
Гетерохроматин это в биологии кратко
Обновлено: 05.07.2024
Гетерохроматин это плотно упакованная форма ДНК или конденсированная ДНК, который существует в нескольких вариантах. Эти разновидности лежат в континууме между двумя крайностями конститутивный гетерохроматин и факультативный гетерохроматин. Оба играют роль в экспрессия генов. Поскольку он плотно упакован, считалось, что он недоступен для полимераз и поэтому не транскрибируется, однако, согласно Volpe et al. (2002), [1] и многие другие документы с тех пор, [2] большая часть этой ДНК фактически транскрибируется, но она постоянно перевернулся через РНК-индуцированное подавление транскрипции (РИТС). Недавние исследования с электронная микроскопия и OsO4 окрашивание показывает, что плотная упаковка не связана с хроматином. [3]
Конститутивный гетерохроматин может влиять на гены, расположенные рядом с ним (например, позиционно-эффектное разнообразие). Это обычно повторяющийся и образует структурные функции, такие как центромеры или теломеры, помимо действия в качестве аттрактора для других сигналов экспрессии или репрессии генов.
Факультативный гетерохроматин является результатом генов, которые замолчать через такой механизм, как деацетилирование гистонов или Piwi-взаимодействующая РНК (пиРНК) через РНКи. Он не повторяется и имеет компактную структуру конститутивного гетерохроматина. Однако при определенных онтогенетических или средовых сигнальных сигналах он может потерять свою конденсированную структуру и стать транскрипционно активным. [4]
Гетерохроматин был связан с ди- и три-метилированием H3K9 в определенных частях генома. [5] H3K9me3-Связанный метилтрансферазы по-видимому, играют ключевую роль в модификации гетерохроматина во время фиксации клонов в начале органогенез и в поддержании верности родословной. [6]
Содержание
Структура
Гетерохроматин в основном состоит из генетически неактивных спутниковые последовательности, [11] и многие гены в разной степени репрессированы, хотя некоторые вообще не могут экспрессироваться в эухроматине. [12] И то и другое центромеры и теломеры гетерохроматичны, как и Тело Барра второй, инактивированной Х-хромосома в женском.
Функция
Гетерохроматин связан с несколькими функциями, от регуляции генов до защиты целостности хромосом; [13] некоторые из этих ролей можно отнести к плотной упаковке ДНК, которая делает ее менее доступной для белковых факторов, которые обычно связывают ДНК или связанные с ней факторы. Например, голые концы двухцепочечной ДНК обычно интерпретируются клеткой как поврежденная или вирусная ДНК, вызывая клеточный цикл арест Ремонт ДНК или уничтожение фрагмента, например эндонуклеазы в бактериях.
Некоторые участки хроматина очень плотно упакованы волокнами, состояние которых сопоставимо с состоянием хромосомы на митоз. Гетерохроматин обычно наследуется клонально; когда клетка делится, две дочерние клетки обычно содержат гетерохроматин в одних и тех же областях ДНК, что приводит к эпигенетическая наследственность. Вариации заставляют гетерохроматин вторгаться в соседние гены или отступать от генов на крайних участках доменов. Транскрибируемый материал может быть подавлен путем размещения (в СНГ) в этих граничных областях. Это приводит к повышению уровней экспрессии, которые варьируются от клетки к клетке, [14] что может быть продемонстрировано позиционно-эффектное разнообразие. [15] Изолятор последовательности могут действовать как барьер в редких случаях, когда конститутивный гетерохроматин и высокоактивные гены сопоставляются (например, инсулятор 5'HS4 перед локусом β-глобина курицы, [16] и локусы в двух Сахаромицеты виды [17] [18] ).
Конститутивный гетерохроматин
Все клетки данного вида упаковывают одни и те же участки ДНК в конститутивный гетерохроматин, и, следовательно, во всех клетках любые гены, содержащиеся в конститутивном гетерохроматине, будут плохо выразил. Например, все хромосомы человека 1, 9, 16, а Y-хромосома содержат большие области конститутивного гетерохроматина. У большинства организмов конститутивный гетерохроматин находится вокруг центромеры хромосомы и около теломер.
Факультативный гетерохроматин
Области ДНК, упакованные в факультативный гетерохроматин, не будут согласовываться между типами клеток внутри вида, и, таким образом, последовательность в одной клетке, которая упакована в факультативный гетерохроматин (а гены внутри нее экспрессируются слабо), может быть упакована в эухроматин в другой клетке. (и гены внутри больше не заглушаются). Однако образование факультативного гетерохроматина регулируется и часто связано с морфогенез или дифференциация. Примером факультативного гетерохроматина является Инактивация Х-хромосомы у самок млекопитающих: один Х хромосома упакован как факультативный гетерохроматин и замалчивается, тогда как другая Х-хромосома упакована как эухроматин и экспрессируется.
Среди молекулярных компонентов, которые, по-видимому, регулируют распространение гетерохроматина, находятся Белки группы поликомб и некодирующие гены, такие как Xist. Механизм такого распространения до сих пор вызывает споры. [19] Поликомб репрессивные комплексы PRC1 и PRC2 регулировать хроматин уплотнение и экспрессия генов и играют фундаментальную роль в процессах развития. PRC-опосредованный эпигенетический аберрации связаны с нестабильность генома и злокачественные новообразования и играют роль в Повреждение ДНК ответ, Ремонт ДНК и в верности репликация. [20]
Гетерохроматин дрожжей
Saccharomyces cerevisiae, или зародышевые дрожжи, это модель эукариот и его гетерохроматин был тщательно определен. Хотя большую часть его генома можно охарактеризовать как эухроматин, С. cerevisiae имеет участки ДНК, которые очень плохо транскрибируются. Эти локусы представляют собой так называемые локусы типа молчащего спаривания (HML и HMR), рДНК (кодирующую рибосомную РНК) и субтеломерные области.Schizosaccharomyces pombe) использует другой механизм образования гетерохроматина на его центромерах. Подавление гена в этом месте зависит от компонентов РНКи путь. Полагают, что двухцепочечная РНК приводит к подавлению этого региона посредством ряда этапов.
В делящихся дрожжах Schizosaccharomyces pombeдва комплекса РНКи, комплекс RITS и комплекс РНК-направленной РНК-полимеразы (RDRC), являются частью механизма РНКи, участвующего в инициации, размножении и поддержании сборки гетерохроматина. Эти два комплекса локализуются в миРНК-зависимо от хромосом, в месте сборки гетерохроматина. РНК-полимераза II синтезирует транскрипт, который служит платформой для набора RITS, RDRC и, возможно, других комплексов, необходимых для сборки гетерохроматина. [21] [22] И РНКи, и процесс деградации РНК, зависимый от экзосом, вносят вклад в молчание гетерохроматических генов. Эти механизмы Schizosaccharomyces pombe может встречаться и у других эукариот. [23] Большая структура РНК, называемая RevCen также участвует в производстве siRNAs, которые опосредуют образование гетерохроматина у некоторых делящихся дрожжей. [24]
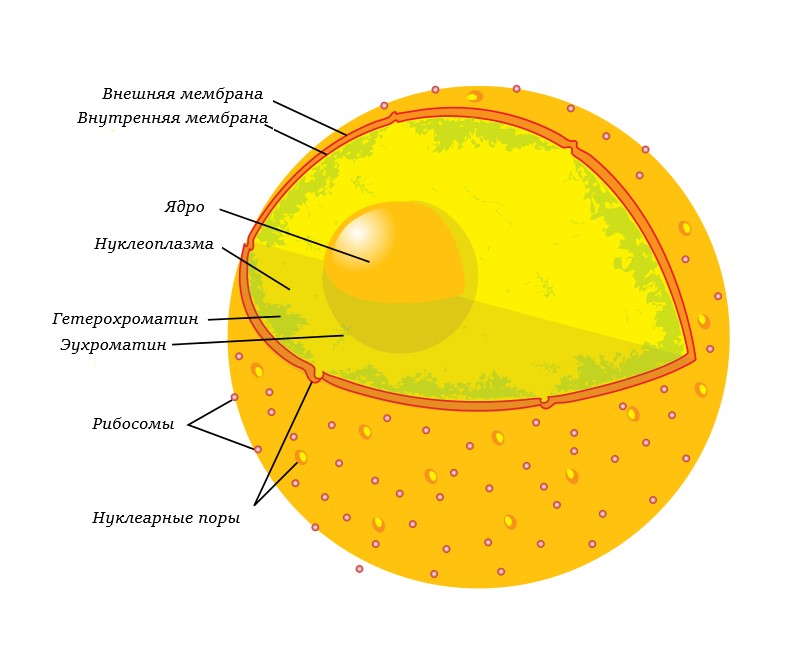
Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоидау прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, —надмолекулярных структур, участвующих в упаковке хромосом.
1.Эухроматин – локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компактный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка.
2.Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. Гетерохроматин - плотно спирализованная часть хроматина, соответствует конденсированным, плотно скрученным сегментам хромосом, что делает их недоступными для транскрипции. Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул.
Метафазные хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. На стадии метафазы сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначается буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.
В зависимости от расположения центромеры различают три типа строения хромосом:
1. Акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
2. Субметацентрические хромосомы с плечами неравной длины;
3. Метацентрические хромосомы, у которых центромерарасположена посередине или почти посередине.
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомальные РНК. Небольшие хромосомные сегменты, отделяемые от основного тела хромосомы вторичными перетяжками, называются спутниками.
Хроматин — это вещество хромосом — комплекс ДНК, РНК и белков. Хроматин находится внутри ядра клеток эукариот и входит в состав нуклеоидау прокариот. Именно в составе хроматина происходит реализация генетической информации, а также репликация и репарация ДНК. Основную массу хроматина составляют белки гистоны. Гистоны являются компонентом нуклеосом, —надмолекулярных структур, участвующих в упаковке хромосом.
1.Эухроматин – локализующийся ближе к центру ядра, более светлый, более деспирилизованный, менее компактный, более активен в функциональном отношении. Эухроматин - неконденсированный хроматин, с которого происходит синтез белка.
2.Гетерохроматин - конденсированный хроматин, с которого белок не синтезируется. Гетерохроматин - плотно спирализованная часть хроматина, соответствует конденсированным, плотно скрученным сегментам хромосом, что делает их недоступными для транскрипции. Он интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид тёмных пятен, гранул.
Метафазные хромосомы состоят из двух продольных копий, которые называются сестринскими хроматидами и которые образуются при репликации. На стадии метафазы сестринские хроматиды соединены в районе первичной перетяжки, называемой центромерой. Центромера отвечает за расхождение сестринских хроматид в дочерние клетки при делении. На центромере происходит сборка кинетохора — сложной белковой структуры, определяющей прикрепление хромосомы к микротрубочкам веретена деления — движителям хромосомы в митозе. Центромера делит хромосомы на две части, называемые плечами. У большинства видов короткое плечо хромосомы обозначается буквой p, длинное плечо — буквой q. Длина хромосомы и положение центромеры являются основными морфологическими признаками метафазных хромосом.

В зависимости от расположения центромеры различают три типа строения хромосом:
1. Акроцентрические хромосомы, у которых центромера находится практически на конце, и второе плечо настолько мало, что его может быть не видно на цитологических препаратах;
2. Субметацентрические хромосомы с плечами неравной длины;
3. Метацентрические хромосомы, у которых центромерарасположена посередине или почти посередине.
Дополнительным морфологическим признаком некоторых хромосом является так называемая вторичная перетяжка, которая внешне отличается от первичной отсутствием заметного угла между сегментами хромосомы. Вторичные перетяжки бывают короткими и длинными и локализуются в разных точках по длине хромосомы. Во вторичных перетяжках находятся, как правило, ядрышковые организаторы, содержащие многократные повторы генов, кодирующих рибосомальные РНК. Небольшие хромосомные сегменты, отделяемые от основного тела хромосомы вторичными перетяжками, называются спутниками.

Прежде всего… Что такое хроматин?
Хроматин – это нуклеопротеиновый комплекс. А именно хроматин - это полимер, включающий в себя в специальные хромосомные белки, нуклеосомы и ДНК. Белки могут составлять до 65% от массы хромосомы. Хроматин представляет собой динамическую молекулу и может принимать огромное количество конфигураций.

Белки хроматина составляют весомую часть от его массы и подразделяются на две группы:
- Гистоновые белки - содержат основные аминокислоты в своем составе (например, аргинин и лизин). Расположение гистонов хаотично в виде блоков по всей протяженности молекулы ДНК. (примерно 1/5 часть от общего количества гистонов) - представляют собой ядерный белковый матрикс, образующий в интерфазном ядре структурную сеть. Именно она является основой, определяющие морфологию и метаболизм ядра.
В настоящее время в цитогенетике хроматин подразделяется на две разновидности: гетерохроматин и эухроматин. Разделение хроматина на два вида произошло из-за способности каждого вида окрашиваться специфичными красителями. Это эффективный способ визуализации ДНК, используемый цитологами.
Гетерохроматин
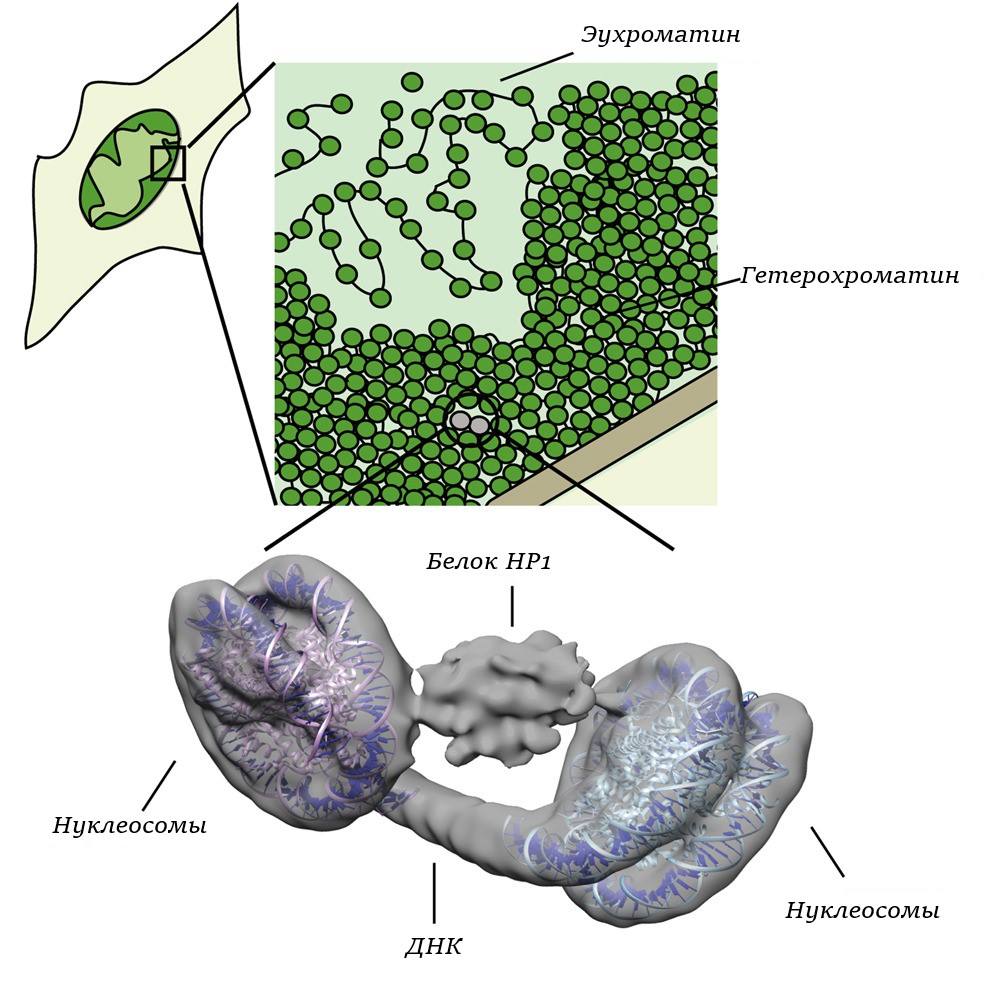
Гетерохроматин – это частично конденсированные в интерфазе участки хромосомы. Функционально гетерохроматин не представляет никакой ценности, так как не активен, именно в отношении транскрипции. Но его способность хорошо окрашиваться широко применяется в гистологических исследованиях.
Строение гетерохроматина
Гетерохроматин имеет несложное строение (см. рисунок).
Гетерохроматин - это не только отдельные части, но и целые хромосомы, которые на протяжении всего клеточного цикла остаются в конденсированном состоянии. Именно они в S-фазе и подвергаются репликации. Ученые считают, что гетерохроматиновые участки не несут в себе гены, которые кодируют белок, либо количество таких генов очень незначительно. Вместо таких генов нуклеотидные последовательности гетерохроматина в большинстве своих состоят из простых повторов.
Виды гетерохроматина
Гетерохроматин бывает двух видов: факультативный и структурный.
- Факультативный гетерохроматин - это хроматин, который образуется в процессе образования спирали одной из двух хромосом одного вида, он не всегда гетерохроматичен, а временами. В нем содержатся гены с наследственной информацией. Она считывается при переходе его в эухроматическое состояние. Конденсированное состояние для факультативного гетерохроматина – явление временное. В этом его главное отличие от структурного. Примером факультативного гетерохроматина можно назвать тело хроматина, определяющего женскую половую принадлежность. Так как такая структура состоит из двух гомологичных Х-хромосом соматических клеток, одна из них может как раз образовать факультативный гетерохроматин.
- Структурный гетерохроматин – это структура, образованная высокоспирализованным состоянием. Оно сохраняется на протяжении всего цикла. Как сказано выше, конденсированное состояние для структурного гетерохроматина – явление постоянное, в отличие от факультативного. Структурный гетерохроматин называют еще конститутивным, он хорошо выявляется С-окраской. Находится подальше от ядра и занимает прицентромерные области, но иногда локализируется и в других областях хромосомы. Часто в процессе интерфазы может произойти агрегация различных участков структурного гетерохроматина, в результате чего образуются хромоцентры. В подобном виде гетерохроматина отсутствует свойство транскрибации, то есть структурных генов нет. Роль такого участка хромасомы не совсем ясна до нынешнего времени, поэтому деятели науки склоняются к лишь опорной функции.
Эухроматин
Эухроматин – это участки хромосом, которые деконденсированы в интерфазе. Такой локус представляет собой разрыхленную, но вместе с тем небольшую компактную структуру.
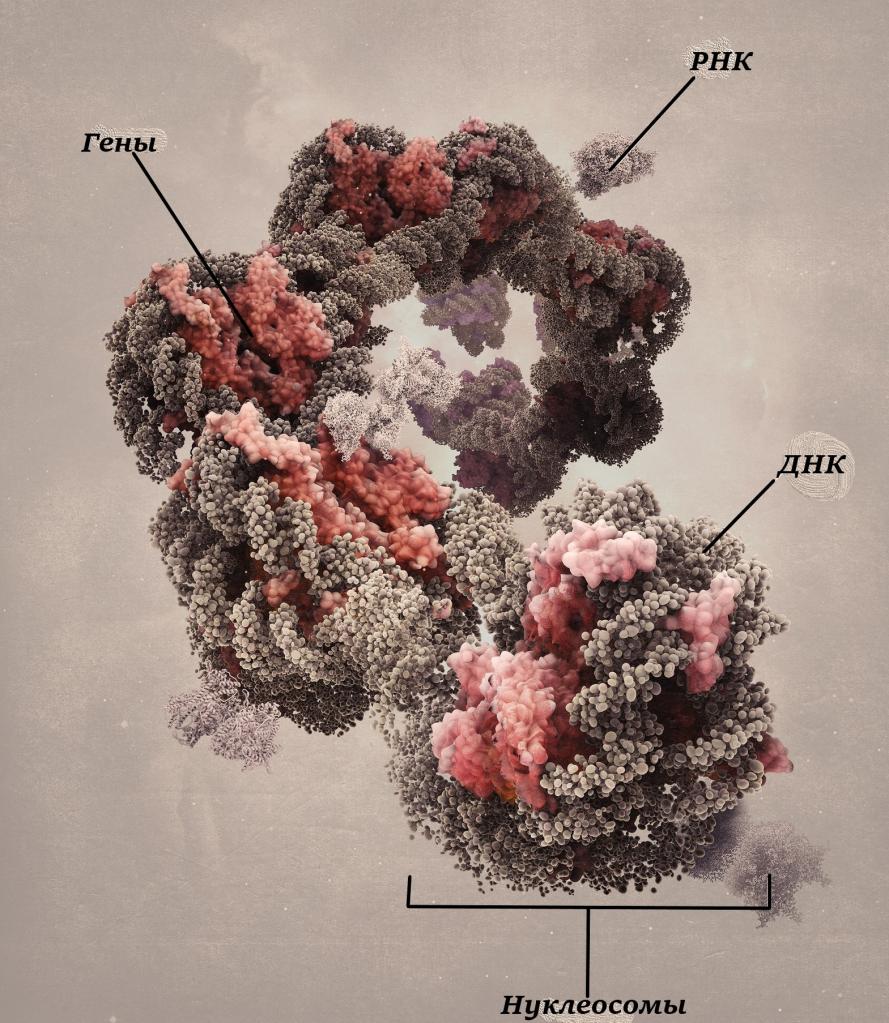
Функциональные особенности эухроматина
Подобный вид хроматина рабочий и функционально активен. Не обладает свойством окрашивания и не определяется при гистологических исследованиях. В фазе митоза эухроматин практически весь конденсируется и становится составляющей частью хромосомы. Синтетические функции в этот период хромосомы не выполняют. Поэтому клеточные хромосомы могут находиться в двух функционально-структурных состояниях:
- Активное или рабочее состояние. В это время хромосомы почти полностью или полностью деконденсированы. Они участвуют в процессе транскрипции и редупликации. Все перечисленные процессы происходят непосредственно в ядре клетки.
- Неактивное состояние метаболического покоя (нерабочее). В этом состоянии хромосомы по максимуму конденсированы и служат транспортом для переноса генетического материала в дочерние клетки. В этом состоянии генетический материал еще и распределяется.
В завершающей фазе митоза происходит деспирализация и образуются слабоокрашенные структуры в виде нитей, содержащие транскрибируемые гены.
В структуре каждой хромосомы свой, уникальный, вариант расположения хроматина: эухроматина и гетерохроматина. Эта особенность клеток позволяет цитогенетикам идентифицировать отдельные хромосомы.
Гетерохроматин является формой хроматина, который плотно упакован – в отличие от эухроматин, который легко упакован – и находится в ядре эукариотических клеток. В то время как эухроматин позволяет реплицировать и транскрибировать ДНК, гетерохроматин находится в такой конденсированной структуре, что не позволяет ДНК- и РНК-полимеразам получать доступ к ДНК, что предотвращает репликацию ДНК и транскрипция, Существует два основных типа гетерохроматина: конструктивный гетерохроматин и факультативный гетерохроматин. Гетерохроматин составляет менее 10% человеческого хроматина, а эухроматин составляет большую его часть – более 90%.
Структура гетерохроматина
Прежде чем мы перейдем к структуре гетерохроматина, давайте посмотрим, как ДНК упаковывается в эукариотических клетках.
ДНК у эукариот собирается в хроматин, который представляет собой комплексы, состоящие из ДНК и белков. Белки, которые образуют хроматин, называются гистонами, и они расположены таким образом, что позволяет ДНК оборачиваться вокруг них. Более конкретно, ДНК (около 200 пар оснований) сворачивается вокруг наборов из восьми гистонов (октамеров), включающих две копии каждого из следующего: H2A, H2B, H3 и H4. Эти единицы, сделанные из гистонов и ДНК, свернутой вокруг них, называются нуклеосомами. Нуклеосомы, в свою очередь, связаны друг с другом через цепочки ДНК, также известные как линкерная ДНК. Другими словами, хроматин – это совокупность нуклеосом (ДНК и гистонов), связанных самой ДНК.
Наиболее свободно упакованную форму хроматина называют эухроматином, также известным как бусы на струне из-за сходства между этой структурой и бусами (нуклеосомами), скрепленными цепочкой (ДНК). Гетерохроматин является более плотно конденсированной версией эухроматина и также известен как волокно с длиной волны 30 нм, потому что диаметр этого спирально спирального гетерохроматина составляет 30 нм. Фактически, в то время как G-полосатость показывает очень слабо окрашенный эухроматин из-за его рыхлой формы, гетерохроматин легко увидеть, потому что он плотно окрашен из-за его более плотной упаковки. Гетерохроматин может также в дальнейшем конденсироваться в активные хромосомы и даже дальше в метафазы хромосомы.
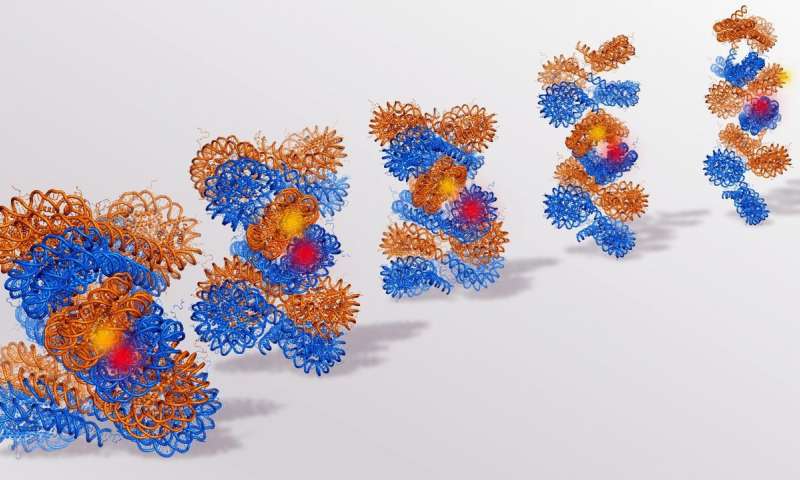
На следующем рисунке показаны различные структурные единицы упаковки ДНК в эукариотических клетках:

Слева направо, двухцепочечная спиральная ДНК (первая иллюстрация) обвивается вокруг гистонов, образуя нуклеосомы (вторая иллюстрация), которые составляют структуру эухроматина или бусы на струне (третья иллюстрация). Эухроматин далее конденсируется в гетерохроматин или 30-нм волокна (четвертая и пятая иллюстрации). Последние четыре иллюстрации изображают более плотно конденсированные ДНК в виде активных и метафазных хромосом.
Глядя на рисунок выше, мы также можем понять, почему ДНК находится в конформации гетерохроматина, когда она не активно реплицируется или транскрибируется: ДНК не подвергается воздействию и, следовательно, регуляторные белки и полимеразы не могут получить к ней доступ. Обратите внимание на разницу между эухроматином (третья иллюстрация) и гетерохроматином (четвертый и пятый): в то время как ДНК-линкер в эухроматической конформации подвергается воздействию полимеразы и других белков и доступен для репликации и транскрипции, ДНК в гетерохроматической конформации плотно скручена. вокруг нуклеосом и не дает доступа к элементам транскрипции.
Два типа гетерохроматина: конститутивный и факультативный
Структура гетерохроматина может быть описана более подробно с учетом его нескольких типов. Двумя основными типами являются конститутивный гетерохроматин и факультативный гетерохроматин. Эти два типа можно различить в зависимости от их особенностей. Предполагается, что существуют также другие типы гетерохроматина и что эти другие типы имеют смешанные признаки конститутивного и факультативного гетерохроматина.
Конститутивный гетерохроматин является стабильной формой гетерохроматина, то есть он не разрыхляется до образования эухроматина и содержит повторяющиеся последовательности ДНК, называемые сателлитной ДНК. Он может быть обнаружен в центромерах и теломерах и обычно участвует в структурных функциях.
Факультативный гетерохроматин, с другой стороны, является обратимым, то есть его структура может изменяться в зависимости от клеточный цикл и характеризуется другим видом повторяющихся последовательностей ДНК, известных как последовательности LINE. Пример факультативного гетерохроматина, который изменяет свою структурную конформацию с клетка цикл инактивированный X-хромосома (Барр тело ) женщин.
Клеточный цикл и экспрессия генов
Не удивительно, что способ, которым упакована ДНК, связан с клеточным циклом. Когда ДНК должна быть скопирована (реплицирована), а белки должны быть синтезированы (транскрипция, а затем перевод ), ДНК находится в форме эухроматина. Когда гены не нуждаются в репликации и транскрибировании, ДНК находится в форме гетерохроматина. Кроме того, когда ДНК находится в активной форме хромосомы, клетка находится в интерфаза стадия клеточного цикла, и когда она находится в форме метафазной хромосомы, клетка находится в делящемся, то есть в митоз или мейоз сцена.
В соответствии с этим было высказано предположение, что регулирование способа упаковки ДНК является способом регулирования ген выражение. Следовательно, гены домашнего хозяйства, которые поддерживают функции и выживание клетки, всегда находятся в форме эухроматина, тогда как гены, которые не нуждаются в экспрессии, находятся в форме гетерохроматина. Средства, с помощью которых это достигается, – это модификация хвоста гистонов, части гистонов, которые могут быть ацетилированы или метилированы. Модификация хвоста гистона приводит к изменениям в упаковке ДНК. Например, гипоацетилирование на хвосте гистонов связано с гетерохроматической конформацией, при которой ДНК не подвергается воздействию и, следовательно, генная транскрипция предотвращается.
викторина
1. Что показывает наличие гетерохроматина?A. Эти клетки транскрипционно активны.B. Это клетки делятся.C. Эта транскрипция гена не происходит.D. Эта ДНК подвергается воздействию полимераз и других регуляторных белков.
Ответ на вопрос № 1
С верно. Гетерохроматин плотно упакован и не позволяет полимеразам и другим регуляторным белкам получать доступ к ДНК, тем самым предотвращая транскрипцию генов.
2. Каковы два основных различия между конститутивным и факультативным гетерохроматином?A. Конститутивный гетерохроматин является обратимым и имеет последовательности LINE, тогда как факультативный гетерохроматин стабилен и имеет сателлитную ДНК.B. Конститутивный гетерохроматин стабилен и имеет последовательности LINE, тогда как факультативный гетерохроматин обратим и имеет сателлитную ДНК.C. Конститутивный гетерохроматин обратим и имеет сателлитную ДНК, тогда как факультативный гетерохроматин стабилен и имеет последовательности LINE.D. Конститутивный гетерохроматин стабилен и имеет сателлитную ДНК, тогда как факультативный гетерохроматин обратим и имеет последовательности LINE.
Ответ на вопрос № 2
D верно. Оба очень похожи, но два основных различия, которые их отличают, состоят в том, что конститутивный гетерохроматин стабилен и имеет сателлитную ДНК, в то время как факультативный гетерохроматин обратим и имеет последовательности LINE.
3. Как называется гетерохроматин?A. Бусины-на-строкиB. 30-нм волокноC. Активная хромосомаD. Метафазная хромосома
Ответ на вопрос № 3
В верно. Гетерохроматин также известен как волокно 30 нм, потому что диаметр спиральной структуры, которую он образует, составляет 30 нм.
Читайте также: