
Инверсия это кратко в биологии
Обновлено: 07.07.2024
Инверсией называют изменение порядка расположения генов в хромосоме, вызванное перевертыванием на 180° большого или маленького участка внутри хромосомы.
Если нормальную последовательность участков с содержащимися в них генами представить в алфавитном порядке АВСД, то при инверсии этот порядок может измениться и стать АСВД.
Для образования инверсии внутри хромосомы необходим разрыв в двух точках, и только при этом условии участок может прикрепиться противоположными концами, перевернувшись на 180°.
Перевертывание концевого участка хромосомы обычно не встречается, вероятно, потому, что конец хромосомы — теломера не обладает свойством воссоединяться с разорванным концом хромосомы. Напротив, разорванным (открытым) концам хромосом свойственна высокая способность к воссоединению. Инверсии часто связаны с рецессивным летальным эффектом, поэтому они не сохраняются в гомозиготном состоянии, и их обычно обнаруживают в гетерозиготе. Однако встречаются и такие инверсии, которые не связаны с летальным эффектом.
Существуют два типа инверсий — парацентрические инверсии и перицентрические инверсии. В случае парацентрической инверсии в хромосоме AB Ο CDEF (где Ο — означает центромеру) расположение генов может быть, например, AB Ο CEDF. В случае перицентрической инверсии последовательность генов ABCD Ο EF изменится на АВЕ Ο DCF.
Типы генных инверсий
При гомозиготной инверсии кроссинговер осуществляется нормально. При гетерозиготной инверсии кроссинговер подавляется полностью или частично. Так, например, в гетерозиготной парацентрической инверсии Ο abcdef/Ο AEDCBF хотя и происходит конъюгация хромосом, однако кроссоверных гамет при одинарном кроссинговере не обнаруживается, и это создает впечатление подавления кроссинговера.
Синапсис и перекрест в случае гетерозиотной инверсии в телоцентрической хромосоме и продукты мейоза
В силу нарушения нормального образования продуктов мейоза при скрещивании гетерозиготной по инверсии самки дрозофилы с самцом, несущим только рецессивные гены в гомозиготном состоянии в потомстве не будет возникать кроссоверных особей по данным генам.
При длинной инверсии, затрагивающей большой район хромосомы (например, в линии СIВ), иногда может обнаруживаться кроссинговер внутри инвертированного участка. В подобном случае кроссоверы возникают за счет двойного перекреста.
Следует заметить, что в гетерозиготных инверсиях по обе стороны от разрывов кроссинговер подавляется в силу отсутствия в этих районах конъюгации хроматид.
В литературе инверсию как подавителя кроссинговера принято обозначать буквой С (начальная буква английского слова crossingover). Это обозначение сохранилось с тех пор, когда подавление кроссинговера приписывалось особому гену в хромосоме.
В отличие от парацентрических инверсий в перицентрических инверсиях ни одиночный, ни двойной кроссинговер не дает ацентрических и дицентрических хроматид, а следовательно, мостов и фрагментов в мейозе. В результате кроссинговера внутри перицентрических инверсий образуются хроматиды с дупликациями и нехватками. Пыльца и яйцеклетки высших растений, имеющие указанные хроматиды, оказываются нежизнеспособными. Церицентрические инверсии могут изменять место центромеры в группе сцепления.
Инверсию можно обнаружить также цитологически в гигантских хромосомах или на пахитенной стадии мейоза.
Возможные типы инверсий
Для того чтобы осуществилась конъюгация хромосомы, имеющей инвертированный участок, с нормальной хромосомой у особи, гетерозиготной по инверсии, эти гомологичные хромосомы должны образовать петлю. При этом гены точь-в-точь пригоняются друг к другу. Следовательно, между гомологичными локусами хромосом существует взаимное притяжение огромной силы. Это очень важное явление для понимания физических и химических причин конъюгации гомологичных хромосом в профазе мейоза и ь политенных хромосомах.
Итак, в генетическом отношении инверсии характеризуются следующими свойствами:
- в гомозиготных инверсиях, если они не связаны с летальным эффектом, кроссинговер протекает нормально. Причина этого очевидна, поскольку такие гомологичные хромосомы в профазе мейоза могут свободно конъюгировать и обмениваться идентичными участками;
- в гетерозиготных инверсиях, возникающих в двуплечих хромосомах, кроссинговер подавляется только в том плече хромосомы, в котором имеется инверсия; в другом плече подавления перекреста не обнаруживается.
Однако следует иметь в виду, что не всякое подавление кроссинговера может быть отнесено за счет действия инверсии. Существуют такие генные мутации, которые могут препятствовать нормальному синапсису хромосом в профазе мейоза (асинаптические гены), как это установлено у кукурузы, ржи, хлопчатника, дурмана, ячменя, а также у дрозофилы.
Инверсии встречаются в природных популяциях животных и растений, а также могут быть получены в эксперименте, особенно под влиянием ионизирующих излучений и ряда химических веществ. Генетики полагают, что инверсии имеют существенное значение для дивергенции видов. Так, рядом исследований установлено, что отдельные расы внутри вида, например у Drosophila psetrdoobscura, могут различаться по инверсиям. Близкие виды дрозофилы в сходных хромосомах могут иметь обратный порядок расположения генов. Н. Н. Соколов провел цитогенетическое сравнение гигантских хромосом у двух видов дрозофилы (D. virilis и D. littoralis) и у их гибридов, в результате чего выяснилось, что эти два вида различаются между собой шестью инверсиями и несколькими участками, не способными к нормальной соматической конъюгации.
Экспериментально полученные инверсии часто используются в методических целях для создания линий с подавленным кроссинговером.
Сбалансированные перестройки хромосом: инверсия
Хромосомные перестройки обычно не имеют фенотипического эффекта, если они сбалансированы, поскольку весь хромосомный материал присутствует, даже если он иначе упакован. Важно различать разницу между истинно сбалансированными перестройками и теми, которые цитогенетически представляются сбалансированными, но на молекулярном уровне не сбалансированы.
Более того, из-за высокой частоты в геноме CNP, увеличивающей и без того существующие различия в много миллионов пар оснований между геномами неродственных индивидуумов, понятие сбалансированности или несбалансированности отчасти произвольно и подлежит дальнейшему исследованию и уточнению.
Даже когда структурные перестройки истинно сбалансированы, они могут представлять угрозу последующим поколениям, поскольку носители могут иметь высокую частоту несбалансированных гамет и, следовательно, имеют повышенный риск аномального потомства с несбалансированными кариотипами; в зависимости от специфической перестройки, риск может колебаться от 1 до 20%.
Существует также возможность, что один из хромосомных разрывов повредит ген, вызвав мутацию. Это хорошо описаная причина Х-сцепленных болезней у женщин-носителей сбалансированных транслокаций Х-хромосомы и аутосом, такие транслокации могут быть путеводной нитью к уточнению позиции гена, ответственного за развитие болезни.
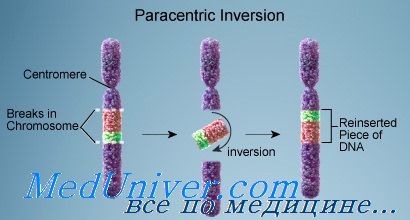
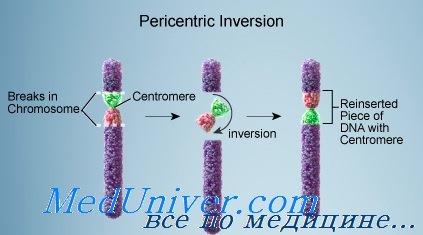
Инверсия хромосом
Инверсия появляется, когда в хромосоме происходят два разрыва, а сегмент между ними переворачивается. Инверсии бывают двух типов: парацентрические (не включающие центромеру), при которых оба разрыва происходят в одном плече; и перицентрические (включающие центромеру), при которых разрывы находятся в разных плечах. Поскольку парацентрические инверсии не изменяют соотношение плеч хромосомы, их можно выявить (если это вообще удастся) только при дифференциальной окраске или FISH-методом с локусспецифическими зондами.
Перицентрические инверсии цитогенетически выявить легче, поскольку они могут изменять соотношение плеч хромосом, а также расположение полос.
Инверсия обычно не вызывает аномалий фенотипа у носителей, поскольку относится к сбалансированным перестройкам. Ее медицинское значение — влияние на потомство; носитель любой инверсии попадает в группу риска по наличию аномальных гамет, приводящих к несбалансированному потомству, так как при инверсии при конъюгации хромосом в мейозе I формируется петля. Рекомбинация отчасти подавляется в пределах инверсионных петель, но если она происходит, то может вести к формированию несбалансированных гамет.
Формируются как гаметы со сбалансированным хромосомным набором (нормальным или с инверсией), так и гаметы с несбалансированными хромосомами в зависимости от позиции точек рекомбинации. Если инверсия парацентрическая, несбалансированные рекомбинантные хромосомы обычно ацентрические или дицентрические и не могут привести к жизнеспособному потомству, хотя есть и редкие исключения. Таким образом, на самом деле риск того, что носитель парацентрической инверсии будет иметь живорожденного ребенка с аномальным кариотипом, очень низкий.
Перицентрическая инверсия, со своей стороны, может привести к появлению несбалансированных гамет как с дупликацией, так и с делецией хромосомных сегментов. Удвоенный и потерянный сегменты — расположенные дистальнее инверсии. В целом риск для носителя перицентрической инверсии родить ребенка с несбалансированным кариотипом оценивают в 5-10%. Каждая перицентрическая инверсия, тем не менее, связана с конкретным риском.
Большие перицентрические инверсии с большей вероятностью, чем малые, приводят к жизнеспособному рекомбинантному потомству, так как в случае больших инверсий несбалансированные сегменты в рекомбинантном потомстве имеют меньший размер. Это положение иллюстрируют три хорошо изученных инверсии.
Перицентрическая инверсия хромосомы 3, происходящая от семейной пары из Ньюфаундленда, вступивших в брак в начале XIX в. — одна из немногих, для которой получено достаточно данных, чтобы оценить расхождение инвертированной хромосомы в потомстве носителей. Кариотип с inv(3) (p25q21) с тех пор был описан в ряде центров Северной Америки, в семьях, предки которых прослеживались до приморских провинций Канады.
Другая перицентрическая инверсия, связанная с выраженным дупликационным или делеционным синдромом в рекомбинантных потомках, находится в хромосоме 8, inv(8) (p23.1q22.1) и первоначально обнаружена среди испанцев юго-запада США. Эмпирические исследования показали, что носители inv(8) имеют 6% риска родить ребенка с синдромом рекомбинантной хромосомы 8, летальным нарушением с серьезными сердечными аномалиями и умственным недоразвитием. В рекомбинантной хромосоме дублирован дистальный участок 8q22.1 и утерян дистальный участок 8р23.1.
Наиболее частая инверсия у человека — небольшая перицентрическая инверсия хромосомы 9, которую отмечают почти у 1% обследованных лабораториями цитогенетики. Кариотип inv(9)(pllql2) не имеет известных опасных эффектов у носителей и не дает значимого риска выкидыша или несбалансированного потомства; поэтому обычно считается вариантом нормы.
Дополнительно к цитогенетически видимым инверсиям геномными методами обнаруживают все большее количество малых инверсий. Полагают, что многие из них клинически благоприятны, без отрицательного влияния на потомство.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
К хромосомным перестройкамотносятся: инверсии, транслокации, делеции, дупликации. Все эти мутации выявляются либо генетически (т. е. при скрещивании различных линий и анализа потомства), либо цитогенетически – путем наблюдения хромосом во время их конъюгации в профазе I мейоза или путем визуального анализа гигантских (политенных) хромосом клеток слюнных желез дрозофилы.
Гомологичные хромосомы в клетках слюнных желез дрозофилы в норме конъюгируют довольно тесно (соматическая конъюгация). При этом идентичные диски тесно прилегают друг к другу. Но в случае гетерозиготного состояния(одна хромосома – нормальная, другая – дефектная) конъюгация в этом районе нарушается. Такой участок нормальной хромосомы, не имея себе партнера в другой хромосоме, образует петлю, в то время как все остальные, гомологичные участки в обеих хромосомах тесно прилегают друг к другу.
Инверсии
Инверсия – это поворот отдельного участка хромосомы на 180°. Такие перестройки открыл А.Стёртевантв1926 г.
Инверсии бывают пара-иперицентрическими. В случае парацентрической инверсии происходят два разрыва хромосом, оба по одну сторону от центромеры. Участок между точками разрывов поворачивается на 180°:
При перицентрической инверсии точки разрывов расположены по обе стороны от центромеры:
Для инверсий приняты специальные обозначения: например, In(1)BEозначает, что инверсия (In) произошла в первой хромосоме (1), BE – инвертированный район.
У гомозигот по инверсиям кроссинговер происходит нормально. У особей, гетерозиготных по любому типу инверсии, в хромосомах при конъюгации образуется петля. Вот так, например, это происходит при парацентрической инверсии:
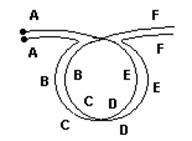
Обе комбинации летальны. Таким образом, в результате кроссинговера образуются нежизнеспособные гаметы, и потомства нет.
Двойной кроссинговер, если он случается в пределах инверсии, восстанавливает образование гамет.
При перицентрической инверсии, в случае перекреста между генами С и D, также получаются два продукта:
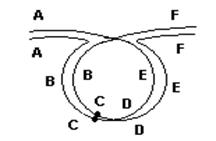
Лекция №10
Тема лекции: Хромосомные перестройки и геномные мутации
План лекции: 1. Инверсии.
5. Механизмы возникновения хромосомных перестроек и их значение.
6. Типы геномных мутаций и их причины. Полиплоидия
К хромосомным перестройкамотносятся: инверсии, транслокации, делеции, дупликации. Все эти мутации выявляются либо генетически (т. е. при скрещивании различных линий и анализа потомства), либо цитогенетически – путем наблюдения хромосом во время их конъюгации в профазе I мейоза или путем визуального анализа гигантских (политенных) хромосом клеток слюнных желез дрозофилы.
Гомологичные хромосомы в клетках слюнных желез дрозофилы в норме конъюгируют довольно тесно (соматическая конъюгация). При этом идентичные диски тесно прилегают друг к другу. Но в случае гетерозиготного состояния(одна хромосома – нормальная, другая – дефектная) конъюгация в этом районе нарушается. Такой участок нормальной хромосомы, не имея себе партнера в другой хромосоме, образует петлю, в то время как все остальные, гомологичные участки в обеих хромосомах тесно прилегают друг к другу.

Инверсии
Инверсия – это поворот отдельного участка хромосомы на 180°. Такие перестройки открыл А.Стёртевантв1926 г.
Инверсии бывают пара-иперицентрическими. В случае парацентрической инверсии происходят два разрыва хромосом, оба по одну сторону от центромеры. Участок между точками разрывов поворачивается на 180°:
При перицентрической инверсии точки разрывов расположены по обе стороны от центромеры:
Для инверсий приняты специальные обозначения: например, In(1)BEозначает, что инверсия (In) произошла в первой хромосоме (1), BE – инвертированный район.
У гомозигот по инверсиям кроссинговер происходит нормально. У особей, гетерозиготных по любому типу инверсии, в хромосомах при конъюгации образуется петля. Вот так, например, это происходит при парацентрической инверсии:
Обе комбинации летальны. Таким образом, в результате кроссинговера образуются нежизнеспособные гаметы, и потомства нет.
Двойной кроссинговер, если он случается в пределах инверсии, восстанавливает образование гамет.
При перицентрической инверсии, в случае перекреста между генами С и D, также получаются два продукта:


Инверсии — это хромосомные перестройки, при которых участок хромосомы поворачивается на 180°. Эволюционное значение инверсий связано в первую очередь с тем, что инверсии затрудняют рекомбинацию — обмен участками гомологичных хромосом при мейозе. Анализ данных по 411 видам воробьинообразных птиц показал, что у близкородственных видов с перекрывающимися ареалами число крупных инверсий повышено по сравнению с видами, ареалы которых не пересекаются. По-видимому, отбор способствует закреплению инверсий у симпатрических видов, способных к гибридизации. Результаты согласуются с гипотезой о том, что инверсии сглаживают вредные последствия межвидовой гибридизации, способствуя сохранению коадаптированных (приспособленных друг к другу) генных комплексов. Благодаря инверсиям гибридное потомство реже получает конфликтующие комбинации генов. С другой стороны, инверсии препятствуют обратному слиянию недавно разделившихся видов, помогая им сохранять свою идентичность даже в условиях частой гибридизации.
1. Полезны или вредны хромосомные инверсии?
Вновь возникающие инверсии, по идее, должны снижать приспособленность — а значит, отсеиваться отбором. Есть совсем очевидные случаи: например, если граница повернувшегося на 180° участка хромосомы разрежет пополам какой-то важный ген, то ген выйдет из строя. Такая инверсия, конечно, будет быстро отбракована и не сможет закрепиться в популяции. Но даже если границы инвертированного участка попадут в нефункциональные районы хромосомы, инверсия всё равно создаст определенные проблемы у особей, гетерозиготных по инверсии (то есть таких, у которых в двух гомологичных хромосомах данный участок повернут в разные стороны). Если при мейозе у такой особи рекомбинация (см. Chromosomal crossover) произойдет внутри инвертированного участка, то могут получиться гаметы с неправильным набором генов. Например, если инвертированный участок содержит гены А и B, то одни гаметы могут получить две копии гена А и ни одной копии B, другие — наоборот. Это, скорее всего, плохо скажется на приспособленности потомства.
Впрочем, сам механизм кроссинговера устроен таким образом, что вероятность рекомбинации участков, повернутых в разные стороны, резко снижается. Иными словами, инверсия блокирует рекомбинацию инвертированного участка. Благодаря этому многие инверсии оказываются безвредными (или, по крайней мере, не настолько вредными, чтобы исключить возможность их распространения за счет дрейфа в маленьких популяциях). Аллели, расположенные на инвертированном участке, при этом начинают наследоваться все вместе, единым блоком.
2. Две гипотезы о пользе инверсий
Геномные исследования показали, что в эволюции птиц крупные инверсии фиксируются (то есть не только возникают у какой-то одной особи, но достигают 100-процентной частоты в генофонде вида) довольно часто — в отличие от других крупномасштабных хромосомных перестроек, таких как транслокации (переносы участков с одной хромосомы на другую), слияние или разделение хромосом. Это наводит на мысль, что инверсии, возможно, оказываются полезными (или, что то же самое, поддерживаются отбором) чаще, чем принято считать.
Вопрос о том, в чем состоит эта предполагаемая польза, далек от разрешения. С одной стороны, она может быть никак не связана с блокировкой рекомбинации. Например, инверсия может привести к тому, что какие-то гены, оказавшись в ином окружении, будут регулироваться более выгодным для организма образом.
3. Проверяемые следствия
Если эта польза не связана с блокировкой рекомбинации, а связана, например, с выгодным изменением регуляции генов, то полезные инверсии, по идее, должны вести себя в ходе эволюции так же, как и все прочие редкие полезные мутации. А именно, скорость их накопления должна положительно коррелировать с численностью популяции (потому что чем больше выборка, тем чаще в ней, при прочих равных, будут происходить маловероятные события, такие как возникновение полезной мутации). Перекрывание ареалов близкородственных видов в этом случае не должно ускорять фиксацию инверсий. Более того, если польза инверсий не зависит от генетического контекста, то инверсии должны легко пересекать межвидовые границы в зонах гибридизации, то есть легко проникать из одного видового генофонда в другой и свободно распространяться там. В этом случае можно ожидать, что у симпатрических близкородственных видов будет не больше, а, скорее, даже меньше различий по инверсиям, чем у аллопатрических (проживающих на разных территориях).
Если же польза инверсий — в защите коадаптированных генных комплексов от распада и перемешивания, то следует ожидать, что инверсии будут быстрее всего накапливаться не в самых больших популяциях, а в тех, которые реально подвергаются угрозе такого распада и перемешивания. В первую очередь — в симпатрических популяциях близкородственных видов.
4. Инверсии у воробьинообразных
Эволюционные биологи из Чикагского университета Дэниел Хупер (Daniel M. Hooper) и Тревор Прайс (Trevor D. Price) проверили эти предсказания на основе данных о крупных хромосомных инверсиях у воробьинообразных (отряд Passeriformes). Ранее они пытались провести такую проверку на 32 видах вьюрковых ткачиков, представляющих лишь одно из семейств воробьинообразных. Оказалось, что в этом семействе частота фиксации инверсий положительно коррелирует как с численностью вида (которую приблизительно оценивали по площади ареала), так и с перекрыванием видовых ареалов. Это позволило отвергнуть гипотезу о дрейфе как об основном механизме распространения инверсий. Однако вопрос о природе пользы, приносимой фиксирующимися инверсиями, остался открытым. Дело в том, что у вьюрковых ткачиков площади ареалов положительно коррелируют со степенью их перекрывания, а малый размер выборки не позволил понять, какой из факторов в конечном счете важнее (D. M. Hooper, T. D. Price, 2015. Rates of karyotypic evolution in Estrildid finches differ between island and continental clades).
В новой статье, опубликованной в журнале Nature ecology and evolution, Хупер и Прайс рассмотрели более представительную выборку, включающую 411 видов воробьинообразных из 59 семейств. Анализировались только крупные перицентрические (то есть захватывающие центромеру) инверсии, которые можно обнаружить, просто разглядывая хромосомы под микроскопом. Полных геномных данных, позволяющих выявить все инверсии, по большинству рассмотренных видов пока нет.
Наложив данные по инверсиям на эволюционное дерево (которое для воробьинообразных надежно реконструировано по молекулярным и палеонтологическим данным), авторы оценили скорость фиксации инверсий в разных линиях (рис. 2). Оказалось, что эта скорость сильно варьирует: в одних линиях ни одной инверсии не закрепилось за 28 млн лет, тогда как в других инверсии фиксировались в среднем каждые 500 или даже 200 тысяч лет.
Рис. 2. Эволюционное дерево 411 видов воробьинообразных. Цветовая шкала показывает скорость фиксации крупных перицентрических инверсий в разных эволюционных линиях (в штуках за миллион лет). Справа — список семейств. Цифры в скобках — количество фиксированных инверсий, обнаруженных у данного семейства. Первая цифра — аутосомы, вторая — половая хромосома Z, которая присутствует в двух экземплярах у самцов птиц и в одном — у самок (см. ZW-определение пола). У 32 видов был обнаружен внутривидовой полиморфизм по инверсиям (то есть инверсии, имеющиеся только у некоторых особей данного вида), но эти данные на рисунке не отражены. Рисунок из обсуждаемой статьи в Nature Ecology & Evolution
5. Перекрывание видовых ареалов — лучший предиктор числа инверсий
Для дальнейшего анализа была отобрана 81 клада (независимая веточка эволюционного дерева), куда попало в общей сложности 284 вида из 411. Клады отбирались по определенным критериям: в частности, каждая клада должна включать не менее трех видов, эти виды должны быть близкородственными (время расхождения не более 15 млн лет), вся дивергенция клады должна быть приурочена к одному континенту, все виды должны иметь один и тот же тип питания (зерноядные, насекомоядные, всеядные и т. д.) Для каждой клады вычислили общую длину ветвей (она отражает суммарное время существования видов), среднюю площадь видовых ареалов, среднюю величину их перекрывания, а также учитывали ряд экологических показателей.
Статистический анализ этих данных, выполненный с использованием нескольких альтернативных подходов, показал, что число инверсий в кладе тесно связано только с двумя факторами: длиной ветвей и степенью перекрывания видовых ареалов. Площади ареалов (которые использовались в качестве грубой оценки численности вида) сами по себе, по-видимому, не влияют на скорость фиксации инверсий. Они могут быть связаны с ней лишь опосредованно — за счет того, что, при прочих равных, обширный ареал будет с большей вероятностью перекрываться с каким-нибудь другим ареалом. Длина ветвей отражает время существования видов. Если же говорить о скорости фиксации инверсий (то есть о числе инверсий, фиксирующихся за миллион лет), то эта величина, судя по полученным результатам, определяется в первую очередь степенью перекрывания ареалов близкородственных видов.
Типичная ситуация показана на рис. 1, где представлена история фиксации инверсий в одной из рассмотренных клад — у зеленушек (род Chloris). Один из трех видов клады, C. chloris, имеет обширный ареал, не пересекающийся с ареалами близких видов. В соответствии с этим у данного вида число зафиксировавшихся инверсий равно нулю. Два других вида, C. ambigua и C. sinica, имеют частично перекрывающиеся ареалы. При этом известно, что в зоне симпатрии (совместного проживания) между ними происходит гибридизация. Эти виды разделились 2,2 млн лет назад: за такое время между видами птиц, как правило, полная генетическая несовместимость не успевает развиться (в исследованной выборке есть гибридизующиеся виды, разошедшиеся 5–7 млн лет назад, причем нужно учитывать, что гибридизация может стать причиной заниженных оценок времени дивергенции — то есть на самом деле эти пары видов могли разойтись существенно раньше). В соответствии с этим у C. ambigua зафиксировалось три инверсии, у C. sinica — одна, так что общее число различий по инверсиям между ними стало равно четырем. Аналогичная картина наблюдается и в других кладах: инверсии чаще всего фиксируются у симпатрических родственных видов, которые либо гибридизуются сейчас, либо могли гибридизоваться в недавнем прошлом. Что касается площади ареалов, то в данном случае она явно не оказывает положительного влияния на число инверсий. Даже наоборот: у вида с самым обширным ареалом инверсий меньше всего, а вид с наименьшим ареалом оказался чемпионом по инверсиям в данной кладе. При этом время, в течение которого инверсии могли накапливаться, одинаково для всех трех видов клады.
Авторы также использовали имеющиеся геномные данные, чтобы проверить гипотезу о том, что на скорость фиксации инверсий сильно влияет скорость их появления. В таком случае фиксированные (достигшие 100-процентной частоты в генофонде) инверсии должны чаще встречаться в тех участках генома, где инверсии чаще возникают, — например, в районах с большим количеством повторяющихся нуклеотидных последовательностей. Это, однако, не подтвердилось. Напомним, что исследовались только очень крупные инверсии, заметные под микроскопом. Вполне возможно, что с мелкими инверсиями дело обстоит иначе (как на то указывают результаты ряда исследований в области сравнительной геномики). Ну а скорость фиксации крупных инверсий, по-видимому, определяется всё же отбором, а не мутагенезом.
6. Инверсии защищают молодые виды от вредных последствий гибридизации
Таким образом, полученные Хупером и Прайсом результаты согласуются с гипотезой о том, что отбор поддерживает инверсии у близких видов, способных к гибридизации, если их ареалы перекрываются, то есть в том случае, когда существует реальная опасность смешивания и разрушения коадаптированных генных комплексов. Очевидно, из этого следует, что польза, приносимая фиксирующимися инверсиями, связана в первую очередь с блокировкой рекомбинации.
Дополнительным аргументом в пользу этого является обнаруженная авторами повышенная частота фиксации инверсий на половой хромосоме Z. Эта хромосома имеется в двух экземплярах у самцов птиц и в одном — у самок (см. ZW-определение пола). Давно известно, что конфликты между генами двух гибридизующихся видов наиболее опасны для гибридного потомства в том случае, если несовместимые гены находятся на рекомбинирующей половой хромосоме (у млекопитающих это хромосома Х, у птиц — Z). При этом страдает в первую очередь приспособленность гетерогаметного пола. У млекопитающих это самцы, у которых только одна Х-хромосома, у птиц — самки, у которых только одна Z-хромосома (см. Haldane’s rule). У гетерогаметного пола конфликтующие гены, сидящие на хромосоме X или Z, присутствуют в единственном экземпляре, и поэтому конфликт сразу проявляется в фенотипе. У гомогаметного пола есть еще вторые копии этих генов, сидящие на второй половой хромосоме. Они могут сгладить конфликт или вовсе не дать ему проявиться (в этом случае говорят о рецессивной несовместимости). Поэтому, если идеи авторов верны, следует ожидать, что отбор будет сильнее всего поддерживать инверсии именно на хромосоме Z, что и подтверждается.
Это альтернативное объяснение не может быть отвергнуто на основе имеющихся данных, хотя авторы считают его несколько менее правдоподобным, чем их основная гипотеза о положительном влиянии симпатрии на распространение инверсий. Ведь если накопление инверсий — не результат симпатрии, а лишь причина сохранения видовой идентичности в условиях симпатрии, то вопрос о механизмах, способствующих накоплению инверсий (в том числе в условиях аллопатрии), так и остается без ответа. Впрочем, авторы допускают, что обе версии могут быть верны одновременно. То есть инверсии, с одной стороны, распространяются в условиях симпатрии под действием отбора, потому что сглаживают вредные последствия гибридизации, а с другой — не дают двум генофондам полностью смешаться, помогая видам сохранять свою идентичность.
Рассмотренное исследование вносит важный вклад в понимание роли инверсий в видообразовании. Поскольку данная тема сейчас стала весьма популярной, новые открытия в этой области вряд ли заставят себя долго ждать.
Источник: Daniel M. Hooper & Trevor D. Price. Chromosomal inversion differences correlate with range overlap in passerine birds // Nature Ecology & Evolution. Published online 28 August 2017.
Читайте также: