
Химический состав вирионов кратко
Обновлено: 02.07.2024
Вирион – элементарная инфекционная единица вируса, форма в которой вирусы существуют вне живой клетки. Строение, размеры и химический состав вирионов были установлены только с появлением и развитием электронной микроскопии, физико-химических и рентгеноструктурных методов анализа.
Морфология вириона
Вирион (вирусная частица) состоит из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки.
В зависимости от разновидности нуклеиновой кислоты в вирусной частице, вирусы подразделяются на:
- ДНК-содержащие – в качестве генетического материала имеют одно- или двухцепочечную ДНК, линейной или кольцевой конфигурации (вирус простого герпеса, вирус оспы);
- РНК-содержащие – в качестве генетического материала содержат одно- или двухцепочечнуюРНК (вирус гриппа).
Вирусы с одноцепочечной РНК подразделяются на два типа:
Белковая оболочка, окружающая нуклеиновую кислоту называется капсидом. Капсид состоит из отдельных субъединиц – капсомеров.
Капсиды отдельных вирионов может окружать дополнительная мембрана – суперкапсид.
Капсид обычно имеет симметричное строение. Различают несколько типов симметрии:
- спиральную – характерна для вирусов с палочковидной и нитевидной формой строения.
- кубическую (икосаэндрическую) – характерна для изомерических вирусов;
- комбинированный (сложный) тип симметрии – характерен для для бацилловидных и пулевидных вирионов.
В частности, размеры вирионов различных фитовирусов колеблются в значительных пределах. Каждому вирусу соответствуют частицы определенного размера одного или нескольких видов. Вирионы изометрических вирусов по величине диаметра варьируют от 17 нм до 75 нм. Палочковидные, нитевидные и бациловидные вирионы могут достигать в длину от нескольких десятков до 2000 нм. Диаметр у нитевидных и палочковидных – 3–10 нм, у бацилловидных – 18–75 нм.
Химический состав вириона
В химический состав вирионов входят нуклеиновые кислоты, белки и другие компоненты.
Содержание нуклеиновых кислот у вирусов различно:
- вирионы с изометрической (сферической) формой капсида содержат 5–15% нуклеиновой кислоты. Некоторые из них способны формировать частицы совсем лишенные нуклеиновой кислоты;
- вирионы с палочковидной и нитевидной формы со спиральной симметрией – содержание нуклеиновой кислоты составляет 5%;
- бациловидные, липидосодержащие вирусы – около 1% нуклеиновой кислоты.
Нуклеиновая кислота каждого вируса имеет характерную последовательность и нуклеотидный состав, который определяет последовательность аминокислот и свойства и поведение вирус-специфических белков. Молекулярная масса нуклеиновой кислоты в инфекционных частицах (вирионах) варьирует от 0,4х10 у вируса некроза табака до 15,5х10у вируса раневых опухолей клевера. У большинства фитовирусов молекулярная масса нуклеиновой кислоты составляет в среднем 2х10.
Большая часть вириона – это белок. Молекулярная масса белковых субъединиц у различных вирусов может варьировать от 10 до 80 тыс. Размер белковых молекул фитовирусов расположен в диапазоне от 150 до 600 и более аминокислотных остатков. Вирусов со сложной структурой содержат липиды, содержание которых достигает 20%.
В состав вириона входит вода. Она может составлять около 50% веса вирусных кристаллов и 10–50% веса вирусных частиц в суспензии. Отмечается, что при описании состава вирионов содержание воды в них не учитывают и указывают содержание нуклеиновой кислоты, белка и прочих компонентов в пересчете на сухой вес частицы.
Одновременно вирусы могут содержать различное количество ионов металлов, полиамины (около 1% веса). Ряд вирусов содержат фрагменты типа РНК-транскриптазы.
(c) Справочник AgroXXI

Вирион – элементарная инфекционная единица вируса, форма в которой вирусы существуют вне живой клетки [1] .
Строение, размеры и химический состав вирионов были установлены только с появлением и развитием электронной микроскопии, физико-химических и рентгеноструктурных методов анализа [1] .

Вирус пятнистого увядания томата (Tomato spotted wilt virus)

Поражение клеток садового помидора (Solanum lycopersicum) [4]
Морфология вириона
Вирион (вирусная частица) состоит из нуклеиновой кислоты (ДНК или РНК) и белковой оболочки [3] .
В зависимости от разновидности нуклеиновой кислоты в вирусной частице, вирусы подразделяются на:
- ДНК-содержащие – в качестве генетического материала имеют одно- или двухцепочечную ДНК, линейной или кольцевой конфигурации (вирус простого герпеса, вирусоспы);
- РНК-содержащие – в качестве генетического материала содержат одно- или двухцепочечную РНК (вирус гриппа) [3] .
Вирусы с одноцепочечной РНК подразделяются на два типа:
Белковая оболочка, окружающая нуклеиновую кислоту называется капсидом. Капсид состоит из отдельных субъединиц – капсомеров [3] .
Капсиды отдельных вирионов может окружать дополнительная мембрана – суперкапсид [3] .
Капсид обычно имеет симметричное строение. Различают несколько типов симметрии:
- спиральную – характерна для вирусов с палочковидной и нитевидной формой строения.
- кубическую (икосаэндрическую) – характерна для изомерических вирусов;
- комбинированный (сложный) тип симметрии – характерен для для бацилловидных и пулевидных вирионов [2] .
В частности, размеры вирионов различных фитовирусов колеблются в значительных пределах. Каждому вирусу соответствуют частицы определенного размера одного или нескольких видов. Вирионы изометрических вирусов по величине диаметра варьируют от 17 нм до 75 нм. Палочковидные, нитевидные и бациловидные вирионы могут достигать в длину от нескольких десятков до 2000 нм. Диаметр у нитевидных и палочковидных – 3–10 нм, у бацилловидных – 18–75 нм [2] .
Химический состав вириона
В химический состав вирионов входят нуклеиновые кислоты, белки и другие компоненты [2] .
Содержание нуклеиновых кислот у вирусов различно:
- вирионы с изометрической (сферической) формой капсида содержат 5–15% нуклеиновой кислоты. Некоторые из них способны формировать частицы совсем лишенные нуклеиновой кислоты;
- вирионы с палочковидной и нитевидной формы со спиральной симметрией – содержание нуклеиновой кислоты составляет 5%;
- бациловидные, липидосодержащие вирусы – около 1% нуклеиновой кислоты [2] .
Нуклеиновая кислота каждого вируса имеет характерную последовательность и нуклеотидный состав, который определяет последовательность аминокислот и свойства и поведение вирус-специфических белков. Молекулярная масса нуклеиновой кислоты в инфекционных частицах (вирионах) варьирует от 0,4х10 6 у вируса некроза табака до 15,5х10 6 у вируса раневых опухолей клевера. У большинства фитовирусов молекулярная масса нуклеиновой кислоты составляет в среднем 2х10 6 [2] .
Большая часть вириона – это белок. Молекулярная масса белковых субъединиц у различных вирусов может варьировать от 10 до 80 тыс. Размер белковых молекул фитовирусов расположен в диапазоне от 150 до 600 и более аминокислотных остатков. Вирусы со сложной структурой содержат липиды, содержание которых достигает 20% [2] .
В состав вириона входит вода. Она может составлять около 50% веса вирусных кристаллов и 10–50% веса вирусных частиц в суспензии. Отмечается, что при описании состава вирионов содержание воды в них не учитывают и указывают содержание нуклеиновой кислоты, белка и прочих компонентов в пересчете на сухой вес частицы [2] .
Одновременно вирусы могут содержать различное количество ионов металлов, полиамины (около 1% веса). Ряд вирусов содержат фрагменты типа РНК-транскриптазы [2] .
Простые (безоболочечные) вирусы состоят из нуклеиновой кислоты и белка и представляют собой нуклеопротеиды или нуклеокапсиды. Сложные (оболочечные) вирусы кроме нуклеиновой кислоты и белка содержат также липиды и углеводы.
Нуклеиновые кислоты. Клетки всех живых организмов содержат два типа нуклеиновых кислот — ДНК и РНК. Их структура относительно однообразна: ДНК представляет собой двуспиральную молекулу, РНК — односпиральную молекулу. Функция ДНК заключается в хранении и репликации наследственной информации, таким образом, она является клеточным геномом. РНК представлена в клетке в трех формах: информационной (иРНК), рибосомальной (рРНК) и транспортной (тРНК). Каждая из них выполняет определенную функцию: иPHК образуется в результате транскрипции генома и передает информацию с генома на белоксинтезирующий аппарат клетки, рРНК является структурным компонентом рибосом, тРНК доставляет аминокислоты белоксинтезирующему аппарату клетки.
В отличие от клетки вирусы содержат один тип нуклеиновой кислоты — или ДНК, или РНК. Каждая из них выполняет функцию вирусного генома. Структура нуклеиновых кислот у разных вирусов весьма разнообразная. По количеству цепей они бывают одно — и двуспиральными, по форме — линейными и кольцевыми (циркулярными), а также непрерывными и фрагментированными.
Содержание нуклеиновой кислоты в вирионе различных вирусов составляет от 1 % у ортомиксо- и парамиксовирусов до 32 % у парвовирусов и не коррелирует ни с систематическим положением вируса, ни со степенью сложности его организации.
Вирусные ДНК. Молекулярная масса ДНК различных вирусов варьирует в широких пределах: от 2 МД у цирко — и парвовирусов до 375 МД у поксвирусов. Самые большие геномы содержат до нескольких сотен генов, самые маленькие — несколько генов. По структуре молекулы ДНК бывают одно — и двуспиральными, линейными и кольцевыми. У вирусов с двуспиральными ДНК информация обычно закодирована на обеих спиралях, что говорит о максимальной экономии генетического материала. Большинство нуклеотидных последовательностей в молекуле ДНК встречается однократно. Однако в концевом фрагменте линейных ДНК возможно наличие ее начального участка в виде повтора, который бывает прямым или инвертируемым. Благодаря таким повторам молекулы ДНК могут приобретать циркулярную форму, которая обеспечивает их устойчивость к эндонуклеазам. Кроме того, стадия образования циркулярной формы обязательна для интеграции вирусной ДНК с геномом клетки.
Вирусные РНК. Молекулярная масса вирусных РНК варьирует в пределах от 4—5 МД у нодавирусов до 32 МД у реовирусов.
По предложению Балтимора (1971) вирусы с односпиральными РНК из-за различий в функциях генома было принято разделять на две подгруппы. У вирусов первой подгруппы вирусный геном обладает функциями иРНК и их условно обозначают как плюс-нитевые вирусы, или вирусы с позитивным геномом. У вирусов второй группы РНК не обладает функцией иРНК. На ней, как на матрице, синтезируется комплементарная молекула. Это происходит только в присутствии вирусного белка — фермента транскриптаза, который обязательно находится в структуре минус-нитевых вирусов (в клетках — ее аналога нет).
Существуют вирусы, которые содержат как плюс-нитевые, так и минус-нитеные РНК-гены, так называемые амбигюлирные РНК.
Белки. Белки всех известных в настоящее время вирусов позвоночных являются основными компонентами вирионов и составляют от 57 до 90 % массы вириона. По аминокислотному составу вирусные белки принципиально не отличаются от состава белков животных.
В геноме вирусов кодируются две группы белков: структурные, которые входят в состав вирионов потомства, и неструктурные, участвующие в репродукции вируса на разных этапах, но не входящие в состав вирионов.
Структурные белки в составе вириона варьируют в широких пределах, что зависит от сложности организации вириона. Простые вирусы животных содержат 3—4 белка, сложные, например вирусы оспы, — более 30. Среди структурных белков различают две группы — капсидные и пепломеры. Первая группа включает белки, которые формируют капсид (от греч. capsa — вместилище), окружающий нуклеиновую кислоту, а также геномные белки и ферменты; вторая группа — белки суперкапсидной оболочки, называемой пеплос (от греч. peplos — покров, мантия). Простые вирусы содержат только капсидные белки, сложные — и капсидные, и пепломеры.
Белки в составе вирусного калсида называются капсомерами. Основной их функцией является защита вирусного генома от неблагоприятных воздействий внешней среды. Они представляют собой идентичные полипептидные цепи (белковые субъединицы), которые обладают способностью к самосборке. Сборка капсида из субъединиц запрограммирована в первичной структуре белка и происходит самопроизвольно или при взаимодействии с нуклеиновой кислотой. Принцип субъединичности в строении капсида — уникальное свойство капсидных белков, благодаря которому достигается огромная экономия генетического материала. Кроме того, в механизме самосборки заложена возможность контроля за белковыми субъединицами: дефектные и, чужеродные белковые цепи не включаются в капсид. Принцип самосборки характерен только для простых вирусов. Сложные вирусы сборку осуществляют по более сложному многоступенчатому механизму. Однако отдельные ее этапы (формирование капсидов и нуклеокапсидов) основаны на самосборке.
Суперкапсидные белки (пепломеры) находятся в липопротеидной оболочке сложных вирусов. Они либо пронизывают липидный бислой вириона, либо не доходят до его внутренней поверхности. Являясь типичными внутримембранными белками, они, как правило, гликозилированы (гликопротеиды), т. е. к молекуле белка в определенных местах прикреплены углеводные цепи. Гликозилирование осуществляют клеточные ферменты, поэтому один и тот же вирус, но реплицирующийся в разных клетках, может иметь разные углеводные остатки по составу углеводов, длине углеводной цепи, месту прикрепления к белку.
Неструктурные белки менее изучены. Существуют определенные трудности в их выделении из зараженных клеток и очистки от клеточных белков. К ним относятся: предшественники вирусных белков, которые существуют в зараженной клетке очень непродолжительное время, а затем нарезаются; ферменты синтеза РНК. и ДНК — полимеразы; регуляторы стадий репродукции вирусов; ферменты, модифицирующие вирусные белки — протеиназы и протеинкиназы.
Липиды и углеводы. В состав вирионов всех сложных (оболочечных) вирусов позвоночных кроме нуклеиновой кислоты и белков входят липиды и углеводы.
Состав липидов вирионов сходен с липидным составом клетки хозяина: примерно 50—60 % составляют фосфолипиды и 20—30 % — холестерин. У отдельных представителей липидов содержится до 20—35 % от массы вириона (ортомиксо-, ретро-, буньявирусы). Липиды обнаружены только в суперкапсидной оболочке вирионов и имеют клеточное происхождение. Это связано с тем, что оболочечные вирусы формируются путем почкования на плазматической мембране клеток. Поэтому суперкапсидная оболочка вирионов представляет собой мембрану клетки-хозяина, модифицированную за счет встроенных в нее вирусных белков — пепломеров. Липидный компонент стабилизирует структуру вирусных частиц, поэтому их обработка детергентами или липазами приводит к потере инфекционности.
Исключение составляют вирусы оспы. У них липиды не образуют дифференцированной оболочки. Обработка вирусов осповакцины жирорастворителями не приводит к потере инфекционной активности или каким-либо другим структурным изменениям вириона.
Углеводы находятся в вирионах в виде гликопротеинов, встроенных в суперкапсидный слой, а также гликолипидов и имеют клеточное происхождение. У отдельных представителей вирусов позвоночных содержание углеводов доходит до 7—8 % от массы вириона (ортомиксо-, тогавирусы). Химический состав их полностью определяется клеточными ферментами, которые обеспечивают перенос и присоединение сахарных остатков. В вирионах в основном обнаруживают фруктозу, сахарозу, маннозу, галактозу, нейраминовую кислоту, глюкозамин. Углеводы являются каркасом для локальных участков гликопротеидов, обеспечивают сохранение конформации белковых молекул и защищают от действия протеаз.
Компоненты клетки-хозяина. В составе вирионов могут обнаруживаться некоторые компоненты клетки-хозяина. Это могут быть белки или цельные клеточные структуры. Например, в составе некоторых оболочечных вирусов находится белок цитоскелета актин, в составе папиллома — и полиомавирусов содержатся клеточные гистоны, у ареновирусов обнаружены рибосомы. Клеточные компоненты могут включаться в вирион случайно или закономерно. Во втором случае они могут играть существенную роль в репродукции вирусов, как, например, гистоны для папиллома — и полиомавирусов.
Структурная организация вирионов. Вирионы (вирусные частицы) по архитектуре подразделяются на два типа: имеющие внешнюю липопротеидную оболочку (сложные, или оболочечные, вирусы) и не имеющие такой оболочки (простые, или безоболочечные, вирусы).
Капсомеры соединяются друг с другом строго определенным образом и образуют только два типа капсидов: спиральный и изометрический (квазиметрически й). Только такие способы укладки капсомеров вокруг нуклеиновой кислоты (типы симметрии) обеспечивают формирование энергетически экономичных структур с минимумом свободной энергии.
Изометрические капсиды представляют структуры, в которых капсомеры соединяются между собой в правильные многогранники, в центре которых расположен геном. Такой способ укладки капсомеров называется кубическим типом симметрии, а многогранник — икосаэдром. Это означает, что он симметричен в трех взаимно перпендикулярных направлениях и его линейные размеры вдоль прямоугольных осей идентичны. Обычно изометрические капсиды состоят из 60 (или кратных 60) геометрически идентичных элементов, которые имеют 12 вершин, 20 граней и 20 ребер. Так, капсид вируса полиомиелита образован 60 белковыми структурными единицами, каждая из которых состоит из четырех полипептидных цепей (химических единиц) — vp1, vp2, vp3, vp4 (virion protein).
Наиболее сложно структурно организованы вирусы оспы. Их сердцевина (нуклеоид), содержащая вирусную ДНК в составе нуклеотида, имеет форму двояковогнутого кольца и окружена двумя линзообразными латеральными тельцами. Капсид окружен дополнительными внутренними белковыми структурами — вирусным матриксом. По архитектуре вирусы оспы отличаются от всех других вирусов позвоночных и их принято называть вирусами со сложным типом симметрии.
Вирусы. Вирион. Морфология вирусов. Размеры вирусов. Нуклеиновые кислоты вирусов.
Вирусы [от лат. virus, яд] — наименьшие по размерам агенты, имеющие геном, окружённый белковой оболочкой. Вирусы не воспроизводятся самостоятельно, они — облигатные внутриклеточные паразиты, репродуцирующиеся только в живых клетках. Все вирусы существуют в двух формах. В настоящее время известны вирусы бактерий (бактериофаги), грибов, растений и животных.
Внеклеточная форма — вирион — включает в себя все составные элементы (капсид, нуклеиновую кислоту, структурные белки, ферменты и др.). Внутриклеточная форма — вирус — может быть представлена лишь одной молекулой нуклеиновой кислоты, так как, попадая в клетку, вирион распадается на составные элементы.
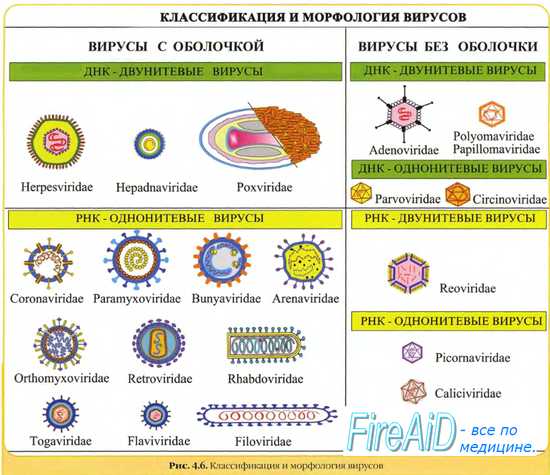
Морфология вирусов. Размеры вирусов.
Несмотря на внутриклеточный паразитизм, среди вирусов имеются крупные виды, соизмеримые по размерам с микоплазмами и хламидиями. Например, вирус натуральной оспы достигает 400 нм и вполне сравним с риккетсиями (300-500 нм) и хламидиями (300-400 нм). По морфологии выделяют вирусы палочковидные (например, возбудитель лихорадки Эбола), пуле-видные (вирус бешенства), сферические (герпесвирусы), овальные (вирус оспы), а также бактериофаги, имеющие сложную форму (рис. 2-1). При всём разнообразии конфигураций, размеров и функциональных характеристик вирусам присущи некоторые общие признаки. В общем виде зрелая вирусная частица (вирион) состоит из нуклеиновой кислоты, белков и липидов, либо в его состав входят только нуклеиновые кислоты и белки.
Нуклеиновые кислоты вирусов
Вирусы содержат только один тип нуклеиновой кислоты, ДИК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпстайна-Барр — ДНК-содержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный. Наиболее простой вирусный геном кодирует 3-4 белка, наиболее сложный — более 50 полипептидов. Нуклеиновые кислоты представлены однонитевыми молекулами РНК (исключая реовиру-сы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине.
Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов). Их молекулярная масса в 10-100 раз меньше массы бактериальных ДНК. Транскрипция вирусной ДНК (синтез мРНК) осуществляется в ядре заражённой вирусом клетки. В вирусной ДНК на концах молекулы имеются прямые или инвертированные (развёрнутые на 180") повторяющиеся нуклеотидные последовательности. Их наличие обеспечивает способность молекулы ДНК замыкаться в кольцо. Эти последовательности, присутствующие в одно- и двух-нитевых молекулах ДНК, — своеобразные маркёры вирусной ДНК.
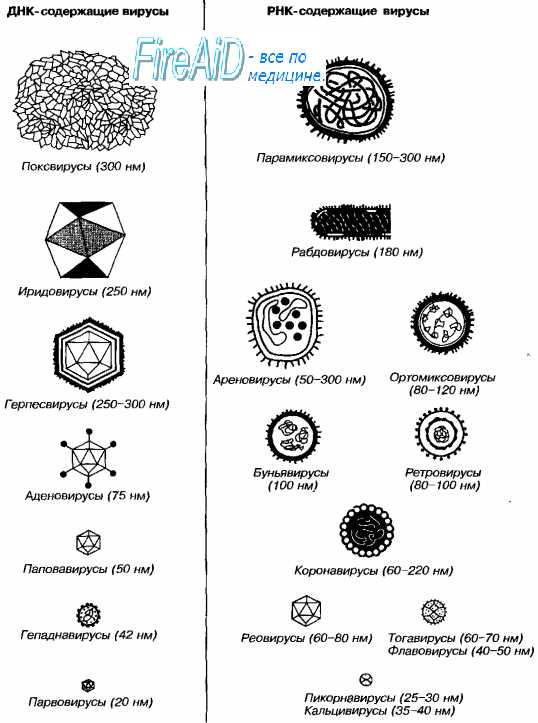
Рис. 2-1. Размеры и морфология основных возбудителей вирусных инфекций человека.
Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. Вирусные РНК подразделяют на следующие группы: плюс-нити РНК (+РНК), минус-нити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК.
Инфекционность нуклеиновых кислот вирусов
Многие вирусные нуклеиновые кислоты инфекционны сами по себе, так как содержат всю генетическую информацию, необходимую для синтеза новых вирусных частиц. Эта информация реализуется после проникновения вириона в чувствительную клетку. Инфекционные свойства проявляют нуклеиновые кислоты большинства +РНК- и ДНК-содержащих вирусов. Двухнитевые РНК и большинство -РНК не проявляют инфекционных свойств.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Читайте также: