
Газообмен у ныряющих животных кратко
Обновлено: 01.07.2024
Чтобы читать весь документ, зарегистрируйся.
Связанные рефераты
газообмен
. Содержание 1. Характеристика системы газообмена и особенности конструкции.
Газообмен
. Газообмен в легких и тканях. Кислород в основном транспортируется к тканям в составе.
газообмен в легких
. План: Актуальность Введение Газообмен в легких:транспорт газов кровью и обмен газов в.
газообмен в лёгких
. Новосибирский Медицинский Колледж Реферат Тему: « Газообмен в Лёгких. Жизненная ёмкость.
газообмен в лёгких
. Медицинский Колледж Реферат Тему: « Газообмен в Лёгких. Жизненная ёмкость.
Сам факт пребывания животных с легочным типом газообмена под водой указывает на их высокую толерантность к недостатку кислорода.
Ныряние становится возможным на фоне адаптации к дефициту кислорода. Млекопитающие способны находиться под водой от нескольких секунд до двух часов.
Китообразные при нырянии погружаются на большую глубину. Кашалотов обнаруживали на глубине до 1,5 км. Подсчеты показывают, что большую часть жизни киты проводят под водой, т. е. в классическом смысле не дышат. Зубатые киты дольше находятся под водой по сравнению с усатыми китами. После часового погружения у кашалота наблюдают 30-40 фонтанов (выдохов) с интервалом в 25-30 с.
У дельфинов дыхательный акт является безусловно-рефлекторным, действующим при смене среды вода-воздух. Благодаря такой регуляции дыхания дельфин продолжает дышать на поверхности воды даже тогда, когда находится в бессознательном состоянии. В целом надо отметить, что ритм дыхания ныряющих животных отличается нерегулярностью, т. е. имеет прерывистый характер. А погружение под воду у большинства животных, включая сухопутных (кошки, собаки, куры) рефлекторно вызывает апноэ — задержку дыхания.
У кашалота вдох и выдох — процессы произвольные. Кит начинает выдох еще находясь под водой.
Длительное пребывание под водой на большой глубине в условиях отсутствия атмосферного воздуха предполагает наличие у животных ряда морфофункциональных адаптаций. Так, относительный объем легких у хороших ныряльщиков значительно больше по сравнению с сухопутными животными. Например, у дельфинов масса легких достигает 4% от массы тела. Это в два с лишним раза больше по сравнению с наземными животными. К примеру, масса легких у крупного рогатого скота равна 0,7%, у свиньи — 0,9%, у лошади — 1,0%, у собаки — 1,4%, у домашней кошки — 1,9%.
Количество легочных альвеол у дельфинов в три раза больше, чем у наземных животных. Как верхние дыхательные пути, так и нижние у китообразных имеют прочные стенки благодаря включению жестких хрящевых колец. Кроме того, у ныряющих животных толстая плевра. В легких, включая альвеолы, обнаруживаются мышечные элементы. Эта важная структурная особенность дыхательных путей, легких и плевральной полости предохраняет воздух от сжатия при нырянии на большую глубину. В противном случае у ныряющих животных после погружения развивалась бы кессонная болезнь.
Еще одна интересная особенность заключается в том, что у вторично водных животных трахея и пищевод разделены физически на всем протяжении. Поэтому при нырянии животные могут питаться без угрозы захлебнуться.
Легкие у вторично водных животных отличаются не только повышенной емкостью, но и высокой эластичностью. Благодаря этим особенностям ныряльщики имеют увеличенную фракцию дыхательного воздуха в составе жизненной емкости легких. Например, если у наземных млекопитающих дыхательный воздух занимает около 10% от жизненной емкости легких, то у водных 80-90%.
Большой объем легких и их противостояние сдавливанию обеспечивает не только полную разовую вентиляцию легких, но и положительную плавучесть телу вторично водных животных. Это для них важно, поскольку у китов, тюленей и других ныряющих животных относительно тяжелый костяк. В костях превалирует компактное вещество при малой доле более легкого губчатого вещества. Масса костей составляет 25-30% от живой массы водных животных.
Усатые киты имеют дополнительное приспособление для запасания воздуха — воздушный мешок. Эти звери имеют одну (левую) ноздрю, через которую происходит заполнение легких воздухом. Правая ноздря сильно расширена и превращена в воздушный мешок, емкость которой равна емкости легких, т. е. 7-8 тыс. л у бутылконоса и 12-15 тыс. л у кашалота.
Заполнение воздушного мешка воздухом происходит следующим образом. У поверхности воды кит производит выдох. После этого следует вдох через левую ноздрю с заполнением легких свежим воздухом. Затем левая ноздря закрывается, а воздух из легких перекачивается в воздушный мешок. Далее вдох повторяется и происходит окончательное заполнение легких атмосферным воздухом.
У ныряющих животных имеются и другие морфологические и физиологические адаптации к нырянию. Так, у многих видов китообразных, ластоногих, грызунов исследователи отмечают более высокую кислородную емкость крови.
Но это справедливо не для всех ныряющих животных. Так, кислородная емкость крови китов даже ниже, чем у наземных животных. Большая кислородная емкость крови ныряльщиков создается за счет повышенного содержания кислорода в одном эритроците, а не за счет повышения общего количества красных клеток, как в высокогорных условиях. Средние показатели гемоглобина ныряльщиков не намного выше показателей наземных животных: кашалот — 15-17%; сейвал — 14-16%; ламантин — 12-13%.
Тем не менее общие запасы кислорода в крови у ныряющих животных выше, чем у наземных. Этому способствует особое строение артериальной системы. У вторично водных животных имеются специфические образования — артериальные сплетения, в которых депонируется насыщенная оксигемоглобином кровь. Поэтому общая кислородная емкость крови, например, у тюленя в 2 раза выше, чем у человека.
У ныряльщиков имеется и более надежный способ создания запаса кислорода. Исследования показали, что у китов и ластоногих содержание миоглобина в несколько раз выше, чем у наземных млекопитающих. Так, у тюленя и быка различия по этому показателю достигают 7-кратной величины. В отдельных мышцах концентрация миоглобина у вторично водных животных в 10 раз превышает таковую у домашней собаки.
Сердце у водных животных находится в условиях, которые предполагают выполнение меньшего объема работы (низкая гравитация, преобладание горизонтальных прямолинейных магистральных сосудов). Поэтому сердечный индекс у водных животных меньше, чем у сухопутных.
Крупные киты в хороших условиях питания накапливают большую массу жировой ткани. Поэтому за счет жировых накоплений в теле у них сердечный индекс понижается до 0,2-0,3%.
Несмотря на относительно малые размеры сердца вторично водных животных, функционально их сердце выигрывает у сердца сухопутных животных. Проявляется это в увеличенном систолическом выбросе и минутном объеме крови. Например, у тюленя после 10 погружений под воду систолический объем возрастает с 38 мл до 100 мл, а минутный — с 5,5 л до 14,7 л (А. Д. Слоним, 1971). У наземных животных эти характеристики сердца могут увеличиваться при функциональных нагрузках менее, чем в два раза.
Строение сердца ныряющих животных имеет особенности. Прежде всего бросается в глаза симметрия в развитии левой и правой половин. Если у собаки толщина стенки правого желудочка составляет всего 33% от средней толщины стенки левого желудочка, то у ладожского тюленя эта разница более выражена и равняется 64%. У гренландского тюленя левый и правый желудочки равны по объему и по массе. А у китов-гигантов правый желудочек больше левого. Объясняется это тем, что разветвленная капиллярная сеть огромных легких создает большое сопротивление току крови. Преодолеть это сопротивление позволяет мощная систола правого желудочка, который выглядит гипертрофированным на фоне левой половины сердца.
Ныряющие животные при нырянии проявляют функциональную брадикардию. При погружении под воду у крупных китов частота сердечных сокращений снижается с 30 до 15 ударов в минуту. У касаток при нырянии работа сердца замедляется с 60 до 30 ударов.
У тюленей брадикардия еще более выражена. У них частота сокращений сердца после погружения под воду рефлекторно понижается в 10 раз — с 60 до 6 ударов в минуту.
При нырянии вследствие апноэ у китов и тюленей диафрагма передавливает заднюю полую вену. В результате кровь скапливается в венозных сосудах, сплетениях и пазухах, разгружая таким образом сердечную мышцу. При этом ограничивается кровоснабжение и мышц тела. Скелетные мышцы переходят на потребление тканевого кислорода, который запасается при помощи миоглобина. К тому же под водой у ныряющих животных резко снижается мышечный тонус, что предполагает снижение потребления кислорода и выделения углекислого газа.
Экспериментальные исследования, проведенные на тюленях, показали, что при насильственном погружении животных под воду в первую минуту отмечается повышение газообмена. Однако далее наступает длительное снижение потребления кислорода тюленем. В итоге за время пребывания под водой животные расходуют значительно меньше кислорода, чем на поверхности воды, т. е. одной из важнейших особенностей ныряющих животных является снижение кислородного запроса организма.
У ныряющих животных отмечена низкая чувствительность мозга к углекислому газу и снижению pH крови. Ныряющие животные остаются под водой до тех пор, пока не иссякнут запасы кислорода, не реагируя при этом на накопление в крови СО2. Наконец, необходимо обратить внимание читателя и на то, что у вторично водных животных не развивается кессонная болезнь — профессиональное заболевание водолазов, ловцов жемчуга и аквалангистов. Причина заключается в том, что до определенных глубин воздух, заполняющий воздухоносные пути и легкие, в силу высокой прочности стенок дыхательных путей и тела животных в целом не подвергается сжатию. Поэтому растворимость азота в крови не увеличивается. Однако на больших глубинах (1000 м и более) давление воды на тело ныряльщика составляет большую величину — 100-150 атм. Тем не менее, это не вызывает насыщения крови азотом, поскольку при нырянии у животных прекращается дыхание. Болезнь развивается только при активной вентиляции легких.
воздухом, пузырьки которого задерживаются между волосками тела:
При этом передко возникают >, в которых посто
янно идет газообмен воздуха в пузырьке с окружающей водой. Эrот
процесс компенсирует уменьшение кислорода в пузырьке за счет
дыхания. Водные лwшнки некоторых насекомых (например, стрекоз)
кнутых трахей, контактирующих с водной средой; и в этом случае дыхание идет с nомощью трахей.
Среди позвоночных все вторично-водные виды имеют легоЧНЬIЙ тип дыхания. Внешний газообмен у них nроходит в нормальных
условиях, однако во время пребывания под водой легочное дыхание
nрекращается, и на тканевом уровне nостепенно развивается состояние
асфиксической гипоксии. У наиболее специализированных ныряль
щиков остановка дыхания может быть довольно длительной, но у
подавляющего большинства видов nродолжительность пребывания под водой измеряется минутами.
Среди рептилий наибольшая (более 1 ч) длительность нахоЖдения
бородавчатых змей Acrochordus granulatus время естественного заныри
вания доходит почти до 2 ч, а тихоокеанская зеленая череnаха Chelonia mydas вьщерживает поrружение до 5 ч. У других водных рептилий
продолжительность активных поrружений обычно не превьШlает 2025 мин. Время пребывания под водой резко увеличивается при спячке.
Отметим, что у водных рептилий в дополнение к легочному дыха
нию хорошо выражено и водное с помощью сосудистых сплетений в
коже, а также в слизистых оболочках ротовой и анальной nолостей.
Соответственно этому, как показано для водной змеи Acrochordus granulatus, во время продолжительного (более 30 мин) ныряния кровь
передвигается только по системным сосудам, минуя легкие; nри воз
душном дыхании восстанавливается нормальный кровоток.
Лабораторные эксперименты с триониксом Trionyx sinensis nоказа
ли, что в погруженном состоянии организм этой черепахи около 70 %
кислорода получает через сосуды глотки и около 30 % -через кожу.
Среди млекопитающих наиболее длительное пребывание под водой
характерно для настоящих вторично-водных форм -китообразных и ластоногих. Продолжительность nоrружений у ряда видов китов ко
леблется от 30 мин (финвал Balaenoptera physalis) до 2 ч (бутьmконос
Hyperodon ampu/atus). Дельфины совершают длительные серии доволь
но коротких ныряний; у косатки, например, продолжительность от
дельноrо заныривания обычно не превышает 4-5 мин. Ластоногие чаше находятся под водой 15-30 мин (тюлень Узддела -до 45 мин).
Полуводные млекопитающие выдерживают пребыванис под водой не
более 15 МИН (ондатра, бобр); бегемоть1, ламантины, дюгони, хотя и
проводят в водоемах весьма длительное время, в nолностью поrружен
ном состоянии могут вынести лишь несколько (обычно тоже не более
Птицы по длительности поrружения в большинстве случаев усту
пают водным млекопитающим. Ныряние буревестников, некоторых
\'"ТИНЫХ. оляпки длится примерно до 15 с, у других утиных, а также у
Гагар и поганок до 30-40 с. При насильственном погружении в воду
(хотя бы только головы) тупик способен задержать дыхание до 4 мин, а чиски и кайры -до 12 мин. Пинrвины (Pygosce/is papua, Р. adeliae, Eudiptes chriso/ophus) при свободном нырянии на глубину до 20 м
находятся под водой 3-7 мин; императорский пинrвин Aptenody fnrsteri -до 18 мин. Последний вид чередует глубокие (100-200 м.
~tакси~tум 305) и мелкие (около 25 м) ныряния длительностью соот
ветственно 4--6 и 1-2,5 мин.
Ныряние вызьшает довольно быстрое уменьшение содержания кислорода и нарастание концентрации со2 в крови. Устойчивость
ныряющих ~отных к такой функциональной гипоксии и геперкап
нии определяется рядом физиолоrnческих адаптаций, направленных,
с одной стороны, на увеличение запасов кислорода в организме перед
нырянием, а сдругой - на более экономное расходование этих запасов во время вынужденной остановки дыхания.
Запасание О2 перед нырянием. Запасы кислорода обычно связывают
с накоплением его в крови и мышцах, поскольку объем его в легких относительно невеm1к. Многие ныряющие животньiе уходят в воду на вьuохе, 'ПО более выгодно, так как снижает плавучесть и затраты энергии на погружение. Для глубоко ныряющих животных это, кроме того, препятствует возникновению «кессонной болезни>>. У финвала, например, объем легких сос'
бу1ъL1коноса -даже 1 %. У человека этот показатель составляет 7 %.
Инструментальные наблюдения показали, что тюлени (Halichoerus
кrypus, Phoca Ьitulina, Mirounga angustiros, Leptonychotes weddelil) ныряют
при заполнении легких на 60-20 % от их общего объема.
В результате относительная роль кислорода воздуха, заполняющего
.lегкис, в общем его запасании на время погружения оказьmается
второстепенной (табл. 6.8). У обыкновенного тюленя из общего запаса кислорода в организме (40 мл/кr) только 14 мл/кr приходится на долю
-lегких, тогда как у человека и собаки соотношение обратное -только
1 О \IJljкr приходится на нелегочные запасы.
Утки и, видимо, большинстводругихптиц также ныряюr на выдохе. Только ШIЯ пингвинов (Pygoscelis adeliae, Р. papua) доказано ныряние
на вдохе; при кратковременности погружения, свойственной этим
видам, такая ситуация не грозит им опасным насьПIIением крови
инертными газами. В то же время эксперименrально показано, что
запас воздуха в дыхательной системе позволяет этим птицам продол
жать легочный газообмен во время ныряния.
Т а б л и ц а 6.8. Кислородные запасы а орrанизме нwр11юших JКRBOПIWX и человека,
Газообмен в легких. Переход в легких кислорода из альвеолярного воздуха в кровь и поступление углекислого газа из крови в легкие подчиняется описанным выше закономерностям. Благодаря работам великого русского физиолога Ивана Михайловича Сеченова стало возможным изучение газового состава крови и условий газообмена в лёгких и тканях.
Газообмен в легких совершается между альвеолярным воздухом и кровью путем диффузии. Альвеолы легких оплетены густой сетью капилляров. Стенки альвеол и стенки капилляров очень тонкие, что способствует проникновению газов из легких в кровь и обратно. Газообмен зависит от поверхности, через которую осуществляется диффузия газов, и разности парциального давления (напряжения) диффундирующих газов. Такие условия есть в легких.
Разность между напряжением газов в венозной крови и их парциальным давлением в альвеолярном воздухе составляет для кислорода 110-40=70 (мм рт. ст.), а для углекислого газа 47-40=7 (мм рт. ст.). Скорость диффузии углекислого газа из крови в 25 раз больше, чем кислорода, поэтому за счет разности в 7 мм рт. ст. углекислый газ успевает выделиться из крови.
Газообмен в тканях. Обмен газов в тканях также происходит за счет диффузии вследствие разности напряжений кислорода и углекислого газа в крови и тканях.
Перенос газов кровью. Переносчиком кислорода от легких к тканям и углекислого газа от, тканей к легким является кровь. В крови, как и во всякой жидкости, газы могут находиться в двух состояниях: в физически растворенном состоянии и в химически связанном виде. И кислород, и углекислый газ в очень небольшом количестве растворяются в плазме крови. Основные количества кислорода и углекислого газа переносятся в химически связанном виде.
Основным переносчиком кислорода является гемоглобин крови. Каждый грамм гемоглобина связывает 1,34 мл кислорода. Гемоглобин обладает способностью вступать в соединение с кислородом, образуя оксигемоглобин. Чем выше парциальное давление кислорода, тем больше образуется оксигемоглобина. В альвеолярном воздухе парциальное давление кислорода 100-110 мм рт. ст. При этих условиях 97% гемоглобина крови связывается с кислородом. В виде оксигемоглобина кислород кровью приносится к тканям. Здесь парциальное давление кислорода низкое и оксигемоглобин (соединение непрочное) высвобождает кислород, который используется тканями. На связывание кислорода гемоглобином оказывает влияние и напряжение углекислого газа. Углекислый газ уменьшает способность гемоглобина связывать кислород и способствует диссоциации оксигемоглобина. Повышение температуры также уменьшает возможности связывания гемоглобином кислорода. Известно, что температура в тканях выше, чем в легких, которые все время вентилируются. Все эти условия помогают диссоциации оксигемоглобина, в результате чего кровь отдает высвободившийся из химического соединения кислород в тканевую жидкость.
Свойство гемоглобина связывать кислород имеет жизненно важное значение для организма. Иногда люди гибнут от недостатка кислорода в организме, окруженные самым чистым воздухом.. Это может случиться с человеком, оказавшимся в условиях пониженного давления (на больших высотах), - где в разреженной атмосфере очень низкое парциальное давление кислорода. 15 апреля. 1875 года воздушный шар "Зенит", на борту которого находились три воздухоплавателя, достиг высоты 8000 м. Когда воздушный шар приземлился, то живым был только один человек, остальные погибли. Причиной гибели было резкое снижение величины парциального, давления кислорода на большой высоте. На больших высотах (7-8 км) артериальная кровь по газовому составу приближается к венозной; все ткани тела начинают испытывать острый недостаток кислорода, что приводит к тяжелым последствиям. Подъем на высоту более 5000 м, как правило, требует пользования специальными кислородными приборами.
При специальной тренировке организм способен приспосабливаться к пониженному содержанию кислорода в атмосферном воздухе. Углекислый газ переносится кровью в виде химических соединений - гидрокарбонатов натрия и калия. Связывание углекислого газа и отдача его кровью зависят от его напряжения в тканях и крови. Кроме того, в переносе углекислого газа участвует гемоглобин крови.
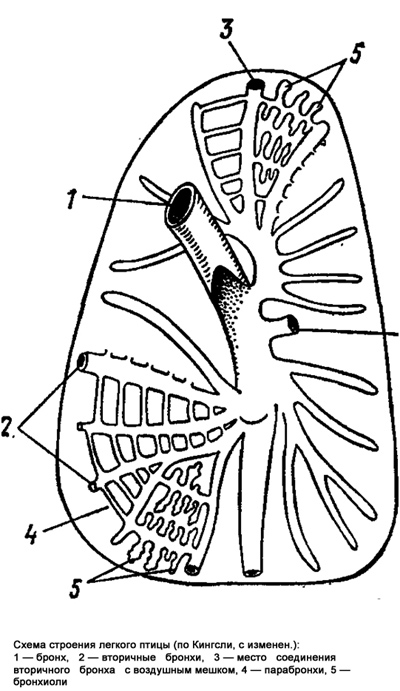
Особенности дыхания птиц и ныряющих животных. Птицы не имеют диафрагмы, поэтому расширение и сжатие легких зависит от действия межреберных мышц. В отличие от легких млекопитающих у птиц два бронха проходят насквозь через легкие и заканчиваются брюшными воздухоносными мешками. При полете птиц особое значение имеет подмышечный воздушный мешок. Во время вдоха воздух поступает частично в воздухоносные мешки, а частично в тончайшие разветвления бронхов, где совершается газообмен. При выдохе воздух из воздушных мешков направляется к бронхам и трахее. Таким образом, благодаря воздушным мешкам птицы получают атмосферный воздух, как при вдохе, так и при выдохе.
От воздухоносных мешков идут отростки, некоторые из них проникают в длинные кости конечностей. Воздухоносные мешки участвуют в поддержании нормального положения тела птицы при полете. Это достигается путем перемещения центра тяжести в зависимости от степени наполнения воздухом тех или иных воздухоносных мешков. Воздухоносная система птиц является весьма вместительным резервуаром для воздуха. Количество воздуха в системе таково, что оно может обеспечить газообмен даже при отсутствии дыхательных движений. Такой резерв воздуха необходим птицам при полете.
При полете птиц благодаря усиленной деятельности грудных мышц образуются значительные количества тепла. Полагают, что воздухоносные мешки благодаря усиленной вентиляции и большой поверхности испарения способствуют охлаждению тела птиц и предохраняют их от перегревания.
У ныряющих животных (киты и др.) увеличены длина и объем грудной полости, сильно развита дыхательная мускулатура. Носовые отверстия расположены на верхней части головы, что дает возможность животным дышать, не высовывая головы из воды. Воздух из носовых ходов попадает лишь в трахею и бронхи. Во время ныряния носовые отверстия закрываются, и вода не попадает в дыхательные пути. В легких ныряющих животных альвеол в три раза больше, чем в легких крупных сельскохозяйственных животных. Кашалот может не подниматься на поверхность воды 80-100 мин, синий кит -20-30 мин.
Регуляция дыхания. Дыхание животных регулируется. Оно может быть частым, редким, более глубоким или поверхностным в зависимости от многих факторов. Изменяется дыхание под влиянием импульсов, приходящих к дыхательной мускулатуре по центробежным нервам, или же под влиянием химических веществ, образующихся в организме.
Французским исследователем Флурансом и русским физиологом Н. А: Миславским было установлено, что в продолговатом мозге имеется группа клеток, разрушение которых ведет к остановке дыхания. Так было положено начало изучению дыхательного центра. Дыхательный центр - сложное образование, состоящее из центра вдоха и центра выдоха. Позже удалось показать, что дыхательный центр имеет более сложную структуру и в процессах регуляции дыхания принимают участие также и вышележащие отделы центральной нервной системы, которые обеспечивают приспособительные изменения в системе органов дыхания к различной деятельности организма. Важная роль в регуляции дыхания принадлежит коре больших полушарий головного мозга.
В дыхательном центре ритмически возникают импульсы возбуждения. Эти импульсы возникают автоматически. Даже после полного выключения приходящих к дыхательному центру по центростремительным нервным волокнам импульсов в нем можно зарегистрировать ритмическую активность. Автоматизм дыхательного центра связывают с процессами обмена в нем. Ритмические импульсы передаются из дыхательного центра по центробежным нейронам к дыхательным мышцам и диафрагме.
Изменение дыхания происходит рефлекторно. Дыхание меняется при болевом раздражении, при раздражении органов брюшной полости, рецепторов кровеносных сосудов, кожи, рецепторов дыхательных путей.
Особое значение в регуляции дыхания имеют импульсы, идущие от рецепторов дыхательных мышц и от рецепторов самих легких. Импульсы от легких обеспечивают чередование вдоха и выдоха. Это происходит так. При вдохе, когда легкие растягиваются, раздражаются рецепторы в стенках легкие Импульсы от рецепторов легких по центростремительным волокнам достигают дыхательного центра, тормозят центр вдоха и возбуждают центр выдоха. В результате дыхательные мышцы расслабляются, грудная клетка опускается, диафрагма принимает вид купола, объем грудной клетки уменьшается и происходит выдох. Поэтому говорят, что вдох рефлекторно вызывает выдох. Во время выдоха легкие спадаются, при этом рефлекторно тормозится центр выдоха и возбуждается центр вдоха.
В регуляции дыхания принимает участие кора головного мозга. С ее помощью и обеспечивается тончайшее приспособление дыханий к потребностям организма в связи с изменением условий внешней среды и жизнедеятельности организма.
Большое влияние на состояние дыхательного центра оказывает химический состав крови, в частности ее газовый состав. Впервые это было показано в опыте с перекрестным кровообращением. У двух собак, находящихся под наркозом, перерезали сонные артерии, несущие кровь к голове, и через трубки соединяли так, что кровь от туловища первой собаки поступала к голове второй, а кровь от туловища второй собаки поступала к голове первой. Пережатие трахеи у первой собаки вызывало одышку у второй, и наоборот. Происходило это потому, что в крови собаки, у которой переживали трахею, накапливалось большое количество углекислого газа. Такая кровь, насыщенная углекислым газом, поступала к голове другой собаки, омывала ее и возбуждала к активной деятельности дыхательный центр. Поэтому у второй собаки и наблюдались частые дыхательные движения (одышка). Последующие опыты с введением в определенный участок продолговатого мозга раствора, содержащего углекислоту, подтвердили это положение.
Литература: Хрипкова А. Г. и др. Физиология животных: Учеб. пособие по факультатив. курсу для учащихся IX-X кл./ А. Г. Хрипкова, А: Б. Коган, А. П. Костин; Под ред. А. Г. Хрипковой. - 2-е изд., перераб.- М.: Просвещение, 1980.-192 с., ил.; 2 л. ил.
Читайте также: