же делит хромосому на две части, названные коротким и долгим плечом, она же отвечает за деление хромосомы, так как именно в ней содержится специальное вещество – кинетохор, к которому крепятся структуры веретена деления.
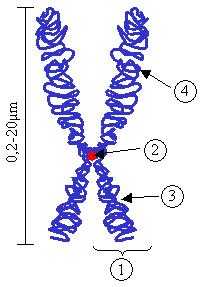
Тут на картинке показано наглядное строение хромосомы: 1. хроматиды, 2. центромера, 3. короткое плечо хроматид, 4. длинное плечо хроматид. На концах хроматид располагаются теломеры, специальные элементы, которые защищают хромосому от повреждений и препятствуют слипанию фрагментов.
Формы и виды
Размеры хромосом растений и животных значительно различаются: от долей микрона до десятков микрон. Средние длины метафазных хромосом человека лежат в диапазоне от 1,5 до 10 микрон. В зависимости от вида хромосомы отличаются и ее способности к окрашиванию. В зависимости от расположения центромеры различают такие формы хромосом:
- Метацентрические хромосомы, для которых характерно срединное расположение центромеры.
- Субметацентрические, для них характерно неравномерное расположение хроматид, когда одно плечо более длинное, а второе более короткое.
- Акроцентрические или палочковидные. У них центромера расположена практически в самом конце хромосомы.
Функции
Основные функции хромосом, как для животных, так и для растений и вообще всех живых существ – передача наследственной, генетической информации от родителей к детям.
Набор хромосом
Значение хромосом столь велико, что их количество в клетках, а также особенности каждой хромосомы определяют характерный признак того или иного биологического вида. Так, например, у мухи-дрозофилы в наличии 8 хромосом, у обезьян – 48, а хромосомный набор человека составляет 46 хромосом.
В природе существует два основных типа набора хромосом: одиночный или гаплоидный (содержится в половых клетках) и двойной или диплоидный. Диплоидный набор хромосом имеет парную структуру, то есть вся совокупность хромосом состоит из хромосомных пар.
Хромосомный набор человека
Как мы уже написали выше, клетки человеческого организма содержат 46 хромосом, которые объединены в 23 пары. Все вместе они и составляют хромосомный набор человека. Первые 22 пары человеческих хромосом (их называют аутосомами) являются общими как для мужчин, так и для женщин, и лишь 23 пара – половых хромосом – разнится у разных полов, она же определяет половую принадлежность человека. Совокупность всех пар хромосом также называется кариотипом.
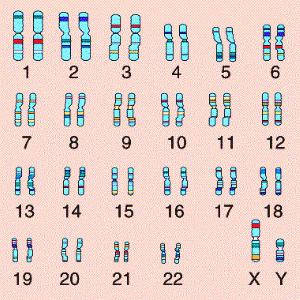
Такой вид имеет хромосомный набор человека, 22 пары двойных диплоидных хромосом содержат всю нашу наследственную информацию, и последняя пара различается, у мужчин она состоит из пары условных X и Y половых хромосом, в то время как у женщин в наличии две хромосомы Х.
Аналогичную структуру хромосомного набора имеют и все животные, только количество неполовых хромосом у каждого из них свое.
Генетические болезни, связанные с хромосомами
Нарушение в работе хромосом, или даже само их неправильно количество является причиной многих генетических заболеваний. Например, синдрома Дауна появляется из-за наличия лишней хромосомы в хромосомном наборе человека. А такие генетические болезни как дальтонизм, гемофилия вызваны сбоями в работе имеющихся хромосом.

Схема строения хромосомы в поздней профазе — метафазе митоза. 1 — хроматида; 2 — центромера; 3 — короткое плечо; 4 — длинное плечо.


Хромосо́мы (др.-греч. χρῶμα — цвет и σῶμα — тело) — нуклеопротеидные структуры в ядре эукариотической клетки (клетки, содержащей ядро), которые становятся легко заметными в определённых фазах клеточного цикла (во время митоза или мейоза). Хромосомы представляют собой высокую степень конденсации хроматина, постоянно присутствующего в клеточном ядре. Исходно термин был предложен для обозначения структур, выявляемых в эукариотических клетках, но в последние десятилетия всё чаще говорят о бактериальных хромосомах. В хромосомах сосредоточена большая часть наследственной информации.
Содержание
История открытия хромосом

Рисунок из книги В. Флемминга, изображающий разные стадии деления клеток эпителия саламандры (W. Flemming. Zellsubstanz, Kern und Zelltheilung. 1882 г.)
Сейчас сложно сказать, кто сделал первое описание и рисунок хромосом. В 1872 году швейцарский ботаник Карл фон Нэгили опубликовал работу, в которой изобразил некие тельца, возникающие на месте ядра во время деления клетки при образовании пыльцы у лилии (Lilium tigrinum) и традесканции (Tradescantia). Однако его рисунки не позволяют однозначно утверждать, что К. Нэгили видел именно хромосомы. В том же 1872 году ботаник Э. Руссов привёл свои изображения деления клеток при образовании спор у папоротника из рода ужовник (Ophioglossum) и пыльцы лилии (Lilium bulbiferum). На его иллюстрациях легко узнать отдельные хромосомы и стадии деления. Некоторые же исследователи полагают, что первыми увидел хромосомы немецкий ботаник Вильгельм Гофмайстер задолго до К. Нэгили и Э. Руссова, ещё в 1848—1849 годах. При этом ни К. Нэгили, ни Э. Руссов, ни тем более В. Гофмейстер не осознавали значения того, что видели.
В 1933 году за открытие роли хромосом в наследственности Т. Морган получил Нобелевскую премию по физиологии и медицине [1] .
Хромосомы эукариот
Основу хромосомы составляет линейная (не замкнутая в кольцо) макромолекула дезоксирибонуклеиновой кислоты (ДНК) значительной длины (например, в молекулах ДНК хромосом человека насчитывается от 50 до 245 миллионов пар азотистых оснований). В растянутом виде длина хромосомы человека может достигать 5 см. Помимо неё, в состав хромосомы входят пять специализированных белков — H1, H2A, H2B, H3 и H4 (так называемые гистоны) и ряд негистоновых белков. Последовательность аминокислот гистонов высококонсервативна и практически не различается в самых разных группах организмов.

В интерфазе хроматин не конденсирован, но и в это время его нити представляют собой комплекс из ДНК и белков. Макромолекула ДНК обвивает октамеры (структуры, состоящие из восьми белковых глобул) гистоновых белков H2A, H2B, H3 и H4, образуя структуры, названные нуклеосомами. В целом вся конструкция несколько напоминает бусы. Последовательность из таких нуклеосом, соединённых белком H1, называется нуклеофиламентом (nucleofilament), или нуклеосомной нитью, диаметром около 10 нм.
В ранней интерфазе (фаза G1) основу каждой из будущих хромосом составляет одна молекула ДНК. В фазе синтеза (S) молекулы ДНК вступают в процесс репликации и удваиваются. В поздней интерфазе (фаза G2) основа каждой из хромосом состоит из двух идентичных молекул ДНК, образовавшихся в результате репликации и соединённых между собой в районе центромерной последовательности.
Перед началом деления клеточного ядра хромосома, представленная на этот момент цепочкой нуклеосом, начинает спирализовываться, или упаковываться, образуя при помощи белка H1 более толстую хроматиновую нить, или хроматиду, (chromatin fiber) диаметром 30 нм. В результате дальнейшей спирализации диаметр хроматиды достигает ко времени метафазы 700 нм. Значительная толщина хромосомы (диаметр 1400 нм) на стадии метафазы позволяет, наконец, увидеть её в световой микроскоп. Конденсированная хромосома имеет вид буквы X (часто с неравными плечами), поскольку две хроматиды, возникшие в результате репликации, по-прежнему соединены между собой в районе центромеры (подробнее о судьбе хромосом при клеточном делении см. статьи митоз и мейоз).
Первичная перетяжка
Хромосомная перетяжка (X. п.), в которой локализуется центромера и которая делит хромосому на плечи.
Вторичные перетяжки
Морфологический признак, позволяющий идентифицировать отдельные хромосомы в наборе. От первичной перетяжки отличаются отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. У человека это 9, 13, 14, 15, 21 и 22 хромосомы.
Типы строения хромосом
Различают четыре типа строения хромосом:
- телоцентрические (палочковидные хромосомы с центромерой, расположенной на проксимальном конце);
- акроцентрические (палочковидные хромосомы с очень коротким, почти незаметным вторым плечом);
- субметацентрические (с плечами неравной длины, напоминающие по форме букву L);
- метацентрические (V-образные хромосомы, обладающие плечами равной длины).
Тип хромосом является постоянным для каждой гомологичной хромосомы и может быть постоянным у всех представителей одного вида или рода.
Спутники (сателлиты)
Сателлит — это округлое или удлинённое тельце, отделённое от основной части хромосомы тонкой хроматиновой нитью, по диаметру равный или несколько меньший хромосоме. Хромосомы, обладающие спутником принято обозначать SAT-хромосомами. Форма, величина спутника и связывающей его нити постоянны для каждой хромосомы.
Зона ядрышка
Зоны ядрышка (организаторы ядрышка) — специальные участки, с которыми связано появление некоторых вторичных перетяжек.
Хромонема
Хромонема — это спиральная структура, которую удаётся увидеть в декомпактизованных хромосомах через электронный микроскоп. Впервые наблюдалась Баранецким в 1880 году в хромосомах клеток пыльников традесканции, термин ввёл Вейдовский. Хромонема может состоять из двух, четырёх и более нитей, в зависимости от исследуемого объекта. Эти нити образуют спирали двух типов:
- паранемическую (элементы спирали легко разъединить);
- плектонемическую (нити плотно переплетаются).
Хромосомные перестройки
Нарушение структуры хромосом происходит в результате спонтанных или спровоцированных изменений (например, после облучения).
-
(точковые) мутации (изменения на молекулярном уровне);
- Аберрации (микроскопические изменения, различимые при помощи светового микроскопа):
Гигантские хромосомы
Такие хромосомы, для которых характерны огромные размеры, можно наблюдать в некоторых клетках на определённых стадиях клеточного цикла. Например, они обнаруживаются в клетках некоторых тканей личинок двукрылых насекомых (политенные хромосомы) и в ооцитах различных позвоночных и беспозвоночных (хромосомы типа ламповых щёток). Именно на препаратах гигантских хромосом удалось выявить признаки активности генов.
Политенные хромосомы
Впервые обнаружены Бальбиани в 1881-го, однако их цитогенетическая роль была выявлена Костовым, Пайнтером, Гейтцем и Бауером. Содержатся в клетках слюнных желёз, кишечника, трахей, жирового тела и мальпигиевых сосудов личинок двукрылых.
Хромосомы типа ламповых щёток
Бактериальные хромосомы
Прокариоты (археи и бактерии, в том числе митохондрии и пластиды, постоянно обитающие в клетках большинства эукариот) не имеют хромосом в собственном смысле этого слова. У большинства из них в клетке имеется только одна макромолекула ДНК, замкнутая в кольцо (эта структура получила название нуклеоид). У ряда бактерий обнаружены линейные (не замкнутые в кольцо) макромолекулы ДНК. Помимо нуклеоида или линейных макромолекул, ДНК может присутствовать в цитоплазме прокариотных клеток в виде небольших замкнутых в кольцо молекул ДНК, так называемых плазмид, содержащих обычно незначительное, по сравнению с бактериальной хромосомой, число генов. Состав плазмид может быть непостоянен, бактерии могут обмениваться плазмидами в ходе парасексуального процесса.
Имеются данные о наличии у бактерий белков, связанных с ДНК нуклеоида, но гистонов у них не обнаружено.
Хромосомы человека
В каждой ядросодержащей соматической клетке человека содержится 23 пары линейных хромосом, а также многочисленные копии митохондриальной ДНК. В нижеприведённой таблице показано число генов и оснований в хромосомах человека.

Обзор
Автор
Редактор
Обратите внимание!
Спонсоры конкурса: Лаборатория биотехнологических исследований 3D Bioprinting Solutions и Студия научной графики, анимации и моделирования Visual Science.

Нажмите на изображение, чтобы увеличить (откроется в отдельном окне).
Генетическая роль хромосом
Сейчас сложно сказать, кто сделал первое описание хромосом. В 1842 году швейцарский ботаник К. Нэгели (C. Nägeli) опубликовал работу, в которой изобразил некие тельца, возникающие на месте ядра во время деления клетки при образовании пыльцы у лилии и традесканции. Возможно, это и были первые рисунки хромосом. Первое (1873 год) подробное описание митоза у плоского червя Mesostoma ehrenbergii принадлежит, как считают, немецкому зоологу А. Шнайдеру (F.A. Schneider). Он описал не просто отдельные стадии митоза, которые видели и до него, а всю последовательность сложных изменений ядра: возникновение на его месте нитевидных телец, их расхождение в противоположные стороны и формирование новых ядер в дочерних клетках. Другой тип деления — мейоз — впервые подробно описал Э. ван Бенеден (E. van Beneden, Бельгия) в 1883 году, наблюдая за образованием гамет у аскариды. Он обнаружил, что в мейозе число хромосом уменьшается вдвое, а при оплодотворении восстанавливается, и, несмотря на различие в размерах, мужская и женская гаметы привносят в зиготу равное число хромосом.
В 1902 году Т. Бовери (T. Boveri, Германия) и в 1902–1903 годах У. Сеттон (W. Sutton, США) независимо друг от друга выдвинули гипотезу о генетической роли хромосом. Т. Бовери обнаружил, что зародыш морского ежа может нормально развиваться только при наличии у него хотя бы одного, но полного набора хромосом. Также он установил, что разные хромосомы не идентичны друг другу по своему составу. У. Сеттон же изучал гаметогенез у саранчи и понял, что поведение хромосом в мейозе и при оплодотворении полностью объясняет закономерности расхождения менделевских факторов и образования их новых комбинаций.
Одним из важных свидетельств генетической функции хромосом было доказательство роли половых хромосом в определении пола. К. Бриджес в 1921–1925 годах сформулировал балансовую теорию определения пола у дрозофилы. Он показал, что пол зависит от баланса (соотношения) числа X-хромосом и наборов аутосом. При этом Y-хромосома в определении пола у дрозофилы (в отличие от человека) не участвует.
У Менделя разные признаки гороха наследовались независимо. То, что это правило выполняется далеко не всегда, показали опыты английских генетиков У. Бэтсона (W. Bateson), Э. Саундерс (E. Saunders) и Р. Пеннета (R. Punnett) с растениями душистого горошка. Результаты опытов были опубликованы в 1905–1906 годах. Позднее Т. Морган с сотрудниками в экспериментах с дрозофилой также показали, что множество пар признаков наследуется сцепленно. Их разъединение и появление новых комбинаций происходит лишь иногда в результате кроссинговера, то есть обмена участками между гомологичными хромосомами.
Цитологические доказательства кроссинговера были получены в 1931 году: К. Штерн (C. Stern, США) использовал для этого дрозофил, а Х. Крейгтон (H. Creighton, США) и Б. МакКлинток (B. McClintock, США) — кукурузу. Они показали, что гомологичные хромосомы во время мейоза действительно способны обмениваться своими участками. Необходимо отметить, что Барбара МакКлинток вообще сыграла выдающуюся роль в генетике и за одно из своих открытий — мобильных генетических элементов * (особых последовательностей ДНК, способных перемещаться по геному) — в 1983 году была награждена Нобелевской премией.
Еще одним вариантом обмена участками является сестринский хроматидный обмен (СХО). Если при кроссинговере обмениваются хроматиды разных хромосом, то в случае СХО обмениваются хроматиды внутри одной хромосомы. Впервые СХО увидел американский генетик Д. Тейлор (J. Taylor) в 1958 году.
С кроссинговером, хоть и неоднозначно, но связано формирование в профазе мейоза особой структуры из пары гомологичных хромосом — синаптонемного комплекса. Он был открыт в 1956 году независимо двумя американскими цитологами: М. Мозесом (M. Moses) у речного рака и Д. Фоцеттом (D. Fawcett) у мыши.
Многообразие хромосом
Если понимать под хромосомами любые носители наследственной информации, то они исключительно разнообразны по размеру, форме, внешнему виду, составу и числу. Хромосомы вирусов и бактерий могут быть кольцевыми и линейными. Хромосомы хлоропластов и митохондрий имеют кольцевую форму. Ядерные хромосомы эукариот имеют линейную форму, и именно они в виде телец X- и V-образной формы обычно приходят на ум при упоминании хромосом. Их называют митотическими или метафазными, поскольку такой вид они имеют во время деления — митоза (а метафаза — это одна из его стадий).
В 1912 году российский ботаник и цитолог С.Г. Навашин показал, что метафазные хромосомы обладают индивидуальным набором признаков, включающим размер, соотношение длин плеч, наличие спутников и перетяжек. Используя положение центромеры или соотношение длин плеч, С.Г. Навашин предложил классификацию митотических хромосом, которую используют и по сей день: метацентрики, субметацентрики, акроцентрики и телоцентрики.
Число хромосом у разных видов организмов может варьировать в самых широких пределах: от двух (у пары видов растений и одного из австралийских муравьев) до 1440 у папоротника Ophioglossum reticulatum и даже 1600 у морской радиолярии Aulacantha scolymantha. У человека число хромосом составляет 46, и оно было определено только в 1955 году, а опубликовано в 1956 цитогенетиком китайского происхождения Д. Чио (J. Tjio) в соавторстве со своим руководителем А. Леваном (A. Levan) в Швеции. Несколькими месяцами позже число подтвердили британцы Ч. Форд (C. Ford) и Д. Хамертон (J. Hamerton). Количество хромосом человека пытались определить еще с конца XIX века. В разных случаях получались разные значения: 18, 24, 47 или 48, — и только в 1955 году убедились, что хромосом у человека 46. В честь этого события на здании Института генетики Университета шведского города Лунда (где это событие и случилось) в 2003 году была открыта мемориальная доска с изображением той самой метафазной пластинки, по которой и были посчитаны хромосомы. Любопытно, что число хромосом шимпанзе (48) было выяснено на 15 лет раньше.
Общепринято, что число хромосом у каждого вида живых организмов постоянно, и в подавляющем большинстве случаев так и есть. Однако у некоторых животных и растений существуют так называемые сверхчисленные, или добавочные, хромосомы. Все хромосомы основного набора называют A-хромосомами. Они присутствуют всегда, и потеря или добавление хотя бы одной из них ведет к серьезным последствиям. Добавочные же хромосомы называют B-хромосомами, и их главные особенности — необязательность наличия и непостоянство числа. Впервые сверхчисленные хромосомы были найдены Э. Уилсоном (E. Wilson, США) в 1906 году у клопа Metapodius terminalis.
Совершенно особое место среди всех типов хромосом занимают политенные хромосомы, которые имеют вид длинных толстых шнуров с поперечными полосками. Их открыл французский эмбриолог Э. Бальбиани (E. Balbiani) в 1881 году в ядрах клеток слюнных желез личинок комара Chironomus plumosus. Политенные хромосомы сыграли выдающуюся роль в развитии генетики, цитогенетики и молекулярной биологии. С их помощью была показана линейность расположения генов и однозначно доказана генетическая роль хромосом. На политенных хромосомах дрозофил был впервые описан хромосомный полиморфизм диких популяций. Именно на политенных хромосомах были открыты гены белков теплового шока — компонентов системы, охраняющей клетки всех организмов от стрессорных воздействий. Политенные хромосомы сыграли ключевую роль в исследовании системы дозовой компенсации у дрозофилы.
Эволюция хромосом и геномов
В современных цитогенетических исследованиях важную роль играет дифференциальная окраска. Впервые способность хромосом окрашиваться дифференциально (то есть неодинаково по длине) продемонстрировали англичане С. Дарлингтон (C. Darlington) и Л. Ла Кур (L. La Cour) в 1938 году. Другой важный метод исследования — это гибридизация in situ, которая позволяет определить положение любого фрагмента ДНК на хромосоме. В основе метода лежит способность нуклеиновых кислот образовывать двуцепочечные молекулы, как ДНК—ДНК, так и РНК—ДНК. Придумали этот метод в 1969 году Д. Голл (J. Gall) и М. Пардью (M. Pardue) из США и Х. Джон (H. John), М. Бирнстил (M. Birnstiel) и К. Джонс (K. Jones) из Великобритании.
Комбинация этих методов дает возможность подробно исследовать эволюцию хромосом и геномов*, а неизменным спутником эволюционного процесса являются хромосомные перестройки. По мере эволюции вида в его хромосомах неизбежно возникают перестройки, которые меняют порядок генов по сравнению с предковым видом. Чем дальше виды уходят друг от друга, тем больше хромосомных перестроек их отличает, и тем больше меняется порядок генов. Известны разные типы перестроек: делеции (потеря), дупликации (удвоение) и транслокации (перемещение) участков хромосом, которые обнаружил К. Бриджес в 1916, 1919 и 1923 годах соответственно. Еще один тип — это инверсии (поворот участка хромосомы на 180°), описанные А. Стёртевантом в 1921 году. Кроме того, существует особый тип перестроек, называемый Робертсоновской транслокацией (или центрическим слиянием). Первым ее описал американец У. Робертсон (W. Robertson) в 1916 году, сравнивая хромосомные наборы близких видов саранчи. Суть этой перестройки сводится к слиянию двух акроцентрических хромосом в одну метацентрическую или субметацентрическую. Существует и обратный процесс — центрическое разделение. В этом случае мета- или субметацентрическая хромосома делится на две акроцентрических.
Положение хромосом в ядре
Состав хромосом. ДНК
Хромосомы представляют собой структуры, состоящие из сложного комплекса ДНК, РНК и белков. Такой комплекс называется хроматином.
ДНК как химическое вещество открыл и выделил в чистом виде молодой швейцарский исследователь Ф. Мишер (F. Miescher), работая в 1868–1869 годах в университете немецкого города Тюбингена. Он изучал химический состав лейкоцитов, источником которых служил гной с бинтов из местной хирургической клиники. Ф. Мишер разработал метод разделения ядер и цитоплазмы клеток и анализировал состав ядер. Помимо белков и липидов он обнаружил вещество, которое назвал нуклеином (от слова nucleus — ядро), а сейчас оно известно как ДНК. То, что именно ДНК является носителем наследственной информации, первыми установили в 1944 году американцы О. Эйвери (O. Avery), К. МакЛауд (C. MacLeod) и М. МакКарти (M. McCarty) в экспериментах по заражению мышей пневмококками.
Состав хромосом. Белки́
C-концевые части молекул гистонов плотно свернуты, а N-концевые не имеют определенной структуры и свободно расходятся в стороны. В 1963–1964 годах было обнаружено, что некоторые аминокислотные остатки в гистонах могут быть ковалентно модифицированы, то есть ацетилированы или метилированы. Сейчас список модификаций значительно расширился, к остаткам аминокислот могут быть присоединены как относительно простые группы — метильная, ацетильная, фосфатная, — так и сложные крупные молекулы: биотин, олигопептиды или цепочки ADP-рибозы. Модификации появляются в основном на N- и, в гораздо меньшей степени, на С-концевой частях молекул гистонов.
В 2007 году был начат проект modENCODE. В его реализации участвует множество лабораторий по всему миру, одной из задач которых является построение профилей распределения самых разных белков и модификаций гистонов на хромосомах дрозофилы и нематоды Caenorhabditis elegans.
Эпигенетика
По мере развития многоклеточного организма из зиготы происходит постепенная дифференцировка клеток из тотипотентных в плюрипотентные, затем в мультипотентные, унипотентные и наконец в полностью дифференцированные клетки, из которых ничего другого уже не получится. На каждом из этих этапов работают свои наборы генов, и после прохождения стадии эти гены инактивируются. С последовательностью ДНК генов ничего не происходит, меняется белковый состав хромосомы в данном участке и, как следствие, эпигенетическое состояние генов.
Пересадка ядер соматических клеток в ооцит — это не единственный способ перезапуска эпигенетической программы. В последнее десятилетие были достигнуты колоссальные успехи в перепрограммировании соматических ядер путем искусственной активации в них генов, характерных для эмбриональных стволовых клеток. В 2006 году японские исследователи К. Такахаши (K. Takahashi) и Ш. Яманака (S. Yamanaka) получили из фибробластов мыши клетки, хоть и не идентичные эмбриональным стволовым, но очень похожие на них по морфологии, набору экспрессирующихся генов, способности к делению и дифференцировке. Они назвали такие клетки индуцированными плюрипотентными стволовыми клетками (ИПСК; iPS cells)*. Перепрограммирование произошло в результате искусственного запуска всего четырех генов (Oct3/4, Sox2, c-Myc и Klf4), которые работают в нормальных стволовых клетках. В 2007 году та же японская группа под руководством Ш. Яманака и — независимо от нее — коллектив Д. Томсона (J. Thomson) в США получили iPS клетки уже из фибробластов человека. Японцы перепрограммировали их с помощью тех же генов, что и в случае мыши, а американцы активировали гены Oct4, Sox2, Nanog и Lin28. За открытие перепрограммирования клеток Д. Гёрдон и Ш. Яманака получили в 2012 году Нобелевскую премию.
Гетерохроматин
Важным свойством гетерохроматина является способность инактивировать помещенные в него эухроматиновые гены. Это явление называется эффектом положения мозаичного типа. Оно было обнаружено в 1930 году Г. Мёллером у дрозофилы. В результате хромосомной перестройки ген white попал в гетерохроматин. Этот ген отвечает за красный цвет глаз, а если он не работает, то глаза становятся белыми. У Г. Мёллера же получились мухи, глаза которых были ни красными, ни белыми, а пятнистыми, и у разных мух пятна были разной формы и размера. Это объясняется тем, что сам ген остается неповрежденным, а лишь случайным образом инактивируется в одних клетках глаза и работает в других.
Теломеры
Дозовая компенсация
У дрозофилы природа изобрела другой механизм, противоположный по сути механизму млекопитающих: единственная X-хромосома самцов гиперактивируется и работает как две X-хромосомы самок. То, что суммарная активность двух копий какого-либо гена из X-хромосомы у самок и одной копии у самцов дрозофилы одинакова, было обнаружено еще на заре развития генетики. Это сделали К. Штерн в 1929 году и Г. Мёллер в 1931 году, так что дрозофила — это первый организм, у которого нашли дозовую компенсацию.
Ну и наконец.
Пара слов об открытии, которое не связано напрямую с хромосомами, но его очень активно используют, в том числе и для исследования разных сторон жизни хромосом. В 2008 году О. Шимомура (O. Shimomura), М. Чалфи (M. Chalfie) и Р. Циен (R. Tsien) из США получили Нобелевскую премию за открытие, выделение и применение зеленого флуоресцирующего белка (GFP) медузы Aequorea victoria. С помощью молекулярных манипуляций можно соединить ген белка GFP с геном любого другого белка и получить химерный белок, который будет выполнять как свою исходную функцию, так и светиться зеленым цветом. Это дает возможность видеть, в каких клетках работает белок, в ядре или цитоплазме, в каких частях хромосом. Кроме зеленого (GFP) сейчас известны красный (RFP) и желтый (YFP) флуоресцирующие белки*.
Читайте также:
- Казасср и туркестанская республика кратко
- Рассказ о царице из сказки о мертвой царевне и семи богатырях 5 класс кратко
- Расскажите об особенностях географического положения древней греции кратко
- Последствия отечественной войны 1812 года для россии и европы кратко
- Опишите кратко ваше физическое состояние