
Генетика популяций кратко и понятно
Обновлено: 05.07.2024
Популяция — это совокупность особей одного вида, длительное время обитающих на определенной территории, свободно скрещивающихся друг с другом, имеющих общее происхождение, определенную генетическую структуру и в той или иной степени изолированных от других таких совокупностей особей данного вида. Популяция не только единица вида, форма его существования, но и единица эволюции. В основе микроэволюционных процессов, завершающихся видообразованием, лежат генетические преобразования в популяциях.
Изучением генетической структуры и динамики популяций занимается особый раздел генетики — популяционная генетика.
С генетической точки зрения, популяция является открытой системой, а вид — закрытой. В общей форме процесс видообразования сводится к преобразованию генетически открытой системы в генетически закрытую.
Каждая популяция имеет определенный генофонд и генетическую структуру. Генофондом популяции называют совокупность генотипов всех особей популяции. Под генетической структурой популяции понимают соотношение в ней различных генотипов и аллелей.
Одними из основных понятий популяционной генетики являются частота генотипа и частота аллеля. Под частотой генотипа (или аллеля) понимают его долю, отнесенную к общему количеству генотипов (или аллелей) в популяции. Частота генотипа, или аллеля, выражается либо в процентах, либо в долях единицы (если общее количество генотипов или аллелей популяции принимается за 100% или 1). Так, если ген имеет две аллельные формы и доля рецессивного аллеля а составляет ¾ (или 75%), то доля доминантного аллеля А будет равна ¼ (или 25%) общего числа аллелей данного гена в популяции.
Большое влияние на генетическую структуру популяций оказывает способ размножения. Например, популяции самоопыляющихся и перекрестноопыляющихся растений существенно отличаются друг от друга.
Впервые исследование генетической структуры популяции было предпринято В.Иоганнсеном в 1903 г. В качестве объектов исследования были выбраны популяции самоопыляющихся растений. Исследуя в течение нескольких поколений массу семян у фасоли, он обнаружил, что у самоопылителей популяция состоит из генотипически разнородных групп, так называемых чистых линий, представленных гомозиготными особями. Причем из поколения в поколение в такой популяции сохраняется равное соотношение гомозиготных доминантных и гомозиготных рецессивных генотипов. Их частота в каждом поколении увеличивается, в то время как частота гетерозиготных генотипов будет уменьшаться. Таким образом, в популяциях самоопыляющихся растений наблюдается процесс гомозиготизации, или разложения на линии с различными генотипами.
Большинство растений и животных в популяциях размножаются половым путем при свободном скрещивании, обеспечивающем равновероятную встречаемость гамет. Равновероятную встречаемость гамет при свободном скрещивании называют панмиксией, а такую популяцию — панмиктической.
| Закон Харди-Вайнберга |
В 1908 г. английский математик Г.Харди и немецкий врач Н.Вайнберг независимо друг от друга сформулировали закон, которому подчиняется распределение гомозигот и гетерозигот в панмиктической популяции, и выразили его в виде алгебраической формулы.
Частоту встречаемости гамет с доминантным аллелем А обозначают p, а частоту встречаемости гамет с рецессивным аллелем а — q. Частоты этих аллелей в популяции выражаются формулой p +q = 1 (или 100%). Поскольку в панмиктической популяции встречаемость гамет равновероятна, можно определить и частоты генотипов.
P 2 + 2pq + q 2 = 1.
АА + 2Аа + аа = 1
Пользуясь этими формулами, можно рассчитать частоты аллелей и генотипов в конкретной панмиктической популяции.
Однако действие этого закона выполняется при соблюдении следующих условий:
© неограниченно большая численность популяции;
© все особи могут свободно скрещиваться друг с другом;
© все генотипы одинаково жизнеспособны, плодовиты и не подвергаются отбору;

© прямые и обратные мутации возникают с одинаковой частотой или настолько редко, что ими можно пренебречь;
© отток или приток новых генотипов в популяцию отсутствует.
В реально существующих популяциях выполнение этих условий невозможно, поэтому закон справедлив только для идеальной популяции. Несмотря на это, закон Харди-Вайнберга является основой для анализа некоторых генетических явлений, происходящих в природных популяциях. Например, если известно, что фенилкетонурия встречается с частотой 1:10000 и наследуется по аутосомно-рецессивному типу, можно посчитать частоту встречаемости гетерозигот и гомозигот по доминантному признаку. Больные фенилкетонурией имеют генотип q 2 (аа) = 0,0001. Отсюда q = 0,01. p = 1 — 0,01 = 0,99. Частота встречаемости гетерозигот равна 2pq, равна 2 х 0,99 х 0,01 ≈ 0,02 или около 2%. Частота встречаемости гомозигот по доминантному и рецессивному признакам: АА = p 2 = 0,99 2 ≈ 98%, аа = 0,01%.
| Факторы, изменяющие генетическую структуру популяции |
Изменение равновесия генотипов и аллелей в панмиктической популяции происходит под влиянием постоянно действующих факторов, к которым относятся:
© дрейф генов и другие.
Именно благодаря этим явлениям возникает элементарное эволюционное явление — изменение генетического состава популяции, являющееся начальным этапом процесса видообразования.
Популяция — это совокупность особей одного вида, длительное время обитающих на определенной территории, свободно скрещивающихся друг с другом, имеющих общее происхождение, определенную генетическую структуру и в той или иной степени изолированных от других таких совокупностей особей данного вида. Популяция не только единица вида, форма его существования, но и единица эволюции. В основе микроэволюционных процессов, завершающихся видообразованием, лежат генетические преобразования в популяциях.
Изучением генетической структуры и динамики популяций занимается особый раздел генетики — популяционная генетика.
С генетической точки зрения, популяция является открытой системой, а вид — закрытой. В общей форме процесс видообразования сводится к преобразованию генетически открытой системы в генетически закрытую.
Каждая популяция имеет определенный генофонд и генетическую структуру. Генофондом популяции называют совокупность генотипов всех особей популяции. Под генетической структурой популяции понимают соотношение в ней различных генотипов и аллелей.
Одними из основных понятий популяционной генетики являются частота генотипа и частота аллеля. Под частотой генотипа (или аллеля) понимают его долю, отнесенную к общему количеству генотипов (или аллелей) в популяции. Частота генотипа, или аллеля, выражается либо в процентах, либо в долях единицы (если общее количество генотипов или аллелей популяции принимается за 100% или 1). Так, если ген имеет две аллельные формы и доля рецессивного аллеля а составляет ¾ (или 75%), то доля доминантного аллеля А будет равна ¼ (или 25%) общего числа аллелей данного гена в популяции.
Большое влияние на генетическую структуру популяций оказывает способ размножения. Например, популяции самоопыляющихся и перекрестноопыляющихся растений существенно отличаются друг от друга.
Впервые исследование генетической структуры популяции было предпринято В.Иоганнсеном в 1903 г. В качестве объектов исследования были выбраны популяции самоопыляющихся растений. Исследуя в течение нескольких поколений массу семян у фасоли, он обнаружил, что у самоопылителей популяция состоит из генотипически разнородных групп, так называемых чистых линий, представленных гомозиготными особями. Причем из поколения в поколение в такой популяции сохраняется равное соотношение гомозиготных доминантных и гомозиготных рецессивных генотипов. Их частота в каждом поколении увеличивается, в то время как частота гетерозиготных генотипов будет уменьшаться. Таким образом, в популяциях самоопыляющихся растений наблюдается процесс гомозиготизации, или разложения на линии с различными генотипами.
Большинство растений и животных в популяциях размножаются половым путем при свободном скрещивании, обеспечивающем равновероятную встречаемость гамет. Равновероятную встречаемость гамет при свободном скрещивании называют панмиксией, а такую популяцию — панмиктической.
| Закон Харди-Вайнберга |
В 1908 г. английский математик Г.Харди и немецкий врач Н.Вайнберг независимо друг от друга сформулировали закон, которому подчиняется распределение гомозигот и гетерозигот в панмиктической популяции, и выразили его в виде алгебраической формулы.
Частоту встречаемости гамет с доминантным аллелем А обозначают p, а частоту встречаемости гамет с рецессивным аллелем а — q. Частоты этих аллелей в популяции выражаются формулой p +q = 1 (или 100%). Поскольку в панмиктической популяции встречаемость гамет равновероятна, можно определить и частоты генотипов.
P 2 + 2pq + q 2 = 1.
АА + 2Аа + аа = 1
Пользуясь этими формулами, можно рассчитать частоты аллелей и генотипов в конкретной панмиктической популяции.
Однако действие этого закона выполняется при соблюдении следующих условий:
© неограниченно большая численность популяции;
© все особи могут свободно скрещиваться друг с другом;
© все генотипы одинаково жизнеспособны, плодовиты и не подвергаются отбору;
© прямые и обратные мутации возникают с одинаковой частотой или настолько редко, что ими можно пренебречь;
© отток или приток новых генотипов в популяцию отсутствует.
В реально существующих популяциях выполнение этих условий невозможно, поэтому закон справедлив только для идеальной популяции. Несмотря на это, закон Харди-Вайнберга является основой для анализа некоторых генетических явлений, происходящих в природных популяциях. Например, если известно, что фенилкетонурия встречается с частотой 1:10000 и наследуется по аутосомно-рецессивному типу, можно посчитать частоту встречаемости гетерозигот и гомозигот по доминантному признаку. Больные фенилкетонурией имеют генотип q 2 (аа) = 0,0001. Отсюда q = 0,01. p = 1 — 0,01 = 0,99. Частота встречаемости гетерозигот равна 2pq, равна 2 х 0,99 х 0,01 ≈ 0,02 или около 2%. Частота встречаемости гомозигот по доминантному и рецессивному признакам: АА = p 2 = 0,99 2 ≈ 98%, аа = 0,01%.
| Факторы, изменяющие генетическую структуру популяции |
Изменение равновесия генотипов и аллелей в панмиктической популяции происходит под влиянием постоянно действующих факторов, к которым относятся:
© дрейф генов и другие.
Именно благодаря этим явлениям возникает элементарное эволюционное явление — изменение генетического состава популяции, являющееся начальным этапом процесса видообразования.
Posted in Биология Tags: Основы генетики
Популяция — это элементарная единица эволюции. Под этим термином понимают совокупность индивидов одного вида, которые связаны общим происхождением, общностью территории, способностью свободно скрещиваться и общностью генофонда. В результате естественного отбора в популяции преобладают организмы, которые обладают определенными фенотипами, а также, как следует из этого, и определенными генотипами. Такие генотипы, отдельные гены или их сочетания широко распространяются в популяции.
Предметом изучения генетики популяций являются не генотипы отдельных особей, а частоты генов (аллелей) и частоты генотипов. При анализе процессов, происходящих в популяции, мы рассматриваем не отдельные индивиды и скрещивания между этими индивидами, а наследование в больших совокупностях организмов, которые часто могут быть неоднородны по своему генотипическому составу. Вся совокупность генов особей, входящих в популяцию, образует ее генофонд. В этой области генетики чрезвычайно важно проследить в популяции динамику частот генов, аллелей, генотипов с течением времени.
В популяционной генетике важное значение имеет понятие идеальной популяции, под ней понимают такую популяцию, которая будет бесконечно велика по численности, в которой могут осуществляться свободные скрещивания (панмиксия) во всех возможных комбинациях организмов и генотипов и при этом не действуют никакие внешние факторы (например, отсутствуют мутационный процесс, нет миграции особей из одной популяции в другую, отбора, случайного дрейфа генов, избирательности скрещиваний и изоляции). Естественно, таких популяций в природе не существует, тем не менее, введение понятия такой модельной системы позволяет понять закономерности, действующие на микроэволюционном (т.е. популяционном) уровне. Говоря о том, что в идеальной популяции не действуют внешние факторы, в действительности подразумевают существование равновесия в противоположных направлениях процессов. Так, частота возникновения прямых мутаций (например А х а) должна быть равна частоте появления обратных мутаций (а х А). В таком случае суммарный результат будет выглядеть как отсутствие мутационного процесса.
Аналогично обстоит дело и с миграциями. Доля (или частота) эмигрантов определенного генотипа (особей, уходящих из популяции) (например, АА) должна быть равна доле иммигрантов (особей, приходящих в данную популяцию). На уровне частот аллелей и генотипов такое выравнивание частот выглядит как отсутствие миграции.
Изменение частот генов (аллелей) или генотипов в идеальных, или менделевских, популяциях описывается основным законом популяционной генетики — законом Харди-Вайнберга. Согласно этому закону в такой популяции частоты аллелей в ряду последовательных поколений не меняются и остаются постоянными. Такое состояние популяции часто называют равновесным.
Если обозначим частоту аллеля А через pA, a частоту аллеля а как qa, то pA + qa = 1.
Соотношение генотипов в популяции в этом случае будет рассчитываться как (pA+qa) 2 =p 2 aa+2pAqa+q 2 a=1, в чем можно легко убедиться, если рассмотреть решетку Пеннета:
Гаметы самцов ⇒
Гаметы самок ⇓
Такое соотношение генов, аллелей и генотипов будет поддерживаться в популяции неопределенно долгое время. Иными словами, популяция может находиться в равновесии неограниченное число поколений, начиная с первого. Если знать частоты генотипов, можно рассчитать частоты аллелей и наоборот, а следовательно, можно предсказать соотношение фенотипов.
Главное следствие из закона Харди-Вайнберга — это существование рецессивных аллелей преимущественно в гетерозиготном состоянии. Закон Харди-Вайнберга рассматривает микроэволюционные процессы, которые действуют на видовом или популяционном уровнях.
Факторы, которые влияют на частоты генотипов, генов и аллелей, называют факторами динамики частот генов (аллелей) в популяциях. Действуя в популяции, они изменяют соответствующие частоты.


- Инбридинг(близкородственное скрещивание). Это еще одно следствие ограничение численности популяции и нарушения панмиксии. Он приводит к:
1.увеличению доли гомозигот;
2.фенотипическому проявлению рецессивных аллелей;
3.инбредной депрессии ослаблению особей, т.к. рецессивные мутации обычно имеют отрицательное влияние на приспособленность;
4.возрастанию изменчивости, т.к. в гомозиготное состояние могут выходить не один, а многие аллели.
-
Изоляция. В результате изоляции в популяциях могут закрепляться разные аллели генов или устанавливаться неодинаковые их частоты, особенно, если в них действует отбор в разных направлениях. Разобщение непрерывного потока генов происходит в результате действия изолирующих механизмов. Они могут иметь самую разнообразную природу. Она может быть географической, пространственной и биологической. Биологические виды изоляции бывают физиологическими (например, неодновременное созревание половых клеток у разных организмов, может привести к подразделению популяции на две или большее число субпопуляций), экологическими (каждая субпопуляция занимает отдельную экологическую нишу) и генетическими (полиплоидия, хромосомные перестройки, ядерно-цитоплазматическая несовместимость, несовместимость генов в результате возникновения в них мутаций и др.).

Что такое популяция
Популяция - это термин, который происходит от латинского — populatio, т.е. население. Он обозначает совокупность организмов относящихся к одному виду, которые длительное время обитают на определённой территории (имеют общий ареал обитания) и полностью либо частично изолированны от особей других аналогичных групп того же вида.

Для чего нужна популяционная генетика
Интерес изучения генетики на популяционном уровне заключается в том, что через популяционный уровень осуществляется генетическая преемственность поколений, а также регуляция таких биологически важных свойств, как численность, плодовитость, устойчивость к заболеваниям и пр.
Таким образом, популяционная генетика — это раздел генетической науки, который изучает распределение частот аллелей (различных форм одного и того же гена, расположенных в одинаковых участках хромосом), а также их изменение под влиянием движущих сил эволюции, определяющих генетическую структуру популяции.
Она является одним из главных элементов синтетической теории эволюции. Механизмы, которые определяют генетическую структуру популяций, можно разделить на две группы:
- поддерживающие равновесие,
- те, которые, его нарушают.
К первым относятся естественный отбор и дрейф генов, а ко вторым мутагенез и поток генов. При этом естественный отбор и мутагенез оказывают систематическое воздействие, а дрейф и поток генов – случайное. Тем самым, популяционная генетика объясняет процессы адаптации и видообразования .
В общем исследования в области генетики популяций можно условно разделить на два направления:
- описание генетического состава популяций
- анализ причин изменения генофонда популяции.
Закон Харди-Вайнберга
Рассмотрение каждого из механизмов определяющих генетическую структуру популяции, заслуживает отдельной статьи, но понимание их невозможно без рассмотрения ключевого закона популяционной генетики – закона Харди-Вайнберга или закона равновесного состояния. На нем и остановим своё внимание.
Итак, механизмы этого закона был открыты в 1908 г. английским математиком Харди (Hardy) и немецким врачом (Weinberg) независимо друг от друга.
Для его понимания рассмотрим такое понятие как случайное скрещивание.
Случайное скрещивание – это скрещивание при котором генетическая структура особей не влияет на вероятность формирования брачных пар между ними. Таким образом, в случайно скрещивающихся популяциях частота спариваний носителей различных генотипов пропорциональна доле, частотности данных генотипов в популяции.
Итак, согласно закону Харди-Вайнберга при отсутствии элементарных эволюционных процессов (естественного отбора, мутаций, дрейфа генов и миграции) частоты генов из поколения в поколение остаются неизменными. Соответственно если скрещивания случайны, частота генотипа связана простым квадратичным соотношением с частотой генов (аллелей). Если речь идет об аутосомном локусе, то равновесие генов достигается за одно поколение, и если дополнительные вмешательства, нарушающие структуру популяции, отсутствуют, сохраняется во всех последующих поколениях. Если же мы говорим о сцепленных с полом локусах, равновесные частоты генотипов устанавливаются постепенно.

Годфри Харолд Харди (1877-1947)
Таким образом равновесные частоты генотипов определяются произведениями частот соответствующих аллелей. В случае наличия только двух аллелей А (с частотой встречаемости p) и а (с частотой - q), частоты каждого их трех возможных генотипов будут выражены уравнением:
(p + q) 2 = p 2 + 2pq + q 2 = 1.
Если мы говорим о трех аллелях А, а, А’ с частотами p, q, r, формула будет иметь вид:
(p + q + r) 2 = p 2 + q 2 + r 2 + 2pq + 2pr + 2qr = 1.
Этот прием возведения в квадрат многочлена может быть использован для определения равновесных частот генотипов при любом числе аллелей. При этом сумма частот генотипов (как и аллелей) всегда должна равняться единице.

Задачи на закон Харди-Вайнберга не всегда просто даются начинающим генетикам. Помните, что Вы всегда можете обратиться к нашим авторам за квалифицированной и своевременной помощью! Сделайте процесс своего обучения приятным и легким вместе с Zaochnik!
Иван Колобков, известный также как Джони. Маркетолог, аналитик и копирайтер компании Zaochnik. Подающий надежды молодой писатель. Питает любовь к физике, раритетным вещам и творчеству Ч. Буковски.

Из видеоурока вы узнаете о том, что такое популяция. О том, что популяциям свойственна изменчивость, которая приводит не только к разнообразию в популяции, но и к образованию новых подвидов и видов. Также вы узнаете о том, что в популяциях имеются большие запасы аллелей, которые не приносят им какой-либо пользы в данном месте или в данное время. Они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными. В данном уроке приводятся следующие понятия: популяция, естественный отбор, популяционная генетика, генофонд, мутационный процесс, гетерозиготность, изменчивость популяции.
В данный момент вы не можете посмотреть или раздать видеоурок ученикам
Чтобы получить доступ к этому и другим видеоурокам комплекта, вам нужно добавить его в личный кабинет, приобретя в каталоге.
Получите невероятные возможности



Конспект урока "Популяция. Генетический состав популяций"
Вид — это исторически сложившаяся совокупность популяций, особи которых обладают наследственным сходством морфологических, физиологических и биохимических особенностей, могут свободно скрещиваться и давать плодовитое потомство, приспособлены к определённым условиям жизни и занимают определённую область — ареал.
В природе совокупность особей вида, группируются в популяции. Популяции населяют какие-то отдельные небольшие участки ареала.
Популяция — это группа одновидовых организмов, занимающих определённый участок территории внутри ареала вида, свободно скрещивающихся между собой и частично или полностью изолированных от других популяций.

Вильгельм Людвиг Иогансен
Численность популяций, да и количество особей в популяциях из-за изменения каких-либо факторов, например климатических, может меняться. Но сами популяции как таковые все равно сохраняются.
Конечно, природа не стабильна и внутри популяций могут возникать мелкие группировки, в которые объединяются особи со сходным поведением или на основе родственных связей. Такие группировки известны нам как стайки рыб или воробьёв прайды львов. Напомним, что прайды ― это семейные группы львов. В которые входят один или нескольких взрослых самцов (обычно не более 3-х), нескольких половозрелых самок и их детёнышей (львят обоих полов). Численность прайда может достигать до 30−40 животных. В прайдах всегда доминируют львы — альфа-самцы.
Но, как ни странно, охотой занимаются только самки. Молодые самцы по достижении половой зрелости изгоняются из прайда. Живя отдельно, они крепнут, взрослеют, а затем создают свои прайды.

В отличие от популяции, группы не способны устойчиво поддерживать сами себя. Они могут смешиваться с другими или распадаться под влиянием каких-либо факторов.
Популяция же занимает ту часть ареала, в которой отсутствуют изоляционные барьеры.
Каждая популяция приспосабливалась (адаптировалась) к среде своего обитания. Характерные черты этого приспособления передаются из поколения в поколение.

Взаимоотношения организмов в популяциях
Составляющие популяцию организмы связаны друг с другом различными взаимоотношениями.
Часто особи одной популяции конкурируют друг с другом за те или иные виды ресурсов. Например, самка гиены рожает от 1 до 3 детенышей. Если оба детеныша самки, то они начинают сражение за доминирование, используя зубы. Эта конкуренция ведет к гибели почти 25% всех гиен на их первом месяце жизни. В результате оставшийся в живых детёныш получает больше различных ресурсов и созревает быстрее.
Особи одной популяции могут поедать друг друга. Например, речной окунь в отсутствии корма питается собственной молодью.
Также особи одного вида могут группироваться и вместе обороняться от хищника.
Таким образом, можно сказать что внутренние взаимоотношения в популяциях парой очень сложны и противоречивы.
В природе действует естественный отбор. Ослабленные организмы погибают. Это улучшает качественный состав популяции (в том числе качество наследственного материала, которым располагает популяция). Естественный отбор повышает способность популяции к выживанию в меняющихся условиях среды.
Сильные организмы занимают место погибших, размножаются и дают начало новому поколению.

В пределах каждой популяции организмов, размножающихся половым путём, идёт постоянный обмен генетическим материалом. А вот скрещивание особей из разных популяций происходит гораздо реже, поэтому генетический обмен между разными популяциями ограничен.
Существование в форме популяции повышает внутреннее разнообразие вида, его устойчивость к местным изменениям условий жизни, позволяет ему закрепляться в новых для себя условиях.
Генетический состав популяции
Научные генетические открытия и теории Дарвина привели к рождению особого направления исследований — популяционной генетики.
Популяционная генетика позволяет объяснить процессы изменения генетического состава популяций, возникновения новых свойств организмов и их закрепление под воздействием естественного отбора.
Каждая популяция характеризуется своим специфическим набором генов (генофондом) с присущим только данной популяции соотношением частот встречаемости разных аллелей.
Генофонд — это совокупное количество генетического материала, который слагается из генотипов отдельных особей.
Как уже отмечалось, природные популяции в разных частях ареала вида обычно более или менее различны.
Почему популяции одного вида могут отличаться? Дело в том, что чаще всего скрещивание между особями одного вида происходит в пределах одной популяции, которая находится на определённой территории.
В каждой достаточно долго существующей совокупности особей могут спонтанно возникать различные мутации, которые в дальнейшем комбинируются (перемешиваются) с разными, уже имеющимися наследственными свойствами.
Под действием мутаций, которые закрепляются в генофондах популяций, изменяются виды, что ведёт к эволюции организмов.
Поэтому можно сказать, что мутационный процесс — это постоянный источник наследственной изменчивости.
В популяции, состоящей из нескольких миллионов особей, в каждом поколении может возникать по несколько мутаций практически каждого гена, который имеется в этой популяции.
Наследственная изменчивость обусловлена возникновением разных типов мутаций и их комбинаций, которые передаются по наследству и впоследствии проявляются у потомства.
Природные популяции насыщены самыми разнообразными мутациями. На это обратил внимание ещё русский учёный Сергей Сергеевич Четвериков, который установил, что значительная часть изменчивости генофонда скрыта от глаз, так как подавляющее большинство возникающих мутаций рецессивны и не проявляются внешне.
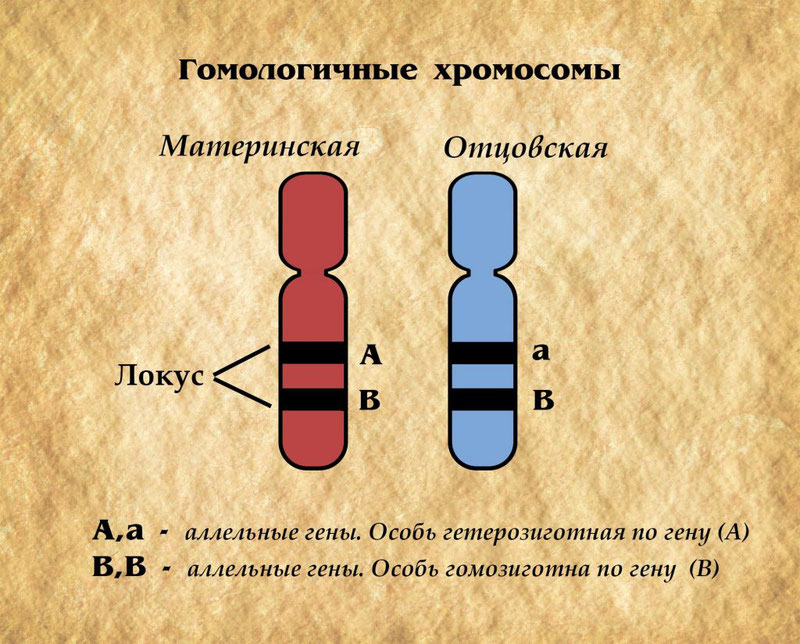
Вспомним, что гетерозиготность ─ это присущее всякому гибридному организму состояние, при котором его гомологичные хромосомы несут разные формы (аллели) того или иного гена или различаются по взаиморасположению генов.

Ген, который способен подавлять действие другого гена в онтогенезе, называется доминантным.
Ген, который оказывается подавленным и не может проявить себя в фенотипе, называется рецессивным.
Обычно доминантный ген обозначается прописной буквой ─ А-большое, рецессивный ген (а-малое). Рецессивный ген проявляется в фенотипе только в тех случаях, когда он находится в паре с аналогичным геном (а-малое, а-малое), т.е. когда оба аллельных гена являются рецессивными.
Если один аллельный ген является доминантным, а второй рецессивным (А-большое, а-малое, то в фенотипе проявляются только признаки, свойственные доминантному гену (А-большое).

Рецессивные мутации можно выявить в экспериментах со скрещиванием близкородственных особей.
При таком скрещивании некоторые рецессивные аллели, находившиеся в гетерозиготном и потому скрытом состоянии, перейдут в гомозиготное состояние и смогут проявиться.
Таким образом, благодаря мутациям популяции могут изменяться.
Изменчивость популяции — это важный фактор эволюции. Она повышает внутреннее разнообразие вида. Позволяет закрепиться виду в новых местах обитания.
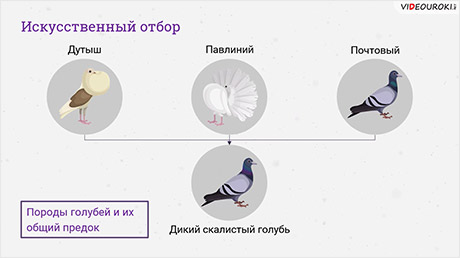
Генетическую изменчивость можно увидеть в ходе искусственного отбора. Когда из популяции выбирают тех особей, у которых какие-либо ценные в хозяйственном отношении признаки выражены наиболее сильно, и скрещивают этих особей между собой.
В фенотипе первого поколения в результате такого скрещивания проявляются необходимые наилучшие выбранные признаки.
Например, у моркови и свёклы ценятся только сладкие корнеплоды, у картофеля — крупные клубни, у мака ― нераскрывающиеся коробочки.
Следовательно, в популяциях возможна генетическая изменчивость буквально по каждому признаку данного организма.
Ещё Сергей Сергеевич Четвериков показал, что большинство вновь возникших мутаций оказываются вредными и, как правило, снижают жизнеспособность особей.
Они сохраняются в популяциях лишь благодаря отбору в пользу гетерозигот.
Однако мутации, вредные в одних условиях, могут повысить жизнеспособность в других условиях.

Так, мутация, вызывающая недоразвитие или полное отсутствие крыльев у насекомых, безусловно, вредна в обычных условиях, и бескрылые особи быстро вытесняются нормальными. Но на островах, где дуют сильные ветры, такие насекомые имеют преимущества перед особями с нормально развитыми крыльями.
Поскольку всякая популяция обычно хорошо приспособлена к своей среде обитания, крупные изменения обычно снижают эту приспособленность.
В популяциях имеются большие запасы таких аллелей, которые не приносят ей какой-либо пользы в данном месте или в данное время; они сохраняются в популяции в гетерозиготном состоянии, пока в результате изменения условий среды вдруг не окажутся полезными.
Как только условия среды изменяются, частота новых аллелей под действием отбора начинает возрастать, и в конечном счёте они становятся основным генетическим материалом.
Таким образом, благодаря приспособлениям к новым условиям среды, популяции адаптируются.
Например, к изменению климата, появлению нового хищника или конкурента и даже к загрязнению среды человеком.
Примером подобной адаптации служит эволюция видов насекомых, устойчивых к инсектицидам. При введении в практику нового инсектицида (яда, действующего на насекомых) для успешной борьбы с насекомым-вредителем бывает достаточно небольшого его количества.
С течением времени концентрацию инсектицида приходится повышать, пока, наконец, он не оказывается недейственным.
Гены, способные обеспечить устойчивость к инсектицидам, очевидно, имелись в каждой из популяций этих видов; их действие и обеспечило в конечном итоге снижение эффективности ядов, использованных для борьбы с вредителями.
Таким образом, мутационный процесс создаёт материал для эволюционных преобразований, формируя резерв наследственной изменчивости в генофонде каждой популяции и виде в целом. Поддерживая высокую степень генетического разнообразия популяций, он создаёт основу для действия естественного отбора и микроэволюции.

Разбираемся с темой популяционной генетики: ее определение, для чего нужна, кто является основателем и главные этапы развития этого раздела биологии.
Что такое популяционная генетика
Это раздел генетики, который занимается изучением генофонда целых популяций и его изменений под влияний движущих сил эволюции, пытается объяснить различные процессы адаптации и формирования видов. Популяционная генетика является одной из важнейших составляющих синтетической теории эволюции.
Для чего нужна популяционная генетика
Необходимость изучения раздела строится на идее о том, что именно через популяционный уровень происходит генетическая преемственность поколений. А значит, что и такие важные биологические свойства, как численность, плодовитость, устойчивость к заболеваниям, тоже можно отслеживать и регулировать.
Существуют определенные механизмы, которые определяют генетически структуру популяций. Их можно разделить на две группы:
- Поддерживающие равновесие. К ним относятся естественный отбор и дрейф генов. Естественный отбор — основной фактор эволюции, в результате которого поддерживается и увеличивается число тех особей, кто сумел приспособиться под новые условия. Дрейф генов — случайные изменения частот аллелей (различные формы одного и того же гена) и генотипов, происходящие в небольшой полиморфной популяции при смене поколений.
- Нарушающие равновесие. Это мутагенез и поток генов. Мутагенез отвечает за процесс изменения в нуклеотидной последовательности ДНК, который в дальнейшем приводит к мутациям. Поток генов — это перенос аллелей генов из одной популяции в другую.
Кто является основателем метода исследования

На становление популяционной генетики повлияли Сьюэл Райт, Джон Холдейн, Рональд Фишер и Сергей Четвериков. Однако ключевые закономерности, определяющие частоты аллелей в популяции, были определены Годфри Харди и Вильгельмом Вайнбергом.
Закон Харди-Вайнберга гласит: в популяции бесконечно большого размера, в которой не действует естественный отбор, не идет мутационный процесс, нет обмена особями с другими популяциями, не осуществляется дрейф генов, а скрещивания случайны — частоты генотипов по какому-либо аллельному гену будут поддерживаться на одном уровне из поколения в поколение. Такое утверждение соответствует формуле:
\(p^2 + 2pq + q^2 = 1,\) где
\(p^2\) — доля гомозигота по одному из аллелей,
\(p\) — частота этого аллеля,
\(q^2\) — доля гомозигота по альтернативному аллелю,
\(q\) — частота соответствующего аллеля,
\(2pq\) — доля гетерозигот.

Этапы развития популяционной генетики
История становления этого раздела генетики состоит из четырех основных этапов, которые затрагивают определенные промежутки времени и имеют свое значение:
- Вторая половина 1920-х — конец 1930-х годов. Период характеризуется накоплением информации о генетической гетерогенности популяций. Итогом стало появление представлений о полиморфизме (способность некоторых организмов существовать в состояниях с различной внутренней структурой или в разных внешних формах) популяций.
- 1940-е - середина 1960-х годов. На этом этапе осуществляется изучение механизмов поддержания генетического полиморфизма популяций. Появляются представление о важности гетерозиса в становлении генетического полиморфизма.
- Вторая половина 1960-х — конец 1970-х. Широко применяется белковый электрофорез для изучения полиморфизма популяций. Происходит формирование идей о нейтральном характере эволюции.
- С конца 1970-х годов. Происходит смещение в сторону ДНК-технологий для изучения особенностей процессов, осуществляющихся в популяциях. Одним из важнейших моментов этого этапа является начало широкого применения вычислительной техники и специальных программ для анализа различных типов генетических данных.
Остались вопросы? Не справляешься с контрольной работой, курсовой или дипломом? Пиши специалистам ФениксХелп, они помогут разобраться с работой любой сложности.
Читайте также: