
Гамма петля физиология кратко
Обновлено: 04.07.2024
Регуляция длины мышцы. Даже в покое скелетные мышцы редко бывают полностью расслаблены, сохраняя некоторое напряжение, не сопровождающееся признаками утомления и называемое мышечным тонусом. Спинальное животное обладает слабым мышечным тонусом. Этот тонус имеет рефлекторную природу. Доказать это можно, нарушив связь мышцы с ЦНС (спинным мозгом), перерезав задние чувствительные корешки у лягушки (опыт Бронжеста). При этом тонус мышц задней лапки исчезает, и естественное взаимоотношение бедра и голени нарушается. Следовательно, тоническое напряжение мышца сохраняет только до тех пор, пока она связана со спинным мозгом.
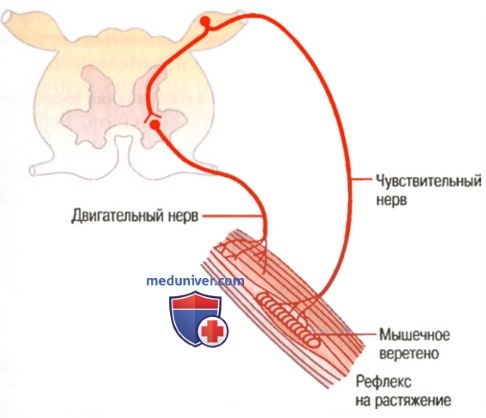
В основе тонуса лежит спинномозговой тонический или другое название - миотатический рефлекс. Это так называемый рефлекс на растяжение. Рецепторами рефлекса являются проприорецепторы самой мышцы - мышечные веретёна. Раздражителем для них служит растяжение мышцы. Вследствие того, что экстрафузальные и интрафузальные мышечные волокна расположены параллельно (о чем мы уже говорили) при растяжении мышцы удлиняются и те и другие. Происходит возбуждение спиральных нервных окончаний, по афферентным волокнам возбуждение направляется в спинной мозг, где происходит активация альфа-мотонейронов (-мотонейронов), иннервирующих экстрафузальные волокна той же мышцы.
Обратите внимание на тот факт, что в спинном мозге расположена основная структура – альфа-мотонейроны, аксоны которых являются единственным каналом, соединяющим нервную систему со скелетными мышцами. Только возбуждение альфа-мотонейронов приводит к активации мышечных волокон.
В результате возбуждения альфа-мотонейронов экстрафузальные волокна сокращаются, и растяжение мышцы уменьшается. И наоборот, когда мышца достаточно сократилась, мышечные веретёна, находящиеся в ней, укорачиваются, и импульсация от рецепторов растяжения становится слабее; уменьшается их активирующее влияние на -мотонейроны, в результате тонус мышцы снижается и её длина возвращается к исходной (рис.3)

Рисунок 3. Рефлекторная дуга спинномозгового миотатического (тонического) рефлекса
Вы видите, что это своеобразный механизм поддержания длины мышцы, в котором использована обратная связь от мышечных веретён. Этот механизм автоматически компенсирует изменение нагрузки на мышцу.
Силу, с которой мышцы сопротивляются изменению свой длины, можно определить как мышечный тонус. Он позволяет сохранять определенное положение тела или позу. Сила гравитации направлена на растяжение мышц-разгибателей, а их ответное рефлекторное сокращение этому противодействует. Если растяжение разгибателей увеличивается, например, когда на плечи опускается тяжелый груз, то и сокращение усиливается – мышцы не позволяют себя растягивать и благодаря этому сохраняется поза. При отклонениях тела вперёд, назад или в сторону определенные мышцы растягиваются, а рефлекторное повышение их тонуса сохраняет необходимое положение тела.
По тому же принципу осуществляется рефлекторная регуляция длины у мышц-сгибателей. При любом сгибании руки или ноги поднимается груз, которым может быть и сама рука или нога, но любой груз – это внешняя сила, стремящаяся растянуть мышцы. И здесь можно обнаружить, что ответное сокращение регулируется рефлекторно в зависимости от величины груза. В этом легко убедиться на практике: попробуйте перекреститься, а потом повторите эти же движения с пудовой гирей в руке, как это делали силачи в старом российском цирке.
На рис. 3 дуга спинномозгового тонического рефлекса изображена в упрощенном виде. На деле все гораздо сложнее и на рисунке 4 она представлена в полном виде.

Рисунок 4. Регуляция мышечного тонуса на спинальном уровне
1 – интрафузальное волокно
2 - ядерная сумка с нервными окончаниями чувствительного нейрона
3 – периферические участки интрафузального волокна
4 – экстрафузальное мышечное волокно
5 – α-мотонейрон спинного мозга
6 – γ- мотонейрон спинного мозга
7 – эфферентный путь от α-мотонейрона к экстрафузальному мышечному волокну
8 - эфферентный путь от γ – мотонейрона к интрафузальному волокну
Мы упоминали о том, что интрафузальные волокна имеют свою иннервацию. Она исходит от более мелких нейронов, тоже расположенных в передних рогах спинного мозга - -мотонейронов. Для чего же служат -мотонейроны, иннервирующие сам проприорецептор?
Повышают чувствительность мышечных веретён к растяжению, что способствует усилению рефлекторной реакции на изменение длины мышцы. Происходит это так: возбуждение гамма-мотонейронов вызывает сокращение интрафузальных волокон мышечного веретена и растяжение капсулы, что в свою очередь вызывает возбуждение расположенных здесь нервных окончаний и поступление от них сигналов к альфа-мотонейронам. Альфа-мотонейроны активируются, и в результате происходит сокращение экстрафузальных волокон мышцы. Активация альфа-мотонейронов через гамма-мотонейроны носит название гамма-петли (гамма-мотонейроны→мышечные веретёна→афферентные нервные окончания→альфа-мотонейроны→мышца) (рис. 5) .

Рисунок 5. Гамма петля (показана пунктиром).
Таким образом, альфа-мотонейроны можно возбудить двумя способами: рефлекторно - за счёт естественного растяжения всей мышцы и, как бы принудительно,- за счёт активации с помощью гамма-мотонейронов,
2. Поддерживая импульсацию веретён в укорачивающейся мышце, они препятствуют их полному расслаблению - в этом случае даже в укороченной при сокращении мышце веретёна будут следить за её длиной.
3. Через гамма-мотонейроны вышележащие отделы ЦНС, и в первую очередь ретикулярная формация (РФ) ствола мозга, оказывают влияние на тонус мышц. Сигнал от вышележащих двигательных центров, поступающий к альфа-мотонейронам, одновременно идёт и на гамма-мотонейроны и тем самым возбуждает интрафузальные мышечные волокна, обеспечивающие натяжение рецепторов растяжения.
Можно сказать, что одна из основных задач, выполняемых -мотонейронами и -эфферентными волокнами, заключается в настройке чувствительности измерительных приборов (веретён) для того, чтобы они могли работать при самой различной длине мышцы.
Запомните:
Cуществуют два механизма, приводящие к возбуждению мышечных веретён: 1) растяжение мышцы и 2) сокращение интрафузальных волокон (гамма-петля).
Следовательно, при управлении движениями высшие моторные уровни помимо прямых влияний на спинальные альфа-мотонейроны могут оказывать на них косвенное влияние - через гамма-петлю. Управление через гамма-петлю является важным механизмом сохранения положения звеньев тела и сохранения требуемых суставных углов. Дело в том, что при управлении, в котором используется спинальная гамма-петля, высшие моторные уровни освобождаются от непосредственного контроля за сохранением длины мышцы и, следовательно, за сохранением положения в каждом отдельном суставе. Им необходимо лишь обеспечить некоторый уровень гамма-активности и тем самым определить требуемую (эталонную) длину мышечных веретён (степень их сокращения). После этого сохранение требуемой длины мышцы может поддерживаться автоматически спинальным моторным уровнем с помощью механизма рефлекса на растяжение.
При осуществлении сложных двигательных актов происходит одновременная активация - и -мотонейронов: активируются -мотонейроны (обеспечивая само движение) и -мотонейроны (поддерживая возбуждение -мотонейронов).

Рисунок 6. Рефлекторная регуляция напряжения мышцы
Таким образом, сухожильные рефлексы играют двоякую роль: 1. Участвуют в регуляции напряжения в мышце. Когда напряжение достигает значения, при котором возбуждаются сухожильные рецепторы Гольджи, -мотонейроны, иннервирующие сокращающуюся мышцу, тормозятся, что приводит к ее расслаблению и падению в ней напряжения; 2. Способствуя быстрому расслаблению мышцы, они защищают её от повреждения при сильных сокращениях. Несмотря на это, при чрезмерных нагрузках и напряжении икроножной мышцы в ряде случаев у спортсменов и артистов балета может происходить отрыв ахиллова сухожилия от места прикрепления.
Запомните: В регуляции деятельности каждой мышцы участвуют две регулирующие системы обратной связи: система регуляции длины, роль датчика в которой играют мышечные веретёна, и система регуляции напряжения, датчиками которой служат сухожильные рецепторы.
Длина и напряжение мышцы взаимообусловлены. Если, например, вытянутая вперёд рука ослабит напряжение мышц, то уменьшится раздражение рецепторов Гольджи, а сила тяжести станет опускать руку. Это приведёт к растяжению мышц, нарастанию возбуждения интрафузальных рецепторов и соответствующей активации мотонейронов. В итоге произойдет сокращение мышц и рука вернётся в прежнее положение.
Сухожильные рефлексы названы так потому, что их можно вызвать, легко ударив неврологическим молоточком по сухожилию более или менее растянутой мышцы. От удара такая мышца растягивается и тотчас рефлекторно сокращается. В неврологической практике сухожильные рефлексы используются для оценки состояния соответствующих сегментов спинного мозга. Наиболее известный рефлекс растяжения - коленный, возникает в ответ на кратковременное растяжение четырехглавой мышцы бедра легким ударом по ее сухожилию ниже коленной чашечки. Расслабленная мышца растягивается, а возникшее в связи с этим возбуждение рецепторов мышечных веретён распространяется по рефлекторной дуге к этой же мышце. После короткого латентного периода происходит сокращение мышцы и разгибание конечности (рис. 7).

Рисунок 7. Схема рефлекторной
дуги коленного сухожильного рефлекса
Этот рефлекс отражает состояние III - IV поясничных сегментов спинного мозга, так как именно там расположены центры этого рефлекса. Кроме коленного рефлекса в клинике применяют еще целый ряд подобного рода рефлекторных проб. Например, постукивание в области локтевого сустава вызывает рефлекторное разгибание руки. Растяжению в этом случае подвергается трёхглавая мышца плеча.
Таким образом, самый низкий уровень в организации движения связан с двигательными системами спинного мозга. В спинном мозге имеются альфа-мотонейроны, которые прямо управляют мышцами; располагаются гамма-мотонейроны и вставочные нейроны (интеронейроны), образующие множество контактов с другими нервными клетками. От возбуждения вставочных нейронов зависит, будет ли то или иное движение облегчено или заторможено. На спинальном уровне осуществляется наиболее простая форма автоматического регулирования состояния мышц – рефлекс на растяжение. В его основе лежит обратная связь от мышечных веретён к альфа-мотонейронам спинного мозга.
Управление работой мышц осуществляется не отдельными нейронами, а мотонейронным пулом, который определяет силу и участие в сокращении всех или части волокон каждой мышы. Мотонейронным или двигательным пулом называются мотонейроны, иннервирующие одну мышцу и рассеянные по нескольким сегментам спинного мозга.
Рассмотренные выше механизмы поддержания мышечного тонуса осуществляются на уровне спинного мозга, поэтому такой тонус называется спинальный. Он характеризуется очень слабой выраженностью тонического напряжения мышц и не может обеспечить поддержание позы животного и акт ходьбы. Но он достаточен для осуществления простейших спинальных рефлексов.
Сгибательные и ритмические рефлексы спинного мозга. Двигательные программы спинного мозга.
В моторных системах можно выделить три типа совершаемых движений: рефлекторные, ритмические и произвольные. Они различаются по степени сложности и участия сознания в контроле выполняемых действий. В каждом из них есть два компонента, нуждающиеся в контроле: тонический, который обеспечивает необходимую для начала движения позу и на время фиксирует в стабильном положении некоторые суставы, и фазный, определяющий направление и скорость движения. Рефлекторные движения относятся к самым простым моторным действиям, они выполняются быстро, стереотипно и в принципе не нуждаются в сознательном контроле, т.е. могут возникать непроизвольно. В качестве примера можно привести все сухожильные рефлексы (такие как коленный), а также защитный сгибательный рефлекс, возникающий в ответ на прикосновение к горячему предмету.
Сто человек из ста, нечаянно прикоснувшиеся рукой к сильно нагретому предмету, немедленно согнут её, что уберегает от ещё большего повреждения. Эта стереотипная защитная реакция возникает раньше, чем осознаётся смысл происшедшего. Она обеспечивается врожденным рефлекторным механизмом, в котором участвуют болевые чувствительные окончания, сенсорный нейрон, интеронейроны спинного мозга и мотонейроны мышц-сгибателей. По такому же рефлекторному стереотипу человек, наступивший босой ногой на колючку или острый камешек, немедленно ее сгибает. Это эволюционно древний рефлекс: ведь даже лишённая головного мозга лягушка сгибает лапку, погружённую в кислоту.
Некоторые двигательные рефлексы состоят из повторяющихся ритмических движений. Например, у многих шерстистых животных в ответ на неприятное раздражение кожи паразитами наблюдается почёсывание. Шеррингтон описал чесательный рефлекс у спинальных собак, что указывает на локализацию его центра в спинном мозгу. У спинальных животных были также выявлены локомоторные рефлексы, состоящие в ритмитческой смене сокращений сгибателей и разгибателей на разных лапах, как при ходьбе. И действительно, в спинном мозге обнаружена цепь нейронов, выполняющая функции генератора шагания. Она ответственна за чередование периодов возбуждения и торможения различных мотонейронов и может работать в автоматическом режиме.
Следовательно, простые двигательные стереотипные программы спинной мозг может осуществлять самостоятельно в зависимости от характера сенсорной информации. Он может, например, прервать сгибание пальцев, берущих какой-нибудь предмет, если этот предмет окажется сильно нагретым.
Запомните! СПИННОЙ МОЗГ
---является центром тонических рефлексов растяжения, обеспечивающих поддержание длины и ограничение напряжения скелетных мышц;
---создает исходный тонус мышц, который недостаточен, чтобы обеспечить стояние и вертикальное положение головы (“антигравитационную позу”);
---осуществляет простейшие двигательные рефлексы сгибания и разгибания конечностей, шагательные движения;
---является исполнительной структурой по отношению к расположенным выше двигательным центрам, нисходящие влияния которых в конечном итоге сходятся на мотонейронах спинного мозга.
Гамма-эфферентная система мышечного сокращения. Стабилизация положения тела
а) Роль мышечных веретен в произвольной двигательной активности. Важность гамма-эфферентной системы подчеркивает тот факт, что 31% всех двигательных нервных волокон к мышцам представлены тонкими эфферентными волокнами типа Аγ, а не толстыми двигательными волокнами типа Аα. Каждый раз, когда сигналы передаются от двигательной коры или от любой другой области головного мозга к альфа-мотонейронам, в большинстве случаев одновременно стимулируются гамма-мотонейроны, что называют коактивацией альфа- и гамма-мотонейронов.
Это ведет к одновременному сокращению экстрафузальных волокон скелетных мышц и интрафузальных волокон мышечных веретен.
Сокращение интрафузальных мышечных волокон одновременно с сокращением крупных мышечных волокон скелетных мышц имеет двойное значение. Во-первых, это удерживает длину рецепторной части мышечного веретена от изменений во время сокращения всей мышцы. Следовательно, коактивация сдерживает противодействие рефлекса с мышечных веретен мышечному сокращению. Во-вторых, это сохраняет соответствующую функцию демпфирования мышечного веретена, независимо от любых изменений длины мышцы.
Например, если бы мышечное веретено не сокращалось и не расслаблялось вместе с крупными мышечными волокнами, рецепторная часть веретена была бы то слишком свободна, то перерастянута, что не соответствует оптимальным условиям для функции веретена.
Нервный контур рефлекса на растяжение
б) Области головного мозга, регулирующие гамма-эфферентную систему. Гамма-эфферентная система возбуждается непосредственно сигналами из бульборетикулярной облегчающей области ствола мозга и опосредованно — импульсами, передаваемыми в бульборетикулярную область от:
(2) базальных ганглиев;
(3) коры большого мозга.
О точных механизмах контроля гамма-эфферентной системы известно мало. Однако поскольку бульборетикулярная облегчающая область прежде всего связана с сокращениями антигравитационных мышц (а эти мышцы имеют очень высокую плотность мышечных веретен), считают, что особое значение гамма-эфферентный механизм имеет для демпфирования (сглаживания) движений разных частей тела во время ходьбы и бега.
Мышечное веретено, его связи с крупными экстрафузальными мышечными волокнами. Обратите внимание также на моторную и сенсорную иннервации мышечного веретена Детали связей с нервной системой разных волокон мышечных веретен ядерной сумки и ядерной цепочки
в) Система мышечных веретен стабилизирует положение тела во время напряженной деятельности. Одной из наиболее важных функций системы мышечных веретен является стабилизация положения тела во время напряженной мышечной деятельности. Для этого бульборетикулярная облегчающая область и связанные с ней области мозгового ствола передают возбуждающие сигналы через гамма-нервные волокна к интрафузальным мышечным волокнам.
Это укорачивает концы веретен и растягивает их центральные рецепторные области, усиливая сенсорный сигнал. Однако если веретена одновременно активируются в скелетных мышцах, расположенных по обе стороны каждого сустава, рефлекторное возбуждение этих мышц также возрастает, обеспечивая в окружении сустава сильное напряжение мышц, противодействующих друг другу. В результате положение сустава становится очень устойчивым, и любой силе, которая пытается его нарушить, противодействуют чрезвычайно чувствительные рефлексы на растяжение, действующие с обеих сторон сустава.
Каждый раз, когда человек должен выполнять мышечную работу, которая требует тонкой и точной регулировки положения тела, возбуждение соответствующих мышечных веретен сигналами из бульборетикулярной облегчающей области ствола мозга стабилизирует положение основных суставов. Это очень помогает выполнению дополнительных тонких произвольных движений (пальцами или другими частями тела), необходимых для сложных двигательных манипуляций.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Особенность двигательных нервных клеток в том, что их цитоплазма не окружает ядро равномерно, а образует два отростка. Один из них более короткий (дендрит) принимает нервный импульс, второй (аксон) передает его дальше.
Таким образом, двигательный периферический нейрон проводит нервный импульс от центральной нервной системы к мышце. В мышечной ткани его длинный отросток разветвляется и соединяется с десятками мышечных волокон.
Виды двигательных нейронов
По локализации моторные нейроны подразделяются на центральные и периферические. Центральные расположены в ткани головного мозга. Они отвечают за сознательные контролируемые мышечные сокращения.
Двигательные нейроны, идущие непосредственно к мышечным волокнам, называют соматическими.

Тела двигательных нейронов соматической нервной системы расположены в области передних рогов спинного мозга и располагаются группами, каждая из которых отвечает за сокращение строго определенной мускулатуры. К примеру, мотонейроны шейного отдела управляют мускулатурой рук, поясничного отдела отвечают за иннервацию ног.
Периферические нервные клетки, отвечающие за движения, классифицируются следующим образом:
- большие альфа-мотонейроны;
- малые альфа-мотонейроны;
- гамма-мотонейроны;
- клетки Реншо.
Большие альфа-клетки формируют крупные проводящие стволы. Малые альфа и гамма-нейроны имеют более тонкие аксоны. Клетки Реншо входят в состав крупных стволов и служат для коммутации сигналов.

Функции мотонейрона
Центральные и периферические двигательные нервные клетки работают согласовано. Совместно они обеспечивают сокращение определенных групп мышц и позволяют человеку выполнять какие-либо действия.
Для координированных движений конечностей необходимо одновременное сокращение сгибателей и разгибателей. При работе сгибателей первоначальный сигнал возбуждения возникает в области прецентральной извилины соответствующего полушария.
За это действие отвечают клетки, называемые пирамидными. Собранные вместе их отростки образуют так называемый пирамидный двигательный путь. Далее сигнал идет к передним рогам спинного мозга, откуда передается уже непосредственно в миофибриллы.
Активирующее влияние на мотонейроны мышц разгибателей оказывают специальные центры задних отделов больших полушарий. Они формируют дорсальный и вентральный пути. Таким образом, в формировании координированного движения участвуют две области головного мозга.

По характеру функции нервные клетки, задействованные в процессе мышечного сокращения, подразделяются на двигательные и вставочные нейроны. Первые ответственны за исполнительную функцию, в то время как для координации нервных импульсов служат вставочные. Эта особая разновидность имеет меньшие размеры и более многочисленна.
Для сравнения — в области передних рогов их в 30 раз больше чем двигательных. Когда возбуждение проводится по аксону двигательного нерва, оно переходит первоначально на вставочный нейрон. В зависимости от характера сигнала он может быть усилен либо ослаблен, после чего передается дальше.
Клетки вставочного типа имеют больше отростков и более чувствительны. Они обладают большим числом отростков и их еще называют мультиполярными.

Для оптимизации сигналов исходящих по аксонам и идущим к мышечным волокнам, служат специальные клетки Реншоу, которые передают возбуждение с одного отростка на другой. Такой механизм служит выравниванию интенсивности нервного сигнала.
По отростку мотонейрона импульс достигает мышечного волокна, которое сокращается. Каждая группа мотонейронов и иннервируемые ими мышечные волокна отвечают за определенные движения.
Нервные клетки, обеспечивающие двигательную функцию:
| Виды нейронов | Локализация | Функция |
|---|---|---|
| центральные иннервирующие сгибатели | область прецентральной извилины | сокращение скелетных мышц сгибателей путем передачи импульса в область передних рогов |
| центральные иннервирующие разгибатели | область заднего мозга | сокращение скелетных мышц разгибателей путем передачи импульса в область передних рогов |
| периферические альфа | передние рога спинного мозга | непосредственное сокращение скелетных мышц |
| периферические гамма | передние рога спинного мозга | регуляция тонуса |
| вставочные | все отделы ЦНС | коммуникация сигналов внутри ЦНС |
Большие альфа-нейроны, проводящие сильный импульс, вызывают сокращение миофибрилл. Малые проводят слабые сигналы и служат для поддержания тонуса мускулатуры.
Помимо волокон, отвечающих за сокращение, в мышечной ткани есть и специальные спиральные фибриллы, регулирующие силу напряжения мускулатуры.
Эти экстрафузальные мышечные волокна иннервируются гамма-нейронами.
Возбуждение гамма-мотонейрона приводит к увеличению растяжения миофибрилл и облегчает прохождение импульса сухожильных рефлексов. Примером может быть прохождение нервного сигнала по дуге коленного рефлекса.
Слаженной работой периферических мотонейронов достигается тонкая настройка мышечного тонуса, что позволяет выполнять точные координированные движения. При поражении периферических двигательных нейронов мышечный тонус исчезает и движения невозможны.

Как работает двигательный нейрон?
Для того чтобы возник биоэлектрический импульс, необходима разница потенциалов на оболочке нервной клетки. Это происходит в результате изменения концентрации ионов калия и натрия с наружной и внутренней поверхности мембраны.
В дальнейшем импульс проходит до конца длинного отростка ‒ аксона и достигает места соединения с другой клеткой. Место такого контакта называют синапс.

С другой стороны синапса к месту контакта прилежит короткий ветвящийся отросток ‒ дендрит. Передача сигнала через синапс обусловлена активными химическими веществами, так называемыми медиаторами.
Возникнув на дендрите, сигнал распространяется по его оболочке и переходит далее на аксон. Для сокращения скелетной мышцы сигнал зарождается в мотонейроне коры, проходит по пирамидному пути, переходит на вставочный нейрон и далее в область передних рогов спинного мозга. Заканчивается эта цепь в мышечной ткани.
Результатом возбуждения двигательного центра коры будет сокращение группы мышечных волокон.
Симптомы поражения центрального двигательного нейрона
Поражения центральных моторных нервных клеток возникают чаще всего при инсульте. При ишемии или кровоизлиянии в вещество больших полушарий участок ткани отмирает. Такие поражения почти всегда односторонние.
Вследствие этого при поражении центральных двигательных нейронов наблюдаются нарушения функции мускулатуры с одной стороны. Самый заметный признак ‒ это односторонний паралич, приводящий к невозможности активных движений в руке и ноге.
С этой же стороны снижается мышечный тонус в туловище и мимической мускулатуре лица. Поражение центральных моторных областей сопровождается рядом изменений рефлекторной деятельности.
Клинически это выражается в появлении разнообразных патологических рефлексов. Их совокупность, снижение тонуса мышц и нарушения чувствительности позволяют врачу установить диагноз.
О том, что такое нервно-мышечное веретено, вы наверняка узнали ещё в процессе обучения. Однако эти сложные нервные рецепторы являются важнейшими фундаментальными частями нашей нервной системы, и будет не лишним иногда повторить теорию, связанную с ними, и узнать что-то новое.
Чтобы обеспечить мышечное сокращение, центральная нервная система (ЦНС) посылает особые сигналы, вследствие чего мышца сокращается. Именно нервно-мышечное веретено передаёт ЦНС информацию, позволяющую точно корректировать различные характеристики сокращения. Эти нервные рецепторы передают проприоцептивную чувствительную информацию о длине мышцы, скорости сокращения, степени сокращения, а также о положении тела в пространстве.
Нервно-мышечное веретено состоит из нескольких видов волокон и устроено значительно сложнее, чем может показаться на первый взгляд.
Один из самых сложных функциональных элементов нервно-мышечного веретена – гамма-эфферентная система, которая включает в себя гамма-мотонейроны. В данной статье мы остановим своё внимание только на гамма-системе, поскольку в рамках одной статьи невозможно охватить всю информацию о различных сложных неврологических системах, участвующих в мышечном сокращении.
Сначала мы кратко рассмотрим анатомию нервно-мышечного веретена.
АНАТОМИЯ
Нервно-мышечные веретена расположены в скелетных мышцах между волокнами и представляют собой совокупность тонких коротких поперечнополосатых мышечных волокон, заключенных в соединительнотканную фасцию. Эти рецепторы выполняют две основные функции. Во-первых, они передают проприоцептивную информацию о длине мышц (в какой степени мышца удлинена) в центральную нервную систему. Это называется тонический ответ. Во-вторых, они передают информацию о скорости изменения длины мышцы (насколько быстро удлиняется мышца). Это называется фазовый ответ.
В мышцах имеются три типа волокон, состоящих из разных нейронов, участвующих в мышечном сокращении – альфа-, бета- и гамма-мотонейронов. Гамма-мотонейроны иннервируют особые сократительные волокна, которые называются интрафузальными. Существует два типа интрафузальных волокон – волокна с ядерной цепочкой и волокна с ядерной сумкой.
Волокна с ядерной сумкой подразделяются на два типа – первый тип, или волокно с динамической ядерной сумкой и второй тип, или волокно со статической ядерной сумкой. Эти волокна получили такое название исходя из вида организации ядер. В этих волокнах клеточные ядра располагаются ближе к центру брюшка мышцы и, скапливаясь, образуют выпуклость.
Ядра волокон с ядерной цепочкой распределены в виде длинной нити, напоминающей цепочку. Все они имеют центральную область, которая не сокращается, и окончания волокон, которые, в свою очередь, сокращаются.
Волокна первого типа, или волокна с динамической ядерной сумкой, наиболее активно реагируют на скорость изменения длины мышц. Волокна второго типа, или волокна со статической ядерной сумкой, также реагируют на скорость изменения длины мышц, однако, помимо всего прочего, они регистрируют и степень удлинения мышц.
Волокна с ядерной цепочкой регистрируют исключительно степень удлинения мышц.

Следовательно, можно сказать, что фазовый ответ мышечного веретена генерируется волокнами с динамической ядерной сумкой и, в некоторой степени, волокнами со статической ядерной сумкой. Тонический ответ генерируется волокнами с ядерной цепочкой и, в некоторой степени, волокнами со статической ядерной сумкой.
Существует два типа афферентных (чувствительных) нервных волокон, иннервирующих интрафузальные волокна мышечного веретена.
Аннулоспиральные окончания передают информацию о фазовом ответе (скорости удлинения мышц). Другой тип афферентных волокон отвечает за передачу информацию о тоническом ответе (степени удлинения мышц). Теперь рассмотрим подробнее гамма-систему.
ГАММА-ЭФФЕРЕНТНАЯ СИСТЕМА
Одно из наиболее удивительных свойств нервно-мышечного веретена заключается в том, что оно является чувствительным (сенсорным) рецептором, но в то же время воспринимает и эфферентную (моторную, двигательную) информацию от центральной нервной системы. Таким образом, эти рецепторы реагируют не только на механические изменения в мышце – их реакция может быть дополнена или видоизменена вследствие влияния центральной нервной системы. Это происходит благодаря гамма-эфферентной системе.
Особые двигательные волокна, идущие к мышечному веретену, называются гамма-волокнами. Гамма-эфферентные волокна соединены с окончаниями волокон как с ядерной сумкой, так и ядерной цепочкой.
Как говорилось ранее, окончания этих волокон могут сокращаться, однако центральная их часть к сокращению не способна. Следовательно, когда по гамма-эфферентным волокнам проходит сигнал, происходит сокращение окончаний волокон с ядерной сумкой и волокон с ядерной цепочкой. Если окончания мышечных волокон сокращается, а центральная часть – нет, центральная часть подвергается растяжению.
Афферентные (чувствительные) нервные волокна интрафузальных мышечных волокон располагаются в не имеющей способности к сокращению центральной части и, будучи растянутыми, они усиливают интенсивность сократительного импульса, идущего к мышце. Яркий пример работы данной системы – рефлекс растяжения.
РЕФЛЕКС РАСТЯЖЕНИЯ
Если происходит очень быстрое растяжение мышцы, центральные части интрафузальных волокон подвергаются механическому растяжению. Это увеличивает интенсивность и скорость сигналов, посылаемых чувствительными клетками, расположенными в центральных частях интрафузальных волокон. Воспринимая высокоинтенсивные стимулы, центральная нервная система инициирует мышечное сокращение, чтобы предотвратить вероятное повреждение мышцы вследствие быстрого растяжения. Этот процесс называется рефлексом растяжения.
Кроме того, мышечное веретено может повышать тонус скелетной мускулатуры в случае повышения активности в гамма-эфферентных волокнах. По гамма-эфферентным волокнам проходят импульсы, вызывающие сокращения окончаний интрафузальных мышечных волокон, что приводит к растяжению их центральной части. Как мы уже знаем, это вызывает мышечное сокращение.
Неадекватно высокая активность гамма-волокон может привести к развитию чрезмерного мышечного напряжения. Повышение гамма-активности может быть вызвано возбуждением центральной нервной системы из-за стресса, а также вследствие развития мышечно-фасциальных триггерных точек, гипертонуса мышц, различных травм, болевого синдрома, неправильного питания и множества других факторов.
Достаточно небольшого повышения гамма-активности, чтобы значительно нарушить функционирование мышцы.
Повышение гамма-эфферентной активности является одной из важнейших причин развития различных дисфункций мышц. Мы часто слышим, что стресс и нервное напряжение может привести к соматическим проблемам, в том числе к перенапряжению мышц и нарушению их функционирования. Повышение активности гамма-системы – значимая часть этого процесса. Понимание значения и устройства гамма-эфферентной системы способствует повышению эффективности разработки стратегий терапии определённых проблем. Для снижения гамма-эфферентной активности подходят такие техники, как техника энергии мышц, терапия триггерных точек и другие методики, включающие в себя растягивание.
Уитни Лове
Читайте также: