
Горизонтальный перенос генов у бактерий кратко
Обновлено: 06.07.2024
Долгое время эволюционная теор ия базировалась на представлении о том, что виды не могут обмениваться друг с другом наследственной информацией. Как только вид делится на два, и между ними возникает репродуктивная изоляция, они эволюционируют дальше сами по себе по схеме "случайные мутации + естественный отбор". На этом основывалась и Дарвиновская схема дивергенции, и столь модный на Западе кладизм – "филогенетическая систематика", и все "эволюционные деревья" и иные реконструкции исторического развития биоты. Однако расчеты показывали, что при такой изолированной эволюции отдельных видов на основе случайных мутаций и отбора жизнь просто не успела бы за сравнительно недолгий срок своего существования (4 млрд. лет) развиться от простейших форм до таких высокоорганизованных, как млекопитающие и человек.
С открытием горизонтального переноса генов между разными видами и даже царствами живых организмов (в последнее время получено множество доказательств, см. ниже) ситуация изменилась, и эволюция предстает в совершенно ином свете. Получается, что любое "удачное изобретение" одного из видов становится доступным и может быть заимствовано всеми остальными. Биосфера теперь представляется единой информационной средой, в которой вирусы и различные мобильные генетические элементы распространяют информацию примерно так же, как в человеческом обществе благодаря устной и письменной речи достижения и открытия одних людей становятся известными другим и могут ими использоваться.
Конечно, этот процесс горизонтального обмена вовсе не является бесконтрольным и неорганиченным. Он более-менее таков только у прокариот, которые действительно обмениваются генами достаточно свободно и в каком-то смысл е могут рассматриваться как единый, огромный и невероятно полиморфный вид. Эукариоты выработали специальные сложные адаптации для того, чтобы ограничивать и контролировать этот процесс. Важнейшими из этих адаптаций являются половое размножение и репродуктивная изоляция видов (изоляция, конечно, тоже не абсолютная); собственно, именно появление полового размножения и репродуктивной изоляции и привело к формированию нового класса биологических систем – эндогамных видов . Но и эукариоты способны заимствовать чужие гены. Например, показано, как анаэробная паразитическая амеба (эукариотический организм) "позаимствовала" у бактерий гены ферментов анаэробного метаболизма (см. ниже в подборке рефератов).
Если уж у Бога (или Природы) была возможность создать механизм горизонтального переноса и тем самым колоссально ускорить прогрессивную эволюцию, поскольку вся биота тем самым фактически превращается в единую "лабораторию" по изобретению новых полезных наследственных признаков, то просто глупо было бы отказаться от такого механизма интеграции или полностью запретить его у высших организмов.
Т.И.Тихоненко. В поисках третьего фактора. Вирусолог о горизонтальном переносе как важнейшем факторе эволюции, ускоряющем прогресс.
Н.А.Колчанов. Эволюция регуляторных генетических систем. (там, в частности, говорится об огромном эволюционном потенциал е горизонтального обмена регуляторными генами)
Горизонтальный перенос генов у архей и бактерий ( из доклада С.В.Шестакова )
число генов в геном е
АРХЕИ
Archaeoglobus fulgidus
Methanococcus jannaschii
Pyrococcus horikoshii
Aeropyrum pernix
ПАТОГЕННЫЕ БАКТЕРИИ
Mycoplasma pneumoniae
Chlamydia trachomatis
Rickettsia prowazekii
Treponema pallidum
Haemophilus influenzae
Helicobacter pylori
Mycobacterium tuberculosis
СВОБОДНОЖИВУЩИЕ БАКТЕРИИ
Aquifex aeolicus
Thermotoga maritima
Escherichia coli
Pseudomonas aeruginosa
Bacillus subtilis
Synechocystis sp.
Из обзора: Koonin E.V., Makarova K.S., Arvind L. 2001. Annual Rev. Microbiol. v.55: 709-42
Рефераты статей
Lateral gene transfers and the evolution of eukaryotes: theories and data.
Katz LA.
Int J Syst Evol Microbiol 2002 Sep;52(Pt 5):1893-900
Vertical transmission of heritable material, a cornerstone of the Darwinian theory of evolution, is inadequate to describe the evolution of eukaryotes, particularly microbial eukaryotes. This is because eukaryotic cells and eukaryotic genomes are chimeric, having evolved through a combination of vertical (parent to offspring) and lateral (trans-species) transmission. Observations on widespread chimerism in eukaryotes have led to new and revised hypothesis for the origin and diversification of eukaryotes that provide specific predictions on the tempo (early vs continuous transfers) and mode (nature of donor and recipient lineages) of lateral gene transfers (LGTs). Analyses of available data indicate that LGTs in eukaryotes largely fall into two categories: (1) LGTs from organelles to the nucleus, only a few of which appear to have occurred at the time of the origin of eukaryotes, and (2) anomalous LGTs involving diverse donor and recipient lineages. Further testing of hypotheses on the origin and diversification of eukaryotes will require complete genome sequences from a number of diverse eukaryotes and prokaryotes combined with sequences of targeted genes from a broad phylogenetic sample.
возможно, образование ядра и особой сложной системы регуляции геном а было обусловлено становлением симбиоза (анаэробная клетка с аэробными симбионтами). Нужно ведь было теперь регулировать не 1, а 2 разных геном а, чтобы они работали слаженно! Кардинально облегчить задачу впоследствии помог переход бол-ства митохондриальных генов в ядро, где они могли более эффективно регулироваться.
Microbial gene transfer: an ecological perspective.
Paul JH.
J Mol Microbiol Biotechnol 1999 Aug;1(1):45-50
Microbial gene transfer or microbial sex is a means of exchanging loci amongst prokaryotes and certain eukaryotes. Historically viewed as a laboratory artifact, recent evidence from natural populations as well as genome research has indicated that this process may be a major driving force in microbial evolution. Studies with natural populations have taken two approaches- either adding a defined donor with a traceable gene to an indigenous community, and detecting the target gene in the indigenous bacteria, or by adding a model recipient to capture genes being transferred from the ambient microbial flora. However, both approaches usually require some cultivation of the recipient, which may result in a dramatic underestimation of the ambient transfer frequency. Novel methods are just evolving to study in situ gene transfer processes, including the use of green fluorescent protein (GFP)-marked plasmids, which enable detection of transferrants by epifluorescence microscopy. A transduction-like mechanism of transfer from viral-like particles produced by marine bacteria and thermal spring bacteria to Escherichia coil has been documented recently, indicating that broad host range transduction may be occurring in aquatic environments. The sequencing of complete microbial genomes has shown that they are a mosaic of ancestral chromosomal genes interspersed with recently transferred operons that encode peripheral functions. Archaeal genomes indicate that the genes for replication, transcription, and translation are all eukaryotic in complexity, while the genes for intermediary metabolism are purely bacterial. And in eukaryotes, many ancestral eukaryotic genes have been replaced by bacterial genes believed derived from food sources. Collectively these results indicate that microbial sex can result in the dispersal of loci in contemporary microbial populations as well as having shaped the phylogenies of microbes from multiple, very early gene transfer events.
Evolution of cyanobacteria by exchange of genetic material among phyletically related strains.
Rudi K, Skulberg OM, Jakobsen KS.
J Bacteriol 1998 Jul;180(13):3453-61
The cyanobacterial radiation consists of several lineages of phyletically (morphologically and genetically) related organisms. Several of these organisms show a striking resemblance to fossil counterparts. To investigate the molecular mechanisms responsible for stabilizing or homogenizing cyanobacterial characters, we compared the evolutionary rates and phylogenetic origins of the small-subunit rRNA-encoding DNA (16S rDNA), the conserved gene rbcL (encoding D-ribulose 1,5-bisphosphate carboxylase-oxygenase large subunit), and the less conserved gene rbcX. This survey includes four categories of phyletically related organisms: 16 strains of Microcystis, 6 strains of Tychonema, 10 strains of Planktothrix, and 12 strains of Nostoc. Both rbcL and rbcX can be regarded as neutrally evolving genes, with 95 to 100% and 50 to 80% synonymous nucleotide substitutions, respectively. There is generally low sequence divergence within the Microcystis, Tychonema, and Planktothrix categories both for rbcLX and 16S rDNA. The Nostoc category, on the other hand, consists of three genetically clustered lineages for these loci. The 16S rDNA and rbcLX phylogenies are not congruent for strains within the clustered groups. Furthermore, analysis of the phyletic structure for rbcLX indicates recombinational events between the informative sites within this locus. Thus, our results are best explained by a model involving both intergenic and intragenic recombinations. This evolutionary model explains the DNA sequence clustering for the modern species as a result of sequence homogenization (concerted evolution) caused by exchange of genetic material for neutrally evolving genes. The morphological clustering, on the other hand, is explained by structural and functional stability of these characters. We also suggest that exchange of genetic material for neutrally evolving genes may explain the apparent stability of cyanobacterial morphological characters, perhaps over billions of years.
Важнейшая мысль: описана эволюция прокариот с постоянно идущим горизонтальным переносом генов и из-за этого эволюция идет очень медленно ("стабильность морфологических признаков"). Дискретность морфологического разнообразия объясняется структурной и ФУНКЦИОНАЛЬНОЙ стабильностью признаков. Т.е. дискретностью ниш, а вовсе не эндогамных биосистем – видов или популяций.
* Еще о большой роли горизонтального переноса генов в эволюции эукариот. Особенно – у одноклеточных эукариот много следов такого переноса. Например у анаэробных амитохондриальных эукариот (паразитических) – гены для брожения и т.п. взяты у прокариот. У высших многоклеточных таких фактов меньше. Дополнительные рефераты статей:
Авторы недавнего (и, по мнению специалистов, довольно плохого) обзора про ГМО-еду вспомнили старую страшилку, заключающуюся в том, что фрагменты ДНК из пищи, которую мы употребляем, могут попадать в клетки человека или населяющих его микроорганизмов и влиять на экспрессию генов хозяина или даже встраиваться в геном. Подобных примеров современная наука не знает, но вообще случаи горизонтального переноса генов — попадания в организм фрагментов ДНК не от родителей, а извне, из окружающей среды, ученым известны, и многие из них описаны довольно хорошо. Мы решили разобраться в этом вопросе.
Несмотря на то, что позвоночные животные едят ДНК-содержащую еду миллионы лет, свидетельств тому, что съеденные гены как-то влияют на собственный геном, ученые пока не нашли. Тем не менее, получить ДНК не от родителей, а откуда-то извне возможно — этот феномен называется горизонтальным переносом генов. С началом эры массового секвенирования геномов и биоинформатики стало понятно, что горизонтальный перенос сыграл значительную роль в эволюции как прокариот (бактерий и архей), так и высших эукариот, например, растений.
Тем не менее, заполучить чужую ДНК не так-то просто, а наличие и количество, к примеру, бактериальной ДНК в геноме человека до сих пор остается дискуссионным вопросом.
Без ядра
Для того чтобы в геноме появился новый элемент, необходимо, чтобы новая ДНК попала в клетку и встроилась в хромосому. Логично, что проще всего выполнить эти условия, если организм одноклеточный и у него нет дополнительной ядерной оболочки, защищающей геном. По всей видимости, прокариоты (бактерии и археи) действительно пользуются горизонтальным переносом очень активно — для них это еще и аналог полового процесса, позволяющий внести разнообразие в генетическую информацию, наряду со случайным мутагенезом.
В лаборатории свойство компетентности используют для того, чтобы искусственно доставлять ДНК в бактериальные клетки, а в природе к развитию компетентности и натуральной трансформации способны как минимум несколько десятков видов бактерий, среди которых множество патогенных. Как в случае с опытами Гриффита, внешним источником ДНК могут быть погибшие клетки, кроме того, некоторые бактерии выделяют ее наружу намеренно, с использованием систем секреции.
Помимо трансформации, бактерии способны обмениваться ДНК путем конъюгации. Этот специализированный процесс передачи ДНК между клетками при непосредственном контакте был открыт на кишечной палочке в середине XX века. Для того чтобы передать ДНК, клетки кишечной палочки должны содержать небольшую кольцевую экстрахромосомную молекулу ДНК — плазмиду, которая содержит гены, необходимые для конъюгации, и которая, собственно, и передается.
Конъюгация осуществляется с образованием половых пилей — белковых трубочек, при помощи которых устанавливается физический контакт. Кроме кишечной палочки, процесс был найден и у множества других бактерий. Помимо генов, необходимых для собственного распространения, конъюгативная плазмида может содержать гены других полезных признаков, поддерживаемых отбором, например, устойчивости к антибиотикам.

Молекулярный механизм передачи F-плазмиды путем конъюгации у бактерий
Еще один распространенный механизм передачи генов путем горизонтального переноса — трансдукция, процесс передачи ДНК при помощи вирусов — бактериофагов. При формировании вирусных частиц фаг захватывает часть хозяйской ДНК и при заражении других клеток может передать им чужие гены.
Встроить в геном можно не любую ДНК — в общем случае полученный фрагмент просто разрушится внутриклеточными ферментами — нуклеазами и рестриктазами, которые защищают клетку от вторжения. Обмен генами происходит чаще между близкородственными штаммами, у которых большой процент похожих последовательностей. В таком случае новый фрагмент ДНК может встроиться в геном по механизму гомологичной рекомбинации, для которой необходимо наличие одинаковых или близких по составу нуклеотидных последовательностей.
Странные деревья
Если существование горизонтального переноса было установлено еще до расшифровки последовательностей генома, то масштабы явления стали понятны только с наступлением эры секвенирования. Попытки построить для всех живых организмов универсальное филогенетическое дерево на основании последовательностей геномов привели к ряду филогенетических конфликтов. Нередко какие-то гены обнаруживаются там, где по логике эволюционного наследования, их быть не должно. Отсутствие гена у предков организма часто наводит исследователей на мысль, что он получен путем горизонтального переноса.
Расшифровка множества эукариотических геномов позволила предположить, что горизонтальный перенос сыграл важную роль в эволюции не только бактерий, но и одноклеточных эукариот, в частности, простейших, а также водорослей и высших растений, многих беспозвоночных животных.
У прокариот механизмы доставки новых генов более или менее изучены и понятны, но у эукариот ДНК дополнительно защищена ядерной мембраной и белками-гистонами. Кроме того, если речь идет о многоклеточных организмах с половым размножением, то для того, чтобы закрепиться в поколениях, хромосома с новым элементом должна попасть в половые клетки. Таким образом, на пути горизонтального переноса у более сложных организмов, нежели бактерии, стоит множество барьеров. Поэтому в его наличие стоит верить только при существовании вероятного механизма передачи.
Обмен генетической информацией между ядерным геномом и митохондриальным детектируют и в настоящее время, например, у человека он иногда наблюдается при злокачественном преобразовании клеток. Происходить это событие может, например, во время митоза, когда ядерная оболочка разрушается. Как при этом преодолевается барьер митохондриальной мембраны, не совсем понятно, скорее всего митохондрии просто деградируют, и митохондриальная ДНК выходит в цитоплазму.
Биологам известны и современные примеры эндосимбиоза и эндопаразитизма, сопровождающиеся горизонтальным переносом генов. К примеру, внутриклеточный симбионт членистоногих и некоторых червей-нематод бактерия вольбахия нередко встраивает большие куски своего генома в геном хозяина. Вероятно, это происходит случайно в процессе репарации ДНК, и большой пользы хозяевам вольбахии от этого нет, так как большинство бактериальных генов при этом неактивны или превращаются в псевдогены, то есть необратимо ломаются.

У организмов, размножающихся половым путем, гены родителей перемешиваются в геноме потомства. Это позволяет естественному отбору работать не с целыми геномами, а с отдельными генами, поддерживая удачные варианты и отбраковывая неудачные. Считалось, что у бактерий отбор работает в основном на уровне целых геномов, что намного менее эффективно. Теоретически, горизонтальный обмен генами может отчасти заменить бактериям половое размножение. Однако было неясно, в какой степени это характерно для природных популяций микробов. Изучив две близкородственные популяции морских бактерий, недавно начавших приспосабливаться к разным нишам, американские биологи пришли к выводу, что адаптивная эволюция бактерий идет за счет распространения отдельных генов, а не целых геномов, то есть так же, как у организмов, размножающихся половым путем. Из этого следует, что в ходе адаптации к новым условиям горизонтальный генетический обмен успешно заменяет бактериям половое размножение.
У бесполых организмов, практикующих клональное размножение (например, путем почкования или деления надвое), отбору приходится работать с целыми геномами. При этом судьбы всех полезных и вредных мутаций, возникающих в геноме, оказываются неразрывно связаны друг с другом. Это накладывает серьезные ограничения на адаптивные возможности организмов.
Биологи из Массачусетского технологического института (MIT) попытались прояснить вопрос о микроэволюционной роли ГГО на примере двух популяций морских планктонных бактерий Vibrio cyclitrophicus. Ранее авторы показали, что эти две популяции, обозначаемые буквами L и S, разделились недавно и в настоящее время находятся в процессе адаптации к разным экологическим нишам. Это видно по их приуроченности, соответственно, к крупным (L) и мелким (S) частицам, отфильтрованным из морской воды, и по результатам генетического анализа (см.: Hunt et al., 2008. Resource Partitioning and Sympatric Differentiation Among Closely Related Bacterioplankton, PDF, 529 Кб). По-видимому, эти две популяции приспосабливаются к жизни на разных представителях зоо- или фитопланктона.
Авторы решили выяснить, какой из двух процессов преобладает на начальных этапах экологической дифференциации — отбор отдельных генов с удачными мутациями, которые распространяются за счет ГГО, или клональный отбор, поддерживающий или отбраковывающий целые геномы. Различить эти две ситуации можно, сравнив внутри- и межпопуляционный генетический полиморфизм (вариабельность) изучаемых популяций.
Если преобладает отбор на уровне генов, две популяции должны четко отличаться друг от друга по небольшому числу генов — тех, от которых зависят экологические свойства популяций. При этом различающиеся участки генома должны иметь пониженный уровень внутрипопуляционного полиморфизма хотя бы в одной из двух популяций. Ведь на каждый такой участок действовал отбор, который поддерживал какой-то один вариант этого участка, вытесняя из генофонда другие его варианты. Напротив, те участки генома, которые у двух популяций сходны, должны быть более полиморфными, причем наборы генетических вариантов (аллелей) могут быть одинаковыми в двух популяциях (потому что отбор, связанный с приспособлением к новой нише, на них не действовал, и они могли сохранить исходный полиморфизм, накопленный предковой популяцией).
Если же в процессе расхождения популяций преобладал клональный отбор, то и уровень полиморфизма, и генетические различия между популяциями должны быть распределены по геному более равномерно.
Авторы отсеквенировали полные геномы тринадцати бактерий из популяции L и семи особей из популяции S. Анализ прочтенных геномов подтвердил первую версию: в недавней эволюции двух бактериальных популяций явно преобладал отбор на уровне отдельных генов, что было бы невозможно без интенсивного ГГО между близкородственными микробами.
Все прочие снипы (полиморфные нуклеотидные позиции), в числе 28 744 штук, равномерно рассеяны по геному и демонстрируют сходный характер полиморфизма в обеих популяциях. Например, в какой-то позиции нуклеотид Ц может стоять у шести, а нуклеотид Г — у семи бактерий из популяции L, тогда как в популяции S нуклеотид Ц обнаружен у трех бактерий, а Г — у четырех.
Обнаруженная картина распределения нуклеотидных различий полностью соответствует предположению о том, что в ходе приспособления к различающимся условиям отбор в сочетании с ГГО (и гомологичной рекомбинацией) способствовал распространению отдельных участков ДНК с удачными мутациями, а не целых геномов, причем в остальных участках генома сохранялся исходный полиморфизм, имевшийся у предковой популяции.
Еще одним подтверждением интенсивного ГГО между близкородственными микробами является то обстоятельство, что филогенетические (эволюционные) деревья, построенные для 20 изученных бактерий, оказываются разными в зависимости от того, какие фрагменты генома используются для их построения. Анализируя эти деревья, а также используя другие методы сравнительной геномики, авторы обнаружили свидетельства многочисленных недавних переносов генетического материала внутри каждой из двух популяций. В частности, удалось показать быстрое распространение целой хромосомы в популяции S. У Vibrio cyclitrophicus геном состоит из двух хромосом, обозначаемых цифрами I и II. Один из вариантов хромосомы II недавно распространился под действием отбора в популяции S (он обнаружен у 5 из 7 бактерий). Хромосома II распространялась независимо от хромосомы I, а это значит, что бактерии запросто могут обмениваться целыми хромосомами, а не только их фрагментами.
Еще один важный вывод состоит в том, что генетический обмен между популяциями в последнее время происходит редко по сравнению с обменом внутри популяций. В прошлом, когда популяции еще не окончательно разошлись по экологическим нишам, межпопуляционный обмен происходил чаще.


Модель экологической дифференциации двух популяций морской бактерии Vibrio cyclitrophicus. Тонкие серые и черные стрелки символизируют внутри- и межпопуляционный ГГО. Толстые цветные стрелки показывают появление (в результате мутации или ГГО) адаптивных аллелей (генетических вариантов), облегчающих жизнь в двух разных местообитаниях (зеленом и красном). Изображение из обсуждаемой статьи в Science
Полученные результаты говорят о необходимости пересмотра некоторых устоявшихся представлений об эволюции прокариот.
Источник: B. Jesse Shapiro, Jonathan Friedman, Otto X. Cordero, Sarah P. Preheim, Sonia C. Timberlake, Gitta Szabó, Martin F. Polz, Eric J. Alm. Population Genomics of Early Events in the Ecological Differentiation of Bacteria // Science. 2012. V. 336. Pp. 48–51.

С момента обнаружения такого феномена, как горизонтальный перенос генов, а именно не от родителей к потомкам, весь живой мир на нашей планете представляется единой информационной системой. И в этой системе становится возможным заимствование удачного эволюционного изобретения одного вида другим. Что такое вертикальный и горизонтальный перенос генов, каковы механизмы этого процесса и примеры в органическом мире – обо всем этом статья.
Гены соседа
Каждому известно, что свои гены мы получаем от родителей. А они — от своих родителей. Это и есть вертикальный перенос. И если вдруг произойдет мутация, которая окажется полезной для выживания или приспособления, и закрепится в геноме популяции, то вид приобретет преимущества в борьбе за существование.
При этом у человека свои гены, у тли — свои, а у акул — свои. Попасть им между видами практически нет возможности. Но иногда такое случается – это и есть перенос генов горизонтальный.
Именно этим и занимается современная генная инженерия. Генномодифицированные организмы являются результатом такого переноса генов (например, светящаяся тихоходка на фото выше). Но в природе это явление существует давно.
Суть вопроса
Вертикальный перенос генов — явление передачи наследственного материала от родительских форм к дочерним организмам.
Горизонтальный перенос генов — это существующая в природе ситуация передачи генов от одного взрослого организма к другому. При этом два организма объективно существуют, а иногда — относятся к разным биологическим видам.
Пример горизонтального переноса генов у бактерий – перенос генов устойчивости (резистентности) от одного штамма бактерий к другому.
Необходимые условия
Для понимания данного явления необходимо знать условия, при которых такой перенос возможен в принципе, а именно:
Названные условия вполне могут выполнять ретровирусы и другие транспозооны (элементы ДНК). И именно такие способы горизонтального переноса генов сегодня взяла на вооружение генная инженерия.
Хотя сегодня механизмы такого переноса генов только изучаются, кроме вирусов, такой перенос может происходить и при помощи свободных участков дезоксирибонуклеиновых кислот (транспозоонов), которые попадают в организм посредством простого заноса или с паразитическими организмами. Последние могут менять не только генетический аппарат хозяина, но и его экологическое место в системе биоценоза.

История вопроса
Именно перенос генов резистентности к антибиотикам между разными штаммами бактерий был впервые описан в Японии в 1959 году.
Уже к середине 90-х годов прошлого столетия молекулярными биологами было доказано, что горизонтальный перенос генов у прокариот и эукариот был вовлечен в эволюционное развитие жизни на нашей планете.
В 2010 году было опубликовано исследование профессора Седрика Фешотта, в котором был представлен анализ генома опоссума и обезьян саймири. Их укусил один вид клопа. В геномах млекопитающих был обнаружен транспозоон, идентичность которого с насекомым составляет 98%. К сведению, кусают эти клопы не только обезьянок и опоссумов.
С этого момента гипотеза горизонтального переноса генов между разными доменами организмов стала новой парадигмой биологии.

Разноцветные букашки
И если горизонтальный перенос генов у бактерий последние 30 лет не вызывал сомнений у биологов, то его возможность у многоклеточных вызывала множество вопросов. Именно тогда внимание биологов привлекла обычная тля, у которой встречаются особи с зеленым и красным цветом тельца.
Анализ пигментов, которые придают окраску красным особям, выявил наличие каротиноидов – растительных пигментов. Откуда же у тли взялись гены, которые присущи исключительно растительным организмам? Сегодня секвенировать геном букашки – дело довольно простое для исследователей. Именно так было обнаружено, что гены тли, ответственные за синтез красного пигмента, полностью идентичны таковому у некоторых грибов, которые паразитируют в организме тли, не причиняя ей видимого вреда.
Вероятнее всего, на заре эволюции тли (около 80 миллионов лет назад) и произошел сбой в генетической машине и гены грибов оказались встроенными в геном букашки.
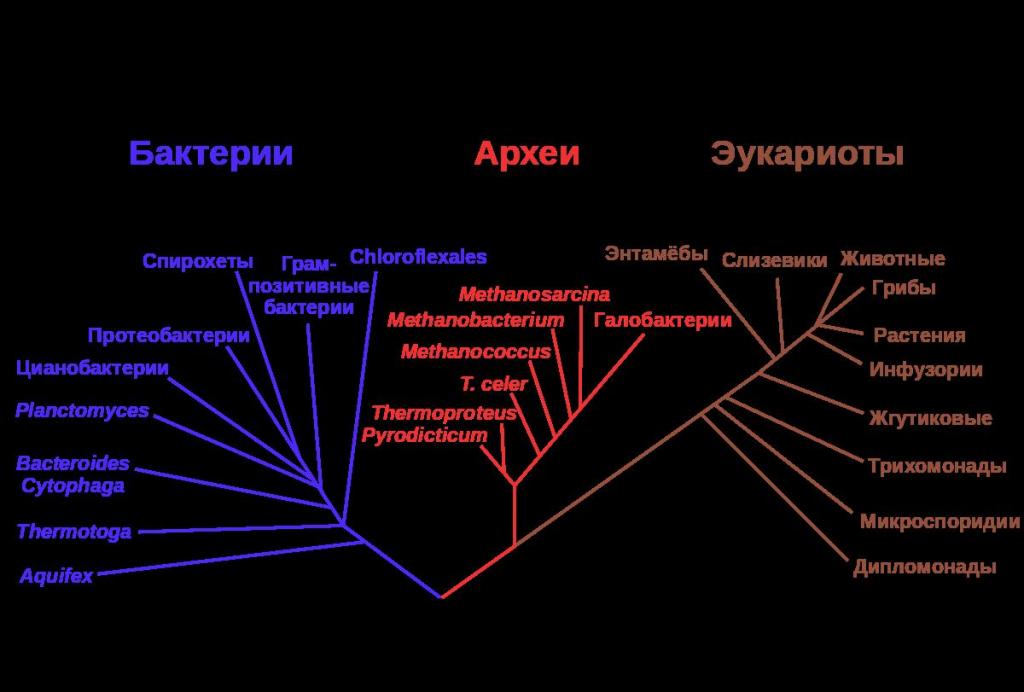
Эволюция и биоразнообразие
Вся филогенетическая систематика органического мира базируется на Дарвиновском понятии дивегренции. Суть его заключается в следующем: как только между популяциями вида возникает репродуктивная изоляция, можно говорить о процессе видообразования. И уже два вида продолжают эволюционировать на основе естественного отбора и случайных мутаций.
Открытие горизонтального переноса генов между видами и более крупными таксонами только доказало, что за столь короткий по космическим меркам срок (4 миллиарда лет) живая материя на нашей планете могла пройти путь от одноклеточных форм до высокоорганизованных многоклеточных.
Таким образом, вся биота планеты становится единой лабораторией по созданию новых наследственных признаков, и именно горизонтальное движение генов смогло и продолжает существенно ускорять эволюционный процесс.
Возьмем взаймы немного генов
Результаты исследований подтвердили наличие в геномах 145 участков, которые стали результатом горизонтального переноса генов у эукариот.
Часть этих генов участвует в метаболизме белков и липидов, другая — в иммунных реакциях. Главное, удалось выявить вероятных доноров этих генов. Ими оказались протисты (простейшие эукариоты), бактерии (прокариоты) и грибы.
А что в отношении нас
Уже достоверно известно, что путем горизонтального переноса генов у человека появились гены, ответственные за группы крови АВ0.
Большинство фактов такого переноса генов у приматов имеет очень древнее происхождение и относится ко времени существования общего предка с другими хордовыми животными.
По данным последних исследований, за формирование плаценты у человека отвечает в том числе и ген вируса, который был захвачен где-то на заре формирования плацентарных животных.

Эра мутантов
Во-первых, механизм горизонтального переноса существует в природе столько, сколько существует сама жизнь. И на примере тли отлично видно, что такой перенос генов был направлен именно на повышение приспособленности организмов к условиям окружающей среды (красненьких хуже видно на определенных частях растений). И генные инженеры в этом смысле не придумали ничего нового. Томаты с генами арктических рыб имеют повышенную холодоустойчивость, что позволяет выращивать их в северных регионах.
Во-вторых, несмотря на возможность генетического переноса, мы пока не наблюдаем унификацию (однообразность) генома всех живых организмов на планете. Стабильность биологической системы, которой является клетка и организм, достаточно высокая и ограничивает неэффективный перенос генов. Но в то же время именно такой перенос является инструментом биологической эволюции, который ведет к биоразнообразию. Так что нескоро медведи станут похожи на коршунов, а собаки — на хамелеонов.
Читайте также: