
Грегарины жизненный цикл кратко
Обновлено: 03.07.2024
Грегарины (в особенности паразитирующие в кишечнике членистоногих) могут достигать значительных размеров - 16 мм. Самые же мелкие виды не превышают 10-15 мкм. Тело кишечных грегарин обычно продолговатой, червеобразной формы . Грегарины из полости тела могут быть почти сферическими.
Передний конец тела большинства грегарин образует органоид прикрепления к стенкам кишечника - эпимерит . Последний имеет крючки, тонкие выросты в форме нитей и другие образования, позволяющие паразиту закрепиться на месте и не быть вынесенному из кишечника с пищевыми массами.
Снаружи тело одето образующей продольные гребни пелликулой, представляющей наружный плотный слой эктоплазмы. За счет нее и формируются крючки и отростки эпимерита. Под пелликулой залегает слой эктоплазмы, который у многих (но не у всех) грегарин примерно на границе передней трети тела образует волокнистую перегородку, отделяющую передний, лишенный ядра участок цитоплазмы, называемый протомеритом. Задний больший и снабженный ядром участок тела называется дейтомеритом.
Таким образом, многие грегарины, оставаясь одноклеточными, становятся трехчленистыми (эпи-, прото- и дейтомерит). Трехчленистые грегарины объединяются в подотряд Cephalina. Это в основном кишечные формы. Другие (преимущественно полостные, паразиты половых органов и т.п.) не обладают трехчленистостью, и их тело червеобразное или сферическое. Такие грегарины объединяются в подотряд Acephalina.
Лежащая под пелликулой эктоплазма у многих грегарин сложного строения: периферическая часть ее состоит из студенистого слоя, под которым у многих грегарин расположена система кольцевых или продольных волоконец- мионем, имеющих сократительный характер. Наличие их обусловливает способность некоторых грегарин к сокращению и вытягиванию тела.
Полужидкая эндоплазма грегарин всегда очень богата включениями полисахарида гликогена, что связано, очевидно, с тем, что обмен веществ носит у большинства грегарин анаэробный характер, в процессе которого потребляется большое количество углеводов. Ротовое отверстие и порошица отсутствуют. Нет и сократительной вакуоли (как и у всех прочих Sporozoa). Питание и дыхание осуществляются всей поверхностью тела.
Взрослые грегарины, достигшие предельного размера, соединяются попарно, в результате чего образуется так называемый сизигий . Нередко, однако, грегарины объединяются довольно рано, еще на стадиях роста . Тела обоих партнеров в дальнейшем округляются, и вокруг них выделяется плотная оболочка . Слияния грегарин внутри цисты не происходит.
Ядро каждой особи многократно делится митотически. В результате образуется множество ядер, рассеянных в цитоплазме. Эти ядра отходят к периферии тела каждой особи и выпячивают цитоплазму в виде многочисленных бугорков. Далее каждый бугорок отшнуровывается от общей массы цитоплазмы и становится гаметой. При этом часть цитоплазмы первоначально объединившихся в сизигий особей остается неиспользованной (остаточное тело) и в дальнейшем дегенерирует. Гаметы, образовавшиеся в разных особях (гамонтах) одного сизигия, попарно копулируют.
Интересно отметить, что у грегарин (так же как и у колониальных жгутиконосцев ) наблюдаются все переходы от полной изогамии канизогамии . При этом в подотряде Acephalina бывает преимущественно изогамная копуляция, тогда как у Cephalina - разная степень анизогамии.
Продукт копуляции - зигота окружается плотной оболочкой, образуя ооцисту (часто не совсем правильно ее называют спорой). Внутри ооцисты протекает процесс спорогонии: ядро ее, последовательно делясь, дает начало 8 ядрам. Два первых деления представляют собой мейоз и приводят к редукции числа хромосом. У грегарин, как и у всех споровиков, редукция зиготическая. Вслед за образованием 8 ядер цитоплазма ооцисты распадается на 8 мелких червеобразных телец - спорозоитов. Этим заканчивается спорогония и ооциста становится способной к заражению новых особей хозяина.
Цисты с развивающимися в них ооцистами выбрасываются вместе с экскрементами хозяина наружу (или же попадают в наружную среду после смерти хозяина).
Для дальнейшего развития ооцисты должны быть проглочены подходящим для их развития животным. В последнем случае оболочка ооцисты лопается и спорозоиты выходят в просвет кишечника. Там они внедряются своим передним заостренным концом в клетку кишечника, и начинается период роста. Постепенно тело спорозоита, особенно часть его, находящаяся вне эпителиальной клетки, удлиняется, формируется эпимерит, появляется перегородка, делящая животное на протомерит и дейтомерит. Растущий спорозоит постепенно принимает облик взрослой грегарины. Паразитируя лишь в беспозвоночных, грегарины не имеют большого практического значения.
Грегарины — большая группа споровиков, включающая около 500 видов. Все без исключения грегарины — паразиты беспозвоночных. Наибольшее число видов их обитает в кишечнике членистоногих, в особенности часто у насекомых. Некоторые грегарины паразитируют и в других группах беспозвоночных — в кольчатых червях, иглокожих, оболочниках. Кроме кишечных грегарин, каковых большинство, имеются виды, живущие в полости тела, в органах размножения и др.
Наиболее сложно устроены грегарины, живущие в кишечнике членистоногих. На их примере мы и ознакомимся с организацией представителей этого отряда.


Разделение цитоплазмы грегарин на два слоя — эктоплазму и эндоплазму — выражено всегда очень отчетливо. Наружный слой эктоплазмы образует прочную пелликулу, определяющую постоянство формы тела грегарин. У некоторых грегарин в эктоплазме найдены тонкие скелетно-опорные волоконца, еще более увеличивающие прочность наружных слоев тела. Немногие виды грегарин способны несколько менять форму своего тела, сокращаться вдоль продольной оси. У таких видов в эктоплазме, на границе с эндоплазмой, найдены сократимые волоконца — миофибриллы.
Ядра, расположенные всегда в дейтомерите, имеют обычно вид пузырька, богатого ядерным соком и снабженного одним или несколькими крупными ядрышками.
Рассмотренное выше строение грегарины свойственно преимущественно кишечным паразитам. Оно характерно для группы видов, объединяемых в семейство Polycystidae. У представителей другого семейства — Monocystidae, которые паразитируют главным образом в полости тела, половых железах и других органах, строение более простое. У Monocystidae нет разделения на эпи-, прото- и дейтомерит, и тело их часто имеет червеобразную форму.
Наблюдая под микроскопом за живыми грегаринами, легко обнаружить, что они способны двигаться, медленно перемещаясь в содержимом кишечника. Это движение носит характер плавного скольжения, не сопровождающегося изменением формы тела.
Механизм этого процесса до настоящего времени не вполне ясен, поскольку у грегарин нет никаких специальных органоидов движения. Исследования Шевякова и других авторов показали, что на заднем конце движущейся грегарины выделяется слизь. По-видимому, она выбрасывается наружу с известной силой, благодаря чему и происходит — по принципу ракетного двигателя — поступательное движение грегарины.
У настоящих грегарин отсутствует бесполое размножение, они размножаются только половым путем. Рассмотрим типичный цикл развития грегарин (рис. 69).

Живущие в кишечнике насекомого грегарины, достигнув предельного роста, теряют эпимерит и соединяются попарно. Вокруг такой парочки (сизигия) выделяется общая оболочка (циста). Вскоре после инцистирования в каждой из соединившихся в сизигий особи начинается многократное последовательное деление ядра. В каждой особи образуется множество ядер. К концу этого процесса все ядра располагаются по периферии грегарины и вокруг каждого из них обособляется небольшой участок цитоплазмы. Так формируется множество мелких одноядерных клеток. Большая часть цитоплазмы при этом остается неиспользованной. Это так называемое остаточное тело, которое в дальнейшем процессе размножения участия не принимает. Мелкие одноядерные клетки представляют собой половые клетки — гаметы. Эти клетки сближаются попарно и сливаются.
У некоторых видов грегарин гаметы, образуемые обоими соединившимися в сизигий особями, по размерам и строению неотличимы друг от друга. У других видов они различны. Одна особь дает начало округлым и неподвижным гаметам. Это женские гаметы. Другая особь образует удлиненные гаметы, снабженные жгутиковидным выростом. Это мужские подвижные гаметы.
Образовавшиеся в результате оплодотворения зиготы сразу же покрываются оболочкой, принимая при этом веретенообразную, а нередко бочонковидную форму. Покрывшаяся оболочкой зигота носит название ооцисты. Оболочка цисты в этот период цикла развития еще не разрушается. Таким образом, многочисленные образовавшиеся после копуляции гамет ооцисты лежат внутри оболочки цисты. На этой стадии, а иногда и несколько позже, цисты с ооцистами выводятся из кишечника наружу. Последним этапом цикла развития грегарины является процесс формирования ооцисты, получивший название спорогонии. Он сводится к следующему. Внутри ооцисты ядро (образовавшееся в результате слияния ядер гамет) трижды делится. Вокруг каждого из возникших таким путем ядер обособляется участок цитоплазмы. В каждой ооцисте получается восемь очень маленьких червеобразной формы клеток, называемых спорозоитами. На этой стадии ооцисты способны к заражению новых особей хозяина. Если циста с ооцистами или даже отдельная ооциста будет проглочена насекомым — хозяином данного вида грегарин, то под воздействием кишечного сока спорозоиты выходят из ооцисты. Они подвижны. В дальнейшем из каждого спорозоита развивается грегарина. Достигнув типичного для данного вида размера, грегарины вновь соединяются попарно, инцистируются, и цикл начинается снова.
Таким образом, рассмотренный выше цикл развития грегарин можно отчетливо разделить на три этапа. Первый — это рост и развитие взрослой грегарины. Второй — образование половых клеток (гамет) и оплодотворение. Третьим этапом цикла является спорогония — развитие зиготы (ооцисты) и формирование спорозоитов.
Жизнь животных: в 6-ти томах. — М.: Просвещение. Под редакцией профессоров Н.А.Гладкова, А.В.Михеева . 1970 .
Грегарины — большая группа споровиков, включающая около 500 видов. Все без исключения грегарины — паразиты беспозвоночных. Наибольшее число видов их обитает в кишечнике членистоногих, в особенности часто у насекомых. Некоторые грегарины паразитируют и в других группах беспозвоночных — в кольчатых червях, иглокожих, оболочниках.
Кроме кишечных грегарин, каковых большинство, имеются виды, живущие в полости тела, в органах размножения и др. Строение. Наиболее сложно устроены грегарины, живущие в кишечнике членистоногих. На их примере мы и ознакомимся с организацией представителей этого отряда.

Он отделен от дейтомерита тонкой прослойкой светлой цитоплазмы, представляющей собой продолжение слоя эктоплазмы. На самом переднем конце грегарины расположен эпимерит. Этот участок тела служит для прикрепления грегарины к стенке кишечника. Часто он бывает снабжен крючьями, иногда нитевидными выростами и может принимать весьма причудливую форму. На рисунке 56 изображены некоторые характерные формы эпимеритов разных видов грегарин.

По достижении предельных размеров, когда у грегарины наступают процессы полового размножения, эпимерит обычно отбрасывается. Он целиком состоит из эктоплазмы. Разделение цитоплазмы грегарин на два слоя— эктоплазму и эндоплазму — выражено всегда очень отчетливо. Наружный слой эктоплазмы образует прочную пелликулу, определяющую постоянство формы тела грегарин. У некоторых грегарин в эктоплазме найдены тонкие скелетно-опорные волоконца, еще более увеличивающие прочность наружных слоев тела.
Немногие виды грегарин способны несколько менять форму своего тела, сокращаться вдоль продольной оси. У таких видов в эктоплазме, на границе с эндоплазмой, найдены сократимые волоконца — миофибриллы. Эндоплазма грегарины, резко отграниченная от эктоплазмы,; при рассматривании под микроскопом живых грегарин выглядит темно-серой. Это обусловлено наличием в эндоплазме большого количества гликогена, нередко сконцентрированного в виде мелких гранул.
В этом проявляется одна из характерных особенностей многих внутренностных паразитов, тело которых богато гликогеном. Кишечные паразиты живут в среде, в которой свободный кислород присутствует в ничтожных количествах. Это затрудняет или делает невозможными дыхательные процессы, сопровождающиеся поглощением кислорода. В этих условиях паразитические организмы вынуждены искать взамен дыхания другие источники энергии.

Такими источниками оказываются процессы анаэробного (без участия свободного кислорода) расщепления гликогена, процессы гликолиза. Не вдаваясь в химическую характеристику этих процессов, отметим лишь, что при этом сложная молекула гликогена распадается на более простые органические соединения (в частности, некоторые органические кислоты).
Рассмотренное выше строение грегарины свойственно преимущественно кишечным паразитам. Оно характерно для группы видов, объединяемых в семейство Polycystidae. У представителей другого семейства — Monocystidae, которые паразитируют главным образом в полости тела, половых железах и других органах, строение более простое. У Monocystidae нет разделения на эпи-, прото- и дейтомерит, и тело их часто имеет червеобразную форму.
Наблюдая под микроскопом за живыми грегаринами, легко обнаружить, что они способны двигаться, медленно перемещаясь в содержимом кишечника. Это движение носит характер плавного скольжения,; не сопровождающегося изменением формы тела. Механизм этого движения до настоящего времени не вполне ясен. По современным представлениям, он связан с имеющимися на наружной поверхности грегарин продольно идущими гребнями пелликулы. Эти гребни совершают периодические волнообразные движения, которые обусловливают медленное скользящее перемещение тела грегарины.
Размножение. У настоящих грегарин отсутствует бесполое размножение, они размножаются только половым путем. Рассмотрим типичный цикл развития грегарин (рис. 57). Живущие в кишечнике насекомого грегарины, достигнув предельного роста, теряют эпимерит и соединяются попарно. Вокруг такой пары (сизигия) выделяется общая оболочка (циста).
Вскоре после инцистирования в каждой из соединившихся в сизигий особи начинается многократное последовательное деление ядра. В каждой особи образуется множество ядер. К концу этого процесса все ядра располагаются по периферии грегарины и вокруг каждого из них обособляется небольшой участок цитоплазмы. Так формируется множество мелких одноядерных клеток. Большая часть цитоплазмы при этом остается неиспользованной. Это так называемое остаточное тело, которое в дальнейшем процессе размножения участия не принимает.
Мелкие одноядерные клетки представляют собой половые клетки — гаметы. Эти клетки сближаются попарно и сливаются. У некоторых видов грегарин гаметы, образуемые обоими соединившимися в сизигий особями, по размерам и строению неотличимы друг от друга. У других видов они различны. Одна особь дает начало округлым и неподвижным гаметам. Это женские гаметы. Другая особь образует удлиненные гаметы, снабженные жгутиковидным выростом. Это мужские подвижные гаметы.
Образовавшиеся в результате оплодотворения зиготы сразу же покрываются оболочкой, принимая при этом веретенообразную, а нередко бочонковидную форму. Покрывшаяся оболочкой зигота носит название о о ц и с т ы. Оболочка цисты в этот период цикла развития еще не разрушается. Таким образом, многочисленные образовавшиеся после копуляции гамет ооцисты лежат внутри оболочки цисты. На этой стадии, а иногда и несколько позже, цисты с ооцистами выводятся из кишечника наружу.

Последним этапом цикла развития грегарины является процесс размножения ооцисты, получивший название спорогонии. Он сводится к следующему. Внутри ооцисты ядро (образовавшееся в результате слияния ядер гамет) трижды делится. Вокруг каждого из возникших таким путем ядер обособляется участок цитоплазмы. В каждой ооцисте получается восемь очень маленьких червеобразной формы клеток, называемых спорозоитами. На этой стадии ооцисты способны к заражению новых особей хозяина.
Если циста с ооцистами или даже отдельная ооциста будет проглочена насекомым — хозяином данного вида грегарин, то под воздействием кишечного сока спорозоиты выходят из ооцисты. Они подвижны. В дальнейшем из каждого спорозоита развивается грегарина. Достигнув типичного для данного вида размера, грегарины вновь соединяются попарно, инцистируются, и цикл начинается снова. Таким образом, рассмотренный выше цикл развития грегарин можно отчетливо разделить на три этапа: рост и развитие взрослой грегарины; образование гамет; оплодотворение. Последним этапом цикла является спорогония — развитие зиготы (ооцисты) и формирование спорозоитов.
Класс Споровики (Sporozoea) включает отряды, отряд Грегарины (Gregarinida), отряд кокцидии (Coccidia).
Отряд Грегарины (Gregarinida). Грегарины - паразиты беспозвоночных животных Известно более 500 видов грегарин Среди них встречаются крупные виды размером до 16 мм и мелкие внутриклеточные паразиты (10- 15 мкм).
В жизненном цикле грегарин своеобразен процесс полового размножения, при котором два гамонта соединяются в сизигий, а затем покрываются общей оболочкой, образуя цисту Бесполое размножение - шизогония может отсутствовать К подклассу грегарин относится несколько подотрядов, из которых мы познакомимся лишь с одним, наиболее многочисленным - собственно грегаринами (Eugregarinina).
Хозяевами грегарин в основном являются насекомые, а также черви, реже водные моллюски, иглокожие.
Большинство грегарин - внутрикишечные паразиты беспозвоночных животных, меньшее число видов паразитируют в полости тела или в гонадах. Кишечные формы грегарин более сложны по строению. Так, у грегарины Corycella armata из кишечника жука-вертячки (Gyrinus natator) тело состоит из трех отделов: эпимерита, протомерита и дейтомерита, а у Gregarina cuneata из кишечника жука мучного хруща (Tenebrio molitor) - из двух отделов: прото- и дейтомерита. Эпимерит служит для прикрепления к стенке кишки и нередко снабжен крючьями. Прото- и дейтомерит разделены между собой слоем прозрачной эктоплазмы. В дейтомерите расположено ядро. Эндоплазма грегарин перегружена зернами парагликогена - запасного энергетического материала Грегарины - эндопаразиты и характеризуются анаэробным, т. е. бескислородным, дыханием, при котором парагликоген расщепляется на более простые вещества с выделением энергии, необходимой для обменных процессов. Тело грегарин, обитающих в гонадах и других внутренних органах, не подразделяется на отделы и имеет червеобразную или сферическую форму. Пелликула грегарин плотная, что определяет их относительно постоянную форму тела. Под пелликулой у некоторых грегарин обнаружены кольцевые и продольные мионемы - сократительные волоконца. Их сокращение обеспечивает способность к медленному движению в плотной жидкости. Движению грегарин может также способствовать ундуляция пелликулы Питаются грегарины сапрофитно, впитывая органические вещества всей поверхностью клетки.
Рассмотрим для примера жизненный цикл грегарины Stylocephalus longicol из кишечника жука-чернотелки (рис. 45).
Перед размножением грегарины соединяются попарно в цепочку (сизигий) В дальнейшем они округляются и покрываются общей оболочкой - цистой. Ядро каждого партнера претерпевает многократное деление Вокруг ядер обособляется цитоплазма и образуются гаметы. Гаметы партнеров могут быть одинаковыми или разными по размеру, т. е. наблюдается изо- или анизогамия. Микрогамета со жгутиком. Часть цитоплазмы от грегарин остается в виде остаточного тела, которое в дальнейшем расходуется как питательный материал для развивающихся зигот. После копуляции гамет партнеров образуются зиготы, которые покрываются плотной оболочкой и формируются ооцисты. Цисты с ооцистами выходят из, кишечника наружу. Их дальнейшее развитие происходит во внешней кислородной среде. Внутри ооцисты ядро зиготы несколько раз делится
и затем образуются узкие клетки - спорозоиты. Этот процесс размножения ооцисты получил название спорогонии. В процессе спорогонии происходит редукционное деление. После образования спорозоитов ооцисты становятся инвазийными, т. е. способными к заражению других особей жуков-чернотелок. Жуки вместе с пищей заглатывают ооцисты грегарины и заражаются паразитами. Под действием пищеварительных соков жука оболочка ооцисты растворяется и спорозоиты выходят в полость кишечника. Они внедряются в клетки кишечника и некоторое время развиваются внутриклеточно. При дальнейшем росте они разрывают клетку кишечника и вырастают в крупную грегарину - внутриполостного паразита с трехчленностью строения.
Таким образом, рассмотренный нами жизненный цикл грегарины характеризуется тем, что в теле хозяина происходит только половое размножение (гамогония), а в кислородной среде происходит спорогония - с образованием спорозоитов. Инвазия паразита - кишечная. Хозяин заражается грегариной при поедании цист с ооцистами и спорозоитами. У грегарин, как и у всех споровиков, в жизненном цикле доминирует фаза гаплонта. Диплонтом является лишь зигота. Ее первое же деление - мейоз, и потому образующиеся молодые фазы споровиков - спорозоиты уже гаплоидны. Таким образом, грегарины - гаплонты с зиготической редукцией хромосом (рис. 46).
Отряд Кокцидии (Coccidia). Кокцидии - внутриклеточные паразиты, в основном позвоночных и редко беспозвоночных животных. Всего известно более 400 видов этого подкласса. Клетка кокцидий округлая, недифференцированная на отделы, как у грегарин. Это в основном очень мелкие формы, размеры которых достигают всего нескольких микрометров. Отряд включает несколько подотрядов: подотряд Эймериевые (Eimeriina), подотряд Кровяные споровики (Haemosporina), подотряд Пироплазмы (Piroplasmina).
Подотряд Эймериевые (Eimeriina). Эймериевые паразитируют только у позвоночных животных, преимущественно у млекопитающих и птиц. Заболевания, вызываемые кокцидиями, называются кокцидиозами. Кокцидиозам подвержены главным образом молодые животные. От кокцидиоза наиболее часто страдают кролики, овцы, телята, куры. Кокцидии паразитируют в клетках стенок кишечника и вызывают кровавый понос, изнуряющий организм хозяина.
Рассмотрим жизненный цикл эймериевых кокцидий на примере Eimeria magna - возбудителя кокцидиоза у кроликов (рис. 47). Кролики заражаются кокцидиозом, проглотив вместе с пищей ооцисты Eimeria magna. В кишечнике из ооцист выходят спорозоиты, внедряющиеся в клетки стенки кишки. Питающаяся фаза кокцидий называется трофозоитом. Ядро трофозоита начинает многократно делиться и формируется многоядерная форма - шизонт (агамонт), приступающий к бесполому
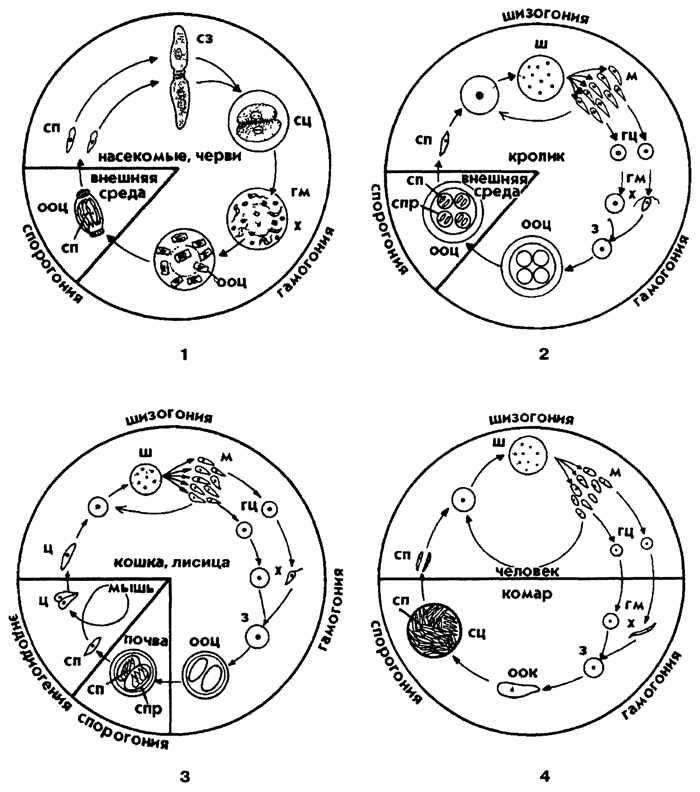
Рис. 46. Схемы жизненных циклов споровиков: 1 - грегарины, 2 - кокцидий, 3 - токсоплазмы, 4 - малярийного плазмодия. Обозначения: гм - гаметы, гц - гаметоциты, з - зигота, м - мерозоиты, ооц - ооциста, сз - сизигий, сп - спорозоит, спр - спора, сц - спороциста, ш - шизонт, ц – цистозоит
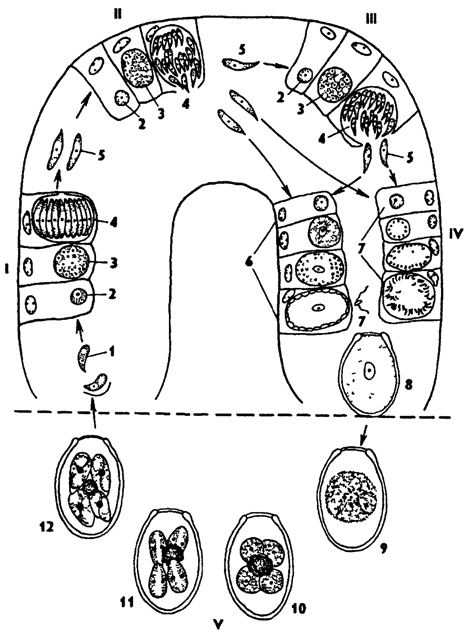
Рис. 47. Жизненный цикл кокцидий рода Eimeria (по схеме Хайсина): I - первое поколение шизогонии, II - второе поколение шизогонии, III - третье поколение шизогонии, IV - гамогония, V - спорогония; 1 - спорозоиты, 2 - одноядерный шизоит, 3 - многоядерный шизоит, 4 - образование мерозоитов, 5 - мерозоиты, 6 - развитие микрогамет, 7 - развитие микрогамет, 8 - ооциста, 9, 10 - образование споробластов (видно остаточное тело), 11 - образование спор, 12 - зрелая ооциста с четырьмя спорами, в каждой споре по два спорозоита
размножению, шизогонии (агамогонии). В результате шизогонии образуются десятки мелких узких клеток - мерозоитов. Пораженная клетка хозяина разрушается, и из нее мерозоиты выходят в полость кишечника. Они поражают здоровые клетки, и цикл шизогонии повторяется. У Eimeria magna наблюдается пять генераций мерозоитов. Последняя генерация мерозоитов преобразуется в клетках кишечника в гамонтов. Одни гамонты (микрогамонты) образуют путем деления множество гамет со жгутиками (микрогамет) Другие - макрогамонты - не делятся, и каждый из них преобразуется в одну макрогамету, соответствующую яйцеклетке.
Микрогаметы выходят в полость кишечника, проникают к макрогамете. После копуляции гамет образуется зигота, покрывающаяся оболочкой - ооциста. Ооцисты выносятся из кишечника наружу. В кислородной среде в ооцистах происходит процесс спорогонии. Вначале образуются четыре клетки - споробласты, покрывающиеся оболочкой, и из них формируются споры. В каждой споре споробласт образует два спорозоита. После завершения спорогонии споры становятся инвазийными, т. е. способными к заражению животных.
Таким образом, в жизненном цикле кокцидий - Eimeria magna наблюдается пять поколений шизонтов (агамонтов), одно поколение гамонтов и несколько поколений в результате деления зиготы (спорогония). После завершения всего жизненного цикла эймерии, на который уходит 175- 208 ч, организм кролика освобождается от паразита. При борьбе с кокцидиозом необходимо принимать меры по предупреждению повторной инвазии. Кокцидиозами заражаются многие домашние животные. Лечение больных животных, их изоляция от здоровых, соблюдение санитарных норм содержания помещений для животных, хороший уход обеспечивают успешную борьбу с кокцидиозами.
Среди эймериевых кокцидий опасность для человека представляет токсоплазма (Toxoplasma gondii). Заболевание, вызываемое этим паразитом, называется токсоплазмозом, который широко распространен по всему миру.
Жизненный цикл токсоплазмы похож на таковой у эймерии, но усложнен сменой хозяев и появлением дополнительных форм размножения (рис. 48).
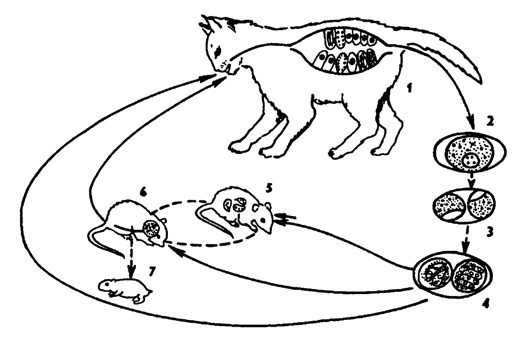
Рис. 48. Жизненный цикл Toxoplasma gondii (по Френкелю): 1 - кошка - окончательный хозяин, в котором паразит проходит шизогонию и стадии полового цикла, 2- 4 - стадии развития ооцисты с образованием двух спор в каждой с четырьмя спорозоитами, 5, 6 - промежуточный хозяин, в котором происходит дополнительное бесполое размножение паразита (эндодиогения) и образование цисты с цистозоитами, 7 - внутриутробное заражение мышей
Основным хозяином токсоплазмы являются кошки, в кишечнике которых паразиты размножаются путем шизогонии, а затем половым путем (гамогония) с образованием ооцист. В дальнейшем ооцисты развиваются во внешней кислородной среде, и в результате спорогонии в них формируется по две споры с четырьмя спорозоитами. Промежуточным хозяином токсоплазмы могут быть любые виды птиц и млекопитающих, в том числе и человек. Заражение промежуточных хозяев происходит путем заглатывания цист токсоплазмы с загрязненной пищей или водой. Особенно опасны для заражения человека токсоплазмозом контакты с кошками. Основной хозяин - кошка заражается, поедая некоторых промежуточных хозяев. Наиболее обычным промежуточным хозяином для Toxoplasma являются мыши.
В кишечнике промежуточного хозяина, к которым относится и человек, оболочки ооцист и спор растворяются и из них выходят спорозоиты, внедряющиеся в ткани и попадающие в кровяные сосуды. Паразиты могут локализоваться в любых органах, в том числе мышцах, печени, мозге, глазах. В местах локализации паразиты размножаются путем эндодиогении. Это особый способ бесполого размножения, когда дочерние клетки образуются внутри материнской и лишь позднее обособляются (рис. 49). В результате эндодиогении образуется скопление паразитов, выделяющих вокруг себя уплотненную оболочку. Это цисты токсоплазмы, внутри которых сосредоточены цистозоиты серповидной формы.
Токсоплазма может через плаценту передаваться плоду млекопитающих и человека, что вызывает обычно гибель потомства (трансплацентарная инвазия).
Токсоплазмозы у человека могут протекать как в легкой форме и даже быть незамеченными, так и в тяжелой - с летальным исходом. Отмечено, что у людей, зараженных СПИДом, часто активизируется токсоплазмоз, нередко приводящий человека к гибели.
В борьбе с токсоплазмозом проводится обследование людей, проводится лечение больных и осуществляются профилактические мероприятия. Соблюдение санитарных норм, гигиена питания, осторожность по отношению к бродячим кошкам предохраняют человека от заражения токсоплазмозом.
Рис. 49. Эндодиогения у кокцидий (схема по Шолтизеку): 1 - формирование дочерней клетки
Подотряд Кровяные споровики (Haemosporina). Кровяные споровики - специализированные внутриклеточные паразиты крови млекопитающих, птиц и рептилий. Эти паразиты поражают эритроциты крови. Некоторые виды рода Plasmodium паразитируют у человека, вызывая опасную болезнь малярию. Только в конце XIX в. французским врачом Лавераном был обнаружен ее возбудитель - малярийный плазмодий в крови человека, а англичанином Россом обнаружены цисты со спорозоитами из желудка малярийного комара.
Полностью жизненный цикл малярийного плазмодия описан итальянским зоологом Грасси. Приоритет в разработке противоэпидемических мероприятий по борьбе с малярией принадлежит отечественным ученым: Е. И. Марциновскому, Л. М. Исаеву, Е. Н. Павловскому, В. Н. Беклемишеву, Н. И. Латышеву.
Жизненный цикл малярийного плазмодия (Plasmodium vivax) характеризуется сменой хозяев и чередованием поколений с половым и бесполым размножением (рис. 50). Перенос паразита осуществляется малярийными комарами рода Anopheles, которые являются окончательными хозяевами плазмодия.
Человек - промежуточный хозяин малярийного плазмодия. Заражение происходит при укусе комара, в слюне которого содержатся спорозоиты. Вначале спорозоиты внедряются в паренхимные клетки печени и размножаются путем шизогонии. Так происходит накопление паразита в крови, после чего мерозоиты внедряются в эритроциты. В процессе развития плазмодий проходит фазу трофозоита, а затем многоядерного шизонта. Пораженные эритроциты разрушаются, и мерозоиты выходят в плазму крови и внедряются в другие эритроциты. Продолжительность одного цикла шизогонии видоспецифична. Так, у P. vivax и P. falciparum цикл шизогонии длится 48 ч, у P. malariae - 72 ч. Завершение шизогони и выход мерозоитов из эритроцитов сопровождается у больного повышением температуры и лихорадкой. Это связано с тем, что из разрушенных эритроцитов в кровь поступают продукты диссимиляции паразита (меланины и др.), вызывающие интоксикацию. После нескольких циклов шизогонии болезненные явления прекращаются, а паразиты развиваются в покоящуюся фазу - гамонтов. Человек становится носителем малярийного паразита.
У комара, напившегося крови больного малярией, продолжается развитие плазмодия (гамонтов). В кишечнике комара происходит гамогония. Из микрогамонта образуются узкие мужские гаметы (4- 8), а из макрогамонта формируется одна крупная макрогамета (яйцеклетка). После копуляции гамет образуется зигота - червеобразная оокинета, которая внедряется в стенку кишки. На внешней поверхности кишечника оокинета преобразуется в цисту, покрытую тонкой оболочкой. В цисте происходит
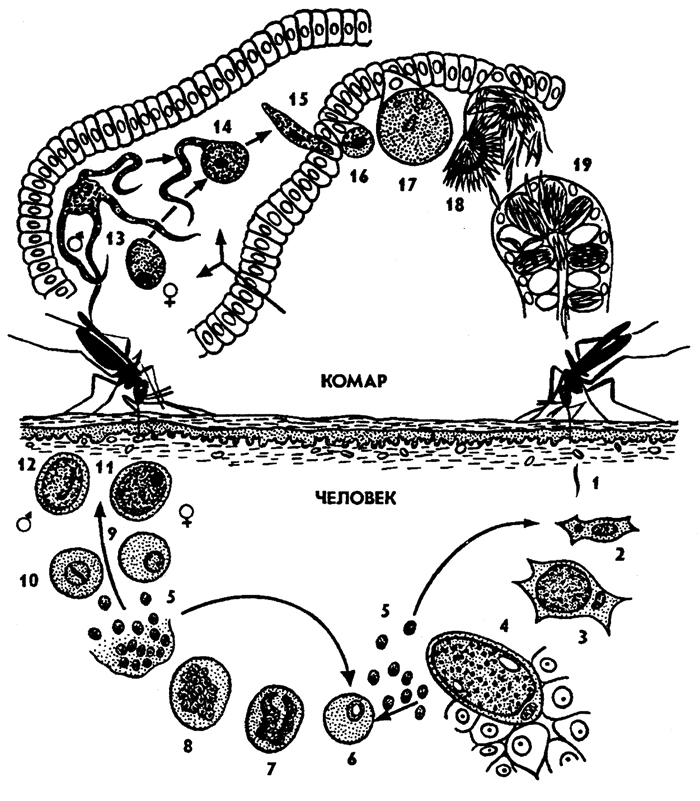
Рис. 50. Жизненный цикл малярийного плазмодия рода Plasmodium (по Хаусману): 1 - спорозоит, 2,3 - рост шизонта (агамонта), 4 - шизогония в клетках печени, 5 - мерозоиты, 6- 8 - эритроцитная шизогония, 9- 12 - образование гамонтов (микро- и макрогамонтов), 13 - образование макрогамет и микрогамет, 14 - копуляция гамет, 15 - зигота (оокинета), 16- 18 - спорогония и образование спороцисты со спорозоитами, 19 - накопление спорозоитов в слюнных железах комара
спорогония паразита с образованием множества спорозоитов (до 500). После разрыва стенки цисты спорозоиты по руслу гемолимфы комара попадают в слюнные железы, где происходит их накопление. При укусе зараженным малярийным комаром в кровь человека попадают спорозоиты. У кровяных споровиков в отличие от кокцидий споры не образуются в связи с тем, что паразит распространяется с помощью переносчика (трансмиссивно).
Малярия и борьба с ней. Малярия - тяжелое трансмиссивное заболевание человека, вызываемое кровяными споровиками рода Plasmodium. Поражение эритроцитов крови человека плазмодиями вызывает малокровие, анемию, интоксикацию, увеличение селезенки. Малярия сопровождается лихорадкой. Течение болезни зависит от видов малярийных плазмодиев. Наиболее распространена трехдневная малярия, возбуждаемая P. vivax (48 ч), которая распространена в регионах с умеренным и тропическим климатом. Четырехдневная малярия, вызываемая P. malariae (приступы через 72 ч), чаще встречается в южных широтах.
Переносят малярию комары рода Anopheles, главным образом вид A. maculipennis. Относительно недавно выявлено, что существует шесть видов-двойников малярийного комара в Европе, которых ранее объединяли в один вид A. maculipennis. Малярийные комары отличаются от наиболее часто встречающихся комаров родов Culex и Aeles по морфологии и поведению имаго и личинок.
Всемирной организацией здравоохранения были разработаны, а затем реализованы меры борьбы с малярией. Во многих странах мира малярия в значительной степени ликвидирована (страны Европы, США и др.). Успехи борьбы с малярией в нашей стране связаны с активной работой Института тропической медицины, Военно-медицинской академии и ряда противоэпидемиологических станций. К 1959 г. были ликвидированы основные очаги малярий. Однако и сейчас в некоторых регионах Закавказья, Средней Азии требуется постоянный контроль и проведение противомалярийных мероприятий.
К мерам борьбы с малярией относятся: обследование населения на заряженность малярийными плазмодиями и их лечение, борьба с малярийными комарами и ликвидация очагов их выплода (временные водоемы, сырые подвалы), а также профилактические мероприятия по предупреждению инвазии.
Подотряд Пироплазмы (Piroplasmina). Пироплазмы паразитируют в эритроцитах крови жвачных животных и вызывают тяжелые заболевания - пироплазмозы, нередко вызывающие летальный исход. Переносчиками пироплазмозов являются иксодовые клещи. Особенно опасные заболевания вызывают пироплазмы рода Babesia. Например, техасскую лихорадку рогатого скота вызывает В. bigemina, случаи которой встречаются в Средней Азии и на Кавказе. Переносчиком болезни является клещ Margaporus. Другие пироплазмы вызывают заболевания лошадей, овец. К профилактическим мероприятиям относятся: борьба с переносчиками пироплазм, лечение больных животных, проведение карантина.
Читайте также: