
Гаструляция у птиц кратко
Обновлено: 05.07.2024
Бластуляция зигот птиц. Этапы гаструляции у птиц
Бластодерма центрального отдела зародышевой мишени, в связи с ее быстрым ростом в ширину, постепенно истончается. Ее клетки, которые сначала не прилегают тесно друг к другу и располагаются слоями, нередко с видимыми щелями между ними, организуются затем по типу однослойного эпителия. При этом зародышевый валик на будущем каудальном конце зародыша более толст, чем на будущем краниальном конце.
Собственное тело зародыша развивается из центрального округлого участка бластодермы (зародышевого щитка или эмбриональной зоны, area embryonalis), отчасти прозрачного и распространяющегося над субгерминативной полостью (аналогичной бластоцелю) на площади, соответствующей приблизительно ее размерам. Эта область бластодермы является более темной и более прозрачной и называется блестящей зоной (area pellucida).
Кнаружи от этой зоны лежит область бластодермы, которая соответствует краевому отделу, ограничивающему со всех сторон субгерминативную полость; таким образом, эта область располагается над зародышевым валиком и представляет собой светлое непрозрачное поле (area ораса). Наконец, по самой периферии лежит слой, соответствующий еще не раздробленной желточной массе (area vitellina).
При дальнейшем развитии дискобластулы пресмыкающихся и птиц сначала возникает энтодерма, которая, однако, в данном случае уже не может развиваться посредством инвагинации и поэтому образуется иным, новым путем. В то время как у некоторых видов пресмыкающихся (например у черепах) и у некоторых рыб еще наблюдаются кое-какие черты инвагинирующего процесса, то у птиц инвагинация отсутствует полностью.
Измененные условия существования, наряду с большим содержанием желтка, сделали у этих животных невозможным образование энтодермы путем инвагинации и обусловили возникновение принципиально иного вида гаструляции.
Энтодерма у пресмыкающихся и у птиц возникает в результат процесса, при котором происходит отделение клеток и клеточных совокупностей от бластодермы (отчасти и от каудального конца зародышевого щитка из отдела зародышевого валика) и их распределение в сплошной клеточный слой, благодаря чему первоначальная бластодерма расщепляется на поверхностную эктодерму и расположенную под ней энтодерму.
Таким образом, гаструляция в данном случае происходит по новому способу — путем деляминации (расщепления) энтодермального слоя.
У пресмыкающихся таким образом возникает двуслойный зародыш — дискогаструла, у птиц — эпигаструла.
После окончания данного процесса развития наступает нотогенез, происходящий в области блестящей зоны (area pellucida), а именно в ее центральной части (в зародышевом щитке, то есть в эмбриональной зоне). У пресмыкающихся в задней части этого зародышевого щитка образуется узелковое утолщение эктодермы — примитивная или первичная ротовая пластинка.
В этой пластинке клеточный материал начинает проникать вглубь, благодаря чему возникает карманоподобное образование — примитивный, первичный карман, проникающий между эктодермой и энтодермой. Полость этого кармана отчасти соответствует полости первичного кишечника, и ее можно считать несовершенным результатом процесса инвагинации. Первичный или иначе хордомезодермальный карман у пресмыкающихся растет в краниальном направлении, причем края отверстия в его полость соответствуют передней губе бластопора.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
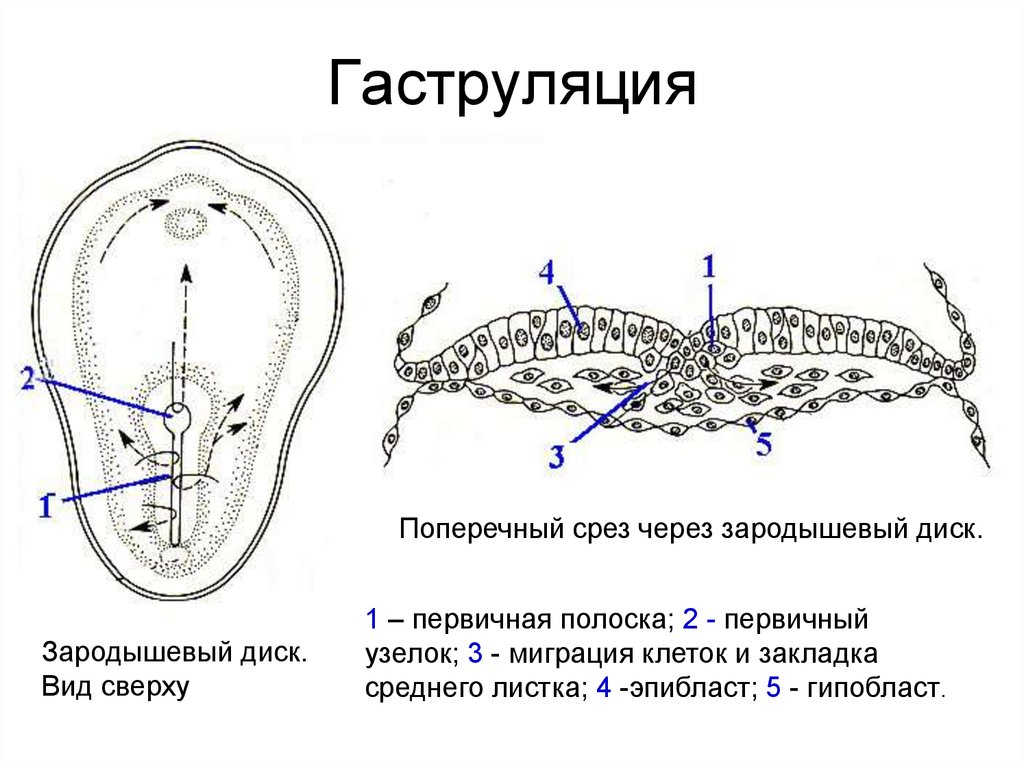
Гаструляция (от лат. gaster – желудок) – процесс превращения бластулы в гаструлу, сопровождающийся усиленной пролиферацией и направленным перемещением клеток с одновременно протекающей в них химической и морфологической перестройкой (дифференцировкой). Морфогенетические перемещения клеток довольно сильно варьируют в разных классах позвоночных. В процессе гаструляции из однослойной бластулы образуется сначала два зародышевых листка - (эпи - и гипобласт), а затем вследствие продолжающихся перемещений клеток формируется третий зародышевый листок - мезодерма и осевые зачатки (рис.3).
Рис. 3. Зародыш курицы на стадии поздней гаструлы и образования осевых зачатков. 1 – эктодерма; 2 – энтодерма; 3 – мезодерма; 4 – нервная трубка; 5 – хорда; 6 – целом; 7 – аорта.
После этого эпибласт называется эктодермой, а гипобласт – энтодермой. Различают 4 способа преобразования бластулы в гаструлу. Они тесным образом зависят от типов яйцеклеток, а, следовательно, типов дробления и бластул:
1. Инвагинация (впячивание) – характерна для животных с олиголецитальными первичными изолецитальными яйцеклетками (ланцегник). При этом вегетативный полюс целобластулы впячивается в бластоцель, в результате чего образуется вначале двухслойный зародыш, Состоящий из экто - и энтодермы, а затем вследствие таких же инвагинационных преобразований из материала внутреннего зародышевого листка (энтодермы) формируется третий листок – мезодерма.
2. Эпиболия (обрастание) – происходит у животных с мезолецитальными яйцеклетками. Мелкие, дробящиеся более ускоренно, бластомеры анимального полюса амфибластулы обрастают макробластомеры вегетативного полюса, в результате чего образуется экто - и энтодерма.
3. Иммиграция (перемещение). Популяции бластомеров перемещаются в полость бластулы и образуются зародышевые листки.
4. Деламинация (расслоение). Зародышевые листки образуются в результате расщепления клеток эмбрионального диска на 2 слоя.
У животных с мезолецитальными и полилецитальными яйцеклетками обычно наблюдается сочетание нескольких типов гаструляции, которые могут протекать или одновременно, или сменять друг друга.
Кратко остановимся на характеристике гаструляции у птиц и млекопитающих. У них гаструляция протекает в две стадии.
В течение первой стадии образовавшийся эмбриональный диск (зародышевый щиток) путем деламинации разделяется на 2 листка – эпибласт (первичная эктодерма) и гипобласт (первичная энтодерма). В состав эпибласта входит материал вторичной эктодермы, зародышевой и внезародышевой мезодермы и хорды. Гипобласт включает материал кишечной и желточной энтодермы. Эпибласт является дном формирующегося амниотического пузырька, а гипобласт – крышей образующегося желточного мешка.
Сразу после образования эпибласта и гипобласта из их состава в полость, ограниченную трофобластом, выселяются клетки внезародышевой мезодермы, в дальнейшем преобразующиеся в мезенхимоциты, которые вместе с трофобластом формируют стенку хориона – ворсинчатую оболочку зародыша.
Вторая стадия гаструляции характеризуется направленным перемещением быстро размножающихся клеток эпибласта спереди назад, к центру и вглубь. В результате такой миграции в сочетании с инвагинацией клеток образуются первичная полоска (с первичной бороздкой), являющаяся аналогом боковых губ бластопора, и первичный (головной) узелок (с первичной ямкой) – аналог дорзальной губы бластопора.
Через края первичной бороздки клетки эпибласта мигрируют вглубь эмбрионального диска и, распространяясь латерально между эпи- и гипобластом, формируют мезодерму, а головной узелок, клетки которого перемещаются через передний край первичной ямки и распространяются между эпи - и гипобластом в виде тяжа в краниальном направлении, даёт начало хорде (головному отростку).
Таким образом, у птиц (аналогично и рептилий) и млекопитающих в процессе образования зародышевых листков ведущими способами являются деламинация и миграция, а вспомогательными – инвагинация и эпиболия.
Гисто-, органо - и системогенез
В результате дифференцировки клеток зародышевых листков часть из них идет на построение тканевых и органных зачатков тела зародыша, а другая – внезародышевых органов. Формирование этих зачатков происходит на основе детерминации и коммитирования.
Детерминация – генетически запрограммированный путь развития клеток и тканей, а коммитирование – ограничение возможностей путей их развития. Эти процессы сопровождаются дальнейшей дифференцировкой клеток, приводящей к образованию тканевых зачатков.
Под дифференцировкой понимают изменения в структуре клеток, связанные с их функциональной специализацией. При этом различают 4 основных этапа дифференцировки: 1-й – оотипическая дифференцировка, когда материал будущих зачатков презумпирован в определенных участках ооплазмы или зиготы; 2-й – бластомерная дифференцировка; 3-й – зачатковая дифференцировка и 4-й этап – гистогенетическая дифференцировка.
Хорда является индуктором развития нейроэктодермы, благодаря чему материал нервной пластинки начинает инвагинировать, образуя последовательно нервный желобок и нервную трубку. Из материала энтодермы, располагающегося под нервной трубкой, формируется кишечная трубка.
Нервная трубка и кишечная трубка являются осевыми органами зародыша.
Таким образом, эмбриональный гистогенез и органогенез – это процесс превращения малодифференцированного клеточного материала эмбриональных зачатков в специализированные ткани и органы. Параллельно с этим осуществляется системогенез.
1. Формирование гипобласта.
Путем деламинации бластодерма разделяется на два слоя – верхний многослойный эпибласт, нижний однослойный первичный гипобласт. Между двумя пластами находится подзародышевая полость.
На заднем конце зародыша в эпибласте формируется скопление клеток – серп Коллара. Эти клетки мигрируют по подзародышевой полости, оттесняют на периферию первичный гипобласт и формируют вторичный гипобласт, который в дальнейшем войдет в состав внезародышевой энтодермы. Подзародышевая область образует бластоцель. Эпибласт приобретает эпителеподобный вид, бластомеры принимают призматическую форму и выстраиваются в один более или менее правильный ряд. Пространство, где клетки двух слоев разделены бластоцелем, кажется светлым и называется area pellucida. Там, где два слоя клеток прилегают друг к другу, находится area opaca, более темная область. Вышеперечисленные процессы происходят еще до откладки яйца в яйцеводе.
2. Образование трех зародышевых листков.
Происходит после откладки яйца, при насиживании.
Клетки многослойного эпибласта начинают мигрировать к центру зародышевого диска. Латеральные потоки клеток образуют первичную полоску – центрально расположенное утолщение эпибласта, которое растет от заднего конца к переднему. Полоска, продвигаясь к переднему краю, соединяется с медиальным потоком клеток и образует гензеновский узелок. Первичная полоска приобретает углубление по средней линии и с этого момента носит название первичной бороздки. На вершине первичного узелка появляется углубление – первичная ямка. Отсюда начинается миграция клеток в подзародышевую полость. Первые мигрировавшие клетки оттесняют гипобласт и образуют энтобласт (презумптивная кишечная энтодерма). Далее, клетки мигрируют через стенки первичную бороздку и гензеновский узелок в пространство между эпибластом и энтобластом с образованием мезобласта. Перемещение клеток под эпибласт из передней части первичной полоски приводит к ее укорочению и смещению к задней части зародыша.
а). Клетки гензеновского узелка через первичную ямку мигрируют в центральную переднюю часть зародыша, не распространяясь при этом вентральном направлении, и образуют головную мезодерму и хордальный вырост.
б). Клетки, мигрирующие через боковые стенки задней части бороздки и отходящие к периферии, образуют внезародышевую мезодерму.
в). поздняя иммиграция клеток через центральную часть первичной бороздки дает начало латеральной мезодерме и сомитам.
Как только клетки будущей мезодермы из состава эпибласта погрузятся в первичную бороздку они приобретают звездчатую форму, характерную для мезенхимы.
Укорочение и смещение назад бороздки приводит к образованию анального отверстия из гензеновского узелка.
По мере того, как презумптивные мезодермальные и энтодермальные клетки движутся внутрь, клетки предшественники эктодермы окружают желток посредством эпиболии.
Клетки гипобласта не принимают участия в построении тела зародыша. Они образуют часть внезародышевых оболочек органов, которые питают и защищают зародыш. Все три зародышевых листка собственно зародыша сформируется из клеток эпибласта.
Материал первичной полоски и клетки в передней части гензеновского узелка образуют средний зародышевый листок. Снаружи остаются покровная и нейральная эктодерма.
Гаструляция у птиц гомологична гаструляции у амфибий. При этом первичная полоска является бластопором, ее края – боковыми губами, а гензеновский узелок –дорсальной губой бластопора. Он же обладает свойствами первичного организатора. Однако, в отличие от клеток амфибий, которые мигрируют в бластоцель целыми пластами, клетки зародыша птиц вступают в первичную полоску индивидуально. Кроме этого, особенностью птиц как амниот является наличие внезародышевых листков, которые образуют оболочки зародыша, способствующие его развитию.

Данные по эмбриональному развитию книдарий, относящихся к низшим многоклеточным животным, часто используются для решения вопросов, связанных с происхождением и ранней эволюцией Metazoa, а также с основными закономерностями эволюции онтогенеза. Особое внимание уделяется гаструляции – морфогенетическому процессу, в ходе которого клетки раннего эмбриона дифференцируются на зародышевые листки и формируется первичный план строения. В статье проведен сравнительный анализ гаструляции различных книдарий. Мы показывали, что для гаструляционных морфогенезов книдарий характерна высокая степень межгрупповой, внутригрупповой и индивидуальной изменчивости. Мы считаем, что способ гаструляции у книдарий определяется не столько эволюционной историей рассматриваемого вида, сколько эволюционно пластичными адаптивными признаками, такими, как размер яйцеклетки и содержание в ней желтка, число клеток на стадии бластулы или морулы, наличие фототрофных симбионтов, экология личинки. Поскольку онтогенез книдарий обладает высокой эволюционной пластичностью, его изучение должно способствовать лучшему пониманию фундаментальных закономерностей эволюции процессов развития.
Как типичные представители низших Metazoa, книдарии обладают простым планом строения, который принято считать эволюционно примитивным (рис. 1). Они двухслойны: их тело состоит из экто- и эндодермы. Книдарии имеют единственную орально-аборальную ось тела, кишечную полость, открывающуюся наружу только одним (ротовым) отверстием. К числу признаков, характерных для типа Cnidaria, можно отнести также разнообразие способов бесполого размножения, высокую способность к регенерации, а также к тканевым и клеточным трансдифференцировкам (способность взрослой региональной стволовой клетки дифференцироваться в клетки другого органа и/или другого зародышевого листка). В настоящее время в типе Cnidaria выделяют пять классов: Anthozoa, Scyphozoa, Staurozoa, Hydrozoa и Cubozoa (рис. 2). Их филогенетические взаимоотношения остаются спорными.

Рис. 2. Филогенетическое дерево книдарий, демонстрирующее представителей 5 классов, выделяемых в настоящее время. Обратите внимание, что у Anthozoa в жизненном цикле имеется только жизненная форма полипа; у Staurozoa сидячая медуза неотличима от полипа; большинство представителей Scyphozoa, Cubozoa, Hydrozoa имеют обе жизненные формы; большинство представителей Anthozoa и Hydrozoa колониальны.
Несмотря на прогресс в изучении развития книдарий современными методами (в том числе методами молекулярной генетики и биоинформатики), наши знания об их эмбриональном развитии, к сожалению, часто остаются на уровне конца XIX века. Могут ли имеющиеся у нас данные по раннему развитию книдарий прояснить спорные моменты их эволюции и филогении? В статье "Гаструляция книдарий: ключ к пониманию филогенеза или хаос вторичных модификаций?" авторы отвечают на этот вопрос, анализируют накопленные к настоящему моменту сведения об эмбриональном развитии, и в первую очередь, о гаструляции различных представителей типа Cnidaria.
У книдарий существует четкая преемственность между орально-аборальной полярностью взрослого животного и полярностью яйцеклетки. Оральный полюс полипа соответствует анимальному полюсу яйца, на котором происходит отделение полярных телец, расположен женский пронуклеус и закладывается первая борозда дробления. У видов, гаструляция которых морфологически поляризована, этот полюс также является полюсом формирования эндодермы. У активно плавающей личинки планулы этот полюс становится задним концом. Из заднего конца планулы в ходе метаморфоза формируется оральный полюс полипа.
Первичный план строения Cnidaria формируется при помощи широкого спектра различающихся путей развития. Эти пути эквифинальны, так как у абсолютного большинства книдарий они приводят к одному и тому же итогу: формированию двухслойной личинки планулы (или паренхимулы, а затем планулы).
Изучение эмбрионального развития, и особенно гаструляции Cnidaria имеет очень длинную историю. Фактически, классификация типов гаструляции и первые описания многих гаструляционных морфогенезов, сделанные в XIX веке, были основаны на изучении эмбриогенеза книдарий (Metschnikoff, 1886). В тот же период данные по развитию книдарий и других низших Metazoa (губок) начинают использоваться для построения теорий происхождения многоклеточности и онтогенеза животных (Metschnikoff, 1886; Haeckel, 1874).
Одна из самых удачных классификаций типов гаструляции (гаструляционных морфогенезов) была предложена О.М. Ивановой-Казас (1975, с. 50). Эта классификация подчёркивает тесную связь типов гаструляции с морфологией бластулы, которая, в свою очередь, зависит от особенностей дробления. В статье приводится её упрощённый вариант (рис. 3).

Рис. 3. Классификация типов гаструляции (по О.М. Ивановой-Казас, с изменениями).
ар - архентерон (гастральное впячивание материала эндодермы), бп - бластопор (отверстие, через которое происходит впячивание эндодермы), бц - бластоцель (полость, формирующаяся во время дробления).
Итак, если в результате дробления получается целобластула (рис. 3, а), гаструляция может осуществляться за счёт первичной (клеточной) деламинации (рис. 3, б), униполярной иммиграции клеток (рис. 3, в), мультиполярной иммиграции клеток (рис. 3, г) и инвагинации (рис. 3, д). Если в результате дробления получается морула (плотная группа клеток) (рис. 3, е), гаструляция может осуществляться за счёт вторичной (морульной) деламинации (рис. 3, ж). Если в ходе дробления получается бластула, состоящая из клеток, резко различающихся по размеру (амфибластула, рис. 3, з), то гаструляция скорее всего будет осуществляться за счёт эпиболии (рис. 3и). Существует также довольно редкий вариант гаструляции – изгибание плакулы (рис. 3, к, л). Если целобластула по каким-то причинам имеет плоскую (дисковидную) форму, то ее можно называть плакулой (рис. 3, к), а гаструляция представляет собой последовательное преобразование этого диска в чашу и сферу с отверстием – бластопором (рис. 3, л). Интересно, что у книдарий обнаружены практически все варианты гаструляционных морфогенезов, приведенные на этом рисунке. При таком разнообразии уместно задаться вопросом, существует ли строгая связь между типом гаструляции и филогенетическим положением тех книдарий, для которых он характерен. Попробуем выяснить так ли это, рассмотрев особенности гаструляционных морфогенезов у представителей разных таксонов книдарий. В статье рассматриваются гаструляция 4-х классов книдарий - Anthozoa, Scyphozoa, Staurozoa и Hydrozoa (информации по раннему развитию Cubozoa в литературе отсутствует). Однако здесь мы остановимся (в качестве примера) на анализе данных по гаструляции Anthozoa и Hydrozoa,
Гаструляция у Anthozoa
В классе Anthozoa эмбриогенез лучше всего изучен у подкласса Hexacorallia. За последние два десятилетия было подробно изучено развитие модельных видов: актинии Nematostella vectensis и склерактинии Acropora millepora . Развитие немодельных склерактиний сейчас очень активно изучает эмбриолог Нами Окубо (Токийский Университет). Наш анализ развития этой группы в значительной степени опирается на её данные (Okubo et al., 2013). Обобщив всю имеющуюся в литературе информацию, мы выделили пять основных вариантов путей развития Hexacorallia (рис. 4, а-д):

Рис. 4. Гаструляция Anthozoa (подкласс Hexacorallia).
гц - гастроцель (гастральная полость); ик - иммигрирующие клетки; псб - псевдобластопор (впячивание стенки бластулы, которое находится в том же месте, где впоследствии сформируется бластопор); р - рот.
Насколько жестко тот или иной путь развития связан с таксономическим положением вида? Самое необычное развитие, при котором формируется плакула (вариант 5), наблюдается только у некоторых родов склерактиний. Однако при более внимательном анализе их развития выясняется, что эмбрион на прегаструляционной стадии далеко не всегда может быть с полным основанием описан как плакула: очень часто полость бластоцеля бывает хорошо выражена. Полностью соответствует варианту 5 развитие представителей лишь нескольких родов, например, рода Acropora . У других родов склерактиний рот формируется непосредственно из отверстия бластопора, а клетки архентерона (гастрального впячивания) не деэпителизуются.
Таким образом, у проанализированных таксонов Hexacorallia внутригрупповое разнообразие путей развития сопоставимо с межгрупповым. Значительная часть этого разнообразия связана с накоплением в яйцеклетках желтка, затрудняющего морфогенетические движения, а также с необходимостью транспортировки симбионтов в клетки эндодермы. При этом разнообразие самих гаструляционных морфогенезов невелико.

Рис. 5. Разнообразие гаструляционных морфогенезов Hydrozoa и их эквифинальность - успешное формирование паренхимулы (м) и планулы (н) как результат всех путей развития. Развитие начинается с формирования морулы (а, в) или бластулы (б, г); гаструляция осуществляется за счет вторичной деламинации (а → д → м; в → з → м), первичной деламинации (б → ж → м; г → и → з → м), униполярной иммиграции (г → к → м), мультиполярной иммиграции (г → л → м).
Для многих гидроидов характерна стадия морулы (рис. 3е; 5а, в, ж, з). Морфология морулы у разных видов заметно различается. Иногда морула представляет собой рыхлый агрегат клеток (рис. 5а). В этом случае гаструляция осуществляется за счет своеобразной формы вторичной деламинации. Для стадии гаструлы тогда характерны неправильная форма эмбриона, а также высокий уровень морфологической и морфогенетической изменчивости (рис. 5, д), который резко снижается при переходе к стадии паренхимулы (рис. 5м). Морула может быть плотной, и тогда вторичная деламинация, не связана с существенным изменением формы эмбриона вплоть до стадии паренхимулы (рис. 5в, з). Из паренхимулы, как правило, формируется личинка - планула, у которой эпителизована не только эктодерма (как у паренхимулы), но и эндодерма (рис. 5н).
Морула также может формироваться в процессе первичной деламинации. Так протекает развитие у гидроидов, которые формируют бластулу со слабо выраженным бластоцелем (рис. 5, б). Благодаря ориентации митотических веретен перпендикулярно поверхности эмбриона дочерние клетки отшнуровываются в полость бластоцеля (рис. 5, е, ж). В результате получается плотная морула (рис. 5з). Окончательное обособление эктодермы от эндодермы осуществляется за счет вторичной деламинации.
У других гидроидов первичная деламинация начинается на относительно поздней стадии. В этом случае успевает сформироваться бластула с хорошо выраженной полостью (рис. 5, г, и), которая постепенно также превращается в плотную морулу (рис. 5, з).
Если в ходе дробления получается бластула, гаструляция может происходить за счет униполярной или мультиполярной иммиграции (рис. 5, к, л).
Униполярная иммиграция - очень распространенный морфогенез. Он связан с униполярным формированием колбовидных клеток, которые постепенно выселяются в бластоцель (рис. 5, к). Этот процесс приводит к формированию паренхимулы (рис. 5, м). Мультиполярная иммиграция, напротив, описана для очень небольшого числа видов гидроидов.
Инвагинационный мофогенез не найден до сих пор ни у одного гидроида. Скорее всего, отсутствие инвагинации среди гаструляционных морфогенезов можно считать признаком, характерным для класса Hydrozoa.
Итак, гаструляционные морфогенезы Hydrozoa очень разнообразны. Еще в XIX веке предпринимались попытки найти закономерности, объясняющие их распространение среди представителей этого класса. Мечников обратил внимание на связь способа гастуляции с экологией эмбриональных стадий. Судя по всему, эмбрионы, ведущие подвижный образ жизни (становящиеся личинками на стадии бластулы), формируют эндодерму с помощью униполярной иммиграции. С другой стороны, эмбрионы, развивающиеся до стадии планулы в материнском организме или лежащие на дне используют мультиполярные морфогенезы (Metschnikoff, 1886). При внимательном анализе большего числа видов обнаруживается, что представители близких родов могут использовать совершенно разные способы гаструляции. Учитывая, что уже описаны виды гидроидов с индивидуально изменчивой гаструляцией, можно сделать вывод, что в классе Hydrozoa разнообразие гаструляционных морфогенезов связано скорее с экологией эмбрионального развития, чем с таксономическим положением вида, причем внутригрупповое разнообразие не уступает межгрупповому.
Заключение
Наш анализ гаструляции Cnidaria показал, что не существует дискретных, четко различающихся путей развития. Мы увидели, что разные варианты гаструляционных морфогенезов распределены по классам и отрядам книдарий в значительной степени мозаично; четкой приуроченности к тем или иным таксонам, как правило, не наблюдается. Скорее следует говорить о существовании непрерывной изменчивости способов развития.
Судя по всему, способ гаструляции у книдарий часто менялся в ходе эволюции. Он подвержен значительной внутригрупповой изменчивости, порой не уступающей межгрупповой. Возможно, способ гаструляции книдарий определяется не столько эволюционной историей рассматриваемого вида и его таксономической принадлежностью, сколько эволюционно пластичными адаптивными признаками, такими как размер яйцеклетки и содержание в ней желтка, число клеток бластулы, наличие фототрофных симбионтов, экология личинки. Более того, ход гаструляции подвержен внутривидовой (индивидуальной) изменчивости. Из этого следует, что гаструляционные морфогенезы книдарий могут иметь лишь весьма ограниченное применение при решении частных вопросов филогенетики. С другой стороны, сравнительное изучение онтогенеза низших Metazoa проливает свет на общие закономерности морфогенетических процессов и их эволюции, что крайне важно для понимания ранних этапов становления животного царства.
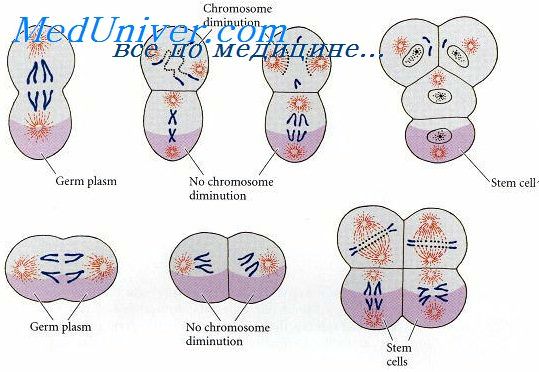
В процессе гаструляции выделяют 2 фазы: 1) фаза образования двухслойного зародыша (I фаза); 2) фаза образования трехслойного зародыша (II фаза). Различают 4 основных способа осуществления гаструляции в I фазе, заканчивающейся образованием двухслойного зародыша.


Александр Онуфриевич Ковалевский (1840-1901)
Христиан Генрих Пандер (1794-1865)
1. Инвагинация - впячивание части бластодермы (стенки бластулы) внутрь зародыша. Бластодерма вегетативного полушария впячивается внутрь так, что полюса бластодермы сближаются, а бластоцель либо исчезает, либо остаётся в виде щелевидной полости (рис. 105). В итоге из шарообразной бластулы образуется двухслойный мешок, состоящий из экто- и энтодермы, внутри которого содержится полость - гастроцель, сообщающаяся с внешней средой отверстием - бластопором. Края бластопора образуют дорсальную, вентральную и две латеральные губы (рис. 105). У первичноротых бластопор превращается в дефинитивный (окончательный) рот, у вторичноротых он преобразуется в анальное отверстие (рис. 106, 107), а рот формируется на противоположном конце зародыша.
2. Эпиболия - обрастание более быстроделящимися микромерами макромеров вегетативного полюса. Располагающиеся снаружи микромеры дают начало эктодерме, находящиеся внутри макромеры формируют энтодерму (рис. 108).
3. Иммиграция - выселение отдельных клеток бластодермы в бластоцель из одного места (униполярная иммиграция) или из разных мест (мультиполярная иммиграция). Оказавшиеся внутри после миграции в бластоцель клетки дают начало энтодерме, а остальные клетки бластодермы превращаются в эктодерму (рис. 109). Гастроцель при этом не образуется (кишечнополостные).
4. Деляминация - расслоение на наружный и внутренний пласты (экто- и энтодерму), характерна для бластулы типа морула. При деляминации митотическое веретено в клетках бластодермы ориентируется перпендикулярно поверхности бластулы, а борозда деления, в свою очередь, рас-
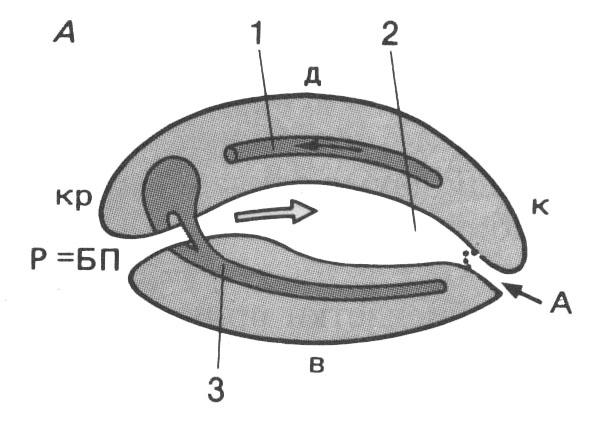

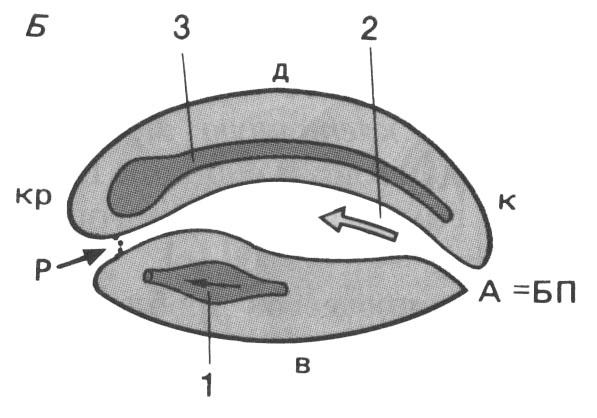
Рис. 105 (слева). Гаструляция путём инвагинации у ланцетника:
А – бластула; Б – более поздняя бластула с уплощённым нижним полюсом; В-Е - последовательные стадии гаструляции: 1 – дорсальная и вентральная губы бластопора; 2 - гастроцель
Рис. 106 (справа). Схема плана строения эмбрионов первичноротых (А) и вторичных (Б) животных:
А – анальное отверстие; БП – бластопор (первичный рот); Р – рот; кр – краниальный отдел; к – каудальный отдел; д – дорсальная сторона; в – вентральная сторона; 1 – сердце; 2 – кишка; 3 – центральная нервная система

Рис. 107. Представители первичноротых и вторичноротых животных
полагается параллельно последней. После деления клеток бластодермы происходит, по сути, её расслоение на наружный (эктодерма) и внутренний (энтодерма) листки (рис. 109).
Эти способы редко встречаются в чистом виде, обычно гаструляция происходит по смешанному типу: инвагинация сочетается с эпиболией (земноводные), деляминация - с иммиграцией (иглокожие).
Закладка третьего зародышевого листа во второй фазе гаструляции происходит:
1) телобластическим способом, имеющим место у первичноротых, т.е. всех беспозвоночных, кроме иглокожих. Мезодерма образуется из 2-х или нескольких клеток-телобластов (у моллюсков 2 телобласта располагаются симметрично в районе бластопора). Телобласты представляют собой две крупные клетки, обособляющиеся во время гаструляции с двух сторон первичной (энтодермальной) кишки. В результате их деления образуются мелкие клетки, располагающиеся между экто- и энтодермой, и формирующие мезодерму;
2) энтероцельным способом (иглокожие, ланцетник, низшие позвоночные) - путём выпячивания и отшнуровывания спинной части энтодермы первичной кишки (рис. 110). В отшнуровывающихся пузырьках (мезодермальных закладках) возникают полости как часть бывшей полости первичной кишки - гастроцеля, образующие целом (вторичную полость тела);
3) путём миграции клеток первичной эктодермы через утолщение первичной полоски (рис. 111) и последующего погружения (инвагинации) под эктодерму (пресмыкающиеся, птицы и млекопитающие).
Особенности гаструляции у амфибий. Ниже границы анимального и вегетативного полушарий бластулы образуется борозда, принимающая подковообразную форму. Затем концы её смыкаются, и образуется бластопор. Последний охватывает вегетативную зону (желточную пробку) кольцом. Клетки в области борозды бластопора погружаются внутрь, в результате чего образуется полость первичной кишки, или гастроцель (рис. 112). Через дорсальную губу мигрируют клетки будущей хорды, через вентральную - клетки формирующейся энтодермы и через латеральные губы - клетки закладывающейся мезодермы. Спустя некоторое время начинает доминировать эпиболия: микромеры, смещаясь в вегетативном направлении, как бы наползают снаружи на макромеры и, достигая бластопора, мигрируют внутрь. Из стенки первичной кишки энтероцельным способом образуется хорда и мезодерма. Начало хорде дают стенки среднего (спинного) пузырька, мезодерма формируется из стенок двух боковых пузырьков (рис. 112).
Особенности гаструляции у птиц. Бластомеры, распластанные на желтке, образуют бластодиск. Его центральная часть, ограниченная валиком, соответствует будущему зародышу и называется зародышевым

Рис. 108. Последовательные этапы гаструляции у амфибий (по Г. Шпеману)
А-Д – медиальные сагиттальные срезы; Е – стадия, соответствующая на предыдущем рисунке (Д) в поперечном разрезе; 1 – эктодерма; 2 – нервная пластинка; 3 – хордомезодерма; 4 – энтодерма; 5 – мезенхима
щитком (рис. 112). Путём расслоения (деляминации) образуются эпибласт и гипобласт (соответственно будущая экто- и энтодерма). В центре зародышевого щитка возникает утолщение - первичная полоска, имеющая первичную борозду и первичный (гензеновский) узелок (рис. 111). Клетки эпибласта мигрируют двумя потоками от периферии зародышевого щитка к первичной полоске и в области первичной борозды погружаются (подворачиваясь как бы через губу) под эктодерму (рис. 111, 113). Через гензеновский узелок мигрируют клетки будущей хорды, через среднюю часть первичной борозды – клетки, образующие сомиты, и через заднюю часть - клетки, формирующие несегментированную мезодерму. Гензеновский узелок постепенно смещается назад, образуя впоследствии анальное отверстие.
В результате гаструляции в организме: 1) создаётся первичный план строения зародыша, во многом, как правило, совпадающий с основным планом строения взрослого организма; 2) проспективные зародышевые листки, раньше граничившие лишь своими краями, теперь существуют реально и приходят в контакт своими поверхностями. Это создаёт возможность для взаимных влияний, которые служат пусковым механизмом для дальнейшего развития - возникновения различий между ранее одинаковыми клетками внутри зародышевого листка.
Читайте также: