
Эксперимент мезельсона и сталя кратко
Обновлено: 06.07.2024
Эксперимент Мезельсона-Сталя — эксперимент, проведённый двумя молекулярными биологами — Мэтью Мезельсоном и Франклином Сталем в 1958 году. Он показал, что репликация ДНК имеет полуконсервативный характер[1]. Это означает, что каждая дочерняя двойная спираль ДНК состоит из одной старой (матричной) цепи и из одной вновь синтезированной цепи В 1957 году Мезельсон, Сталь и Джером Виноград опубликовали статью о новом методе изучения молекулярного веса и парциального удельного объёма макромолекул (например, ДНК) — равновесном ультрацентрифугировании в градиенте плотности[6]. Этот метод позволяет разделять молекулы ДНК по их плотности: каждая молекула остановится в том месте градиента, где плотность раствора совпадает с её плавучей плотностью. Авторы применили этот метод для разделения молекул ДНК, содержащих изотопы азота 14N и 15N[1]. 15N не радиоактивен, а лишь тяжелее 14N. Содержащие тяжелый изотоп молекулы ДНК функциональны и могут удваиваться. Мезельсон и Сталь показали, что если вырастить несколько поколений бактерий Escherichia coli в среде, богатой 15N или 14N, затем центрифугировать их ДНК в градиенте плотности хлористого цезия, то окажется, что более тяжёлая 15N-ДНК останавливается ближе ко дну центрифужной пробирки, чем 14N-ДНК. Для того чтобы установить механизм репликации, E. coli, которые в течение нескольких поколений росли в 15N-содержащей среде (а значит их ДНК содержала только 15N) были перенесены в 14N-содержащую среду, где им было позволено разделиться только один раз. Плотность выделенной из этих клеток ДНК оказалась больше плотности ДНК бактерий, выращенных в среде, богатой 14N, но меньше плотности ДНК бактерий, выращенных в 15N среде. Это противоречило гипотезе о консервативном характере репликации ДНК, при котором ДНК разделились бы на две фракции с высокой и низкой плотностью, но не с промежуточной. Таким образом, первая гипотеза была отброшена. Однако полученный результат не исключал дисперсный механизм репликации, при котором участки материнской ДНК чередуются с участками дочерней ДНК. Чтобы выяснить, какой из оставшихся механизмов верен, была проанализирована плотность ДНК второго поколения бактерий. По гипотезе дисперсной репликации плотность ДНК второго поколения бактерий должна быть одинаковой для всех молекул и занимать промежуточное положение между плотностью ДНК клеток первого поколения и плотностью самой лёгкой ДНК. Оказалось, однако, что клетки второго поколения содержали примерно равные количества лёгких и гибридных ДНК. Этот факт позволил исключить гипотезу дисперсного механизма репликации.
Убиквитизация и деградация белков протеасомами
Убиквин протеосомный путь
важнейшее свойство ДНК — способность к самовоспроизведению. Этот процесс называется репликацией или редупликацией. Опыты М. Мезелсона и Ф. Сталя (1958) показали, что самовоспроизведение ДНК происходит полуконсервативным способом. В этих опытах несколько поколений бактерий кишечной палочки (Escherichia coli) выращивали на среде, содержащей меченый азот (15N). Через несколько поколений ДНК, входящая в состав клеток бактерий, содержала этот изотоп. Включение 15N в ДНК повысило ее плотность (тяжелая ДНК) . Клетки, содержащие тяжелую ДНК, помещали на среду, включающую 14N. После удвоения клеток, т. е. в первом поколении, вся выделенная ДНК оказалась полутяжелой (одна половина содержала 15N, а другая половина — 14N). На основании этого была создана схема воспроизведения ДНК, согласно которой в определенный момент жизни клетки цепочки ДНК расходится и на каждой материнской, как на матрице, из веществ клетки строится соответственная (комплементарная) дочерняя цепочка.

Мезельсон и Шталь эксперимент эксперимент впервые исполнен в 1958 году на Мэтью Месельсон и Франклин Stahl , который продемонстрировал пол-консервативную репликацию из дезоксирибонуклеиновой кислоты . Это означает, что во время репликации двойной спирали каждая из двух новых спиралей состоит из вновь образованной цепи и цепи исходной двойной спирали.
Резюме
Модели репликации ДНК
Для репликации ДНК были предложены три модели:
Описание эксперимента
Азот является одним из основных компонентов ДНК. Азот 14 ( 14 N) на сегодняшний день является наиболее распространенным изотопом на Земле, в отличие от азота 15 ( 15 N), который считается тяжелым. Последний не радиоактивен, просто тяжелее обычного изотопа, потому что он содержит дополнительный нейтрон .
Бактерии Escherichia coli культивируют в среде, содержащей 15 Н. Когда ДНК экстрагируют из этих клеток и затем центрифугируют в градиенте плотности соли, ДНК мигрирует в точку, где ее плотность равна плотности раствора, физиологического раствора.
После этого штаммы E. Coli, ранее культивированные в среде, содержащей 15 N, возвращают в нормальную среду, содержащую 14 N, время единичного деления клетки. Затем ДНК этих бактерий извлекается, а затем центрифугируется для сравнения результатов. Его плотность оказывается промежуточной. Если бы репликация была консервативной, появилось бы эквивалентное количество тяжелой ДНК и легкой ДНК, но не было бы гибридной ДНК промежуточной плотности, что исключает консервативную репликацию.
Однако результат может соответствовать либо дисперсионной, либо полуконсервативной репликации. В этих двух моделях ДНК состоит из эквивалентного количества тяжелого азота и легкого азота, отвечающего за промежуточную плотность.
В оставшейся части эксперимента клетки, культивируемые в среде, содержащей 15 N, затем культивируют в нормальной среде 14 N, время двух клеточных циклов. В результате получается эквивалентное количество ДНК двух плотностей. Один соответствует ДНК промежуточной плотности, обнаруженной в предыдущей части эксперимента, другой соответствует клеткам, культивируемым исключительно в среде, содержащей 14 Н. Этот результат позволяет исключить дисперсионную модель, которая дала бы промежуточную плотность ниже, чем количество клеток, которые делились только один раз, но больше, чем у клеток, культивируемых исключительно в среде, содержащей 14 N, причем количество 15 N распределяется по все большему и большему количеству нитей ДНК.
Результат этих экспериментов согласуется с полуконсервативной моделью репликации ДНК, доля легкой ДНК увеличивается с числом поколений, разделившихся в среде, содержащейся в 14 N.
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.
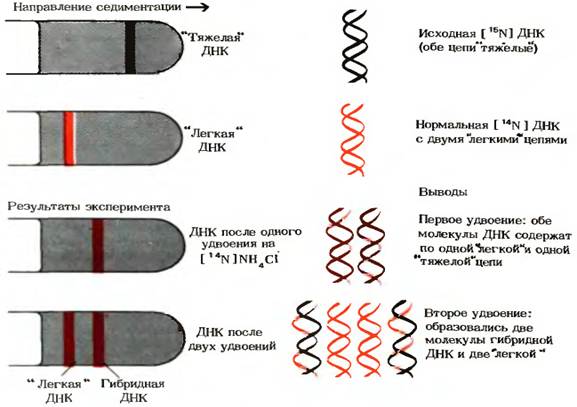
Рис. 7 Результаты эксперимента Мезельсона-Сталя
РЕПЛИКАЦИЯ ДНК
Репликация ДНК – синтез ДНК – происходит по полуконсервативному механизму. Согласно гипотезе Уотсона-Крика, каждая из цепей двойной спирали ДНК служит матрицей для репликации комплементарных дочерних цепей. При этом образуются две дочерние двухцепочечные молекулы ДНК, идентичные родительской ДНК. Каждая из этих молекул содержит одну неизмененную цепь родительской ДНК и одну вновь синтезированную цепь дочерней ДНК.

Рис. 7 Результаты эксперимента Мезельсона-Сталя
Представьте себя в 1953 году, после того, как была обнаружена структура ДНК с двойной спиралью [1]. Какие острые вопросы могут придти в вашу голову и головы других ученых?
Одним из таких больших вопросов стал вопрос касающийся репликации ДНК. Структура двойной спирали ДНК дала намек на то, как может происходить копирование [1,2]. Казалось вероятным, что две взаимодополняющие нити спирали могут разделяться во время репликации, каждая из которых служит шаблоном для построения новой, сочетающейся с ней нити.
Три модели репликации ДНК
Существовали три основные модели репликации ДНК, которые были предложены научным сообществом после открытия структуры ДНК. Эти модели показаны на диаграмме ниже:

Большинство биологов в то время, вероятно, поставили бы свои деньги на полуконсервативную модель. Эта модель имела большой смысл, учитывая структуру двойной спирали ДНК, в которой две цепи ДНК идеально предсказуемо дополняют друг друга (где каждому T (тимину) одной цепи, всегда соответствует A (аденин) другой цепи; и также каждому G (гуанину) одной цепи соответствует C (цитозин) другой цепи и так далее)$^$. Учитывая эти соображения, легко представить, что каждая нить выступает в качестве шаблона для синтеза нового партнера.
Мезельсон и Сталь разгадывают головоломку
Мэтт Мезельсон и Франклин Сталь впервые встретились летом 1954 года, через год после того, как Уотсон и Крик опубликовали свою статью о структуре ДНК. Хотя оба исследователя имели разные научные интересы, они были заинтригованы вопросом репликации ДНК и объединившись, решили попытаться определить механизм репликации [5].
Эксперимент Мезельсона и Сталя
Мезельсон и Сталь провели свои знаменитые эксперименты по репликации ДНК, используя кишечных бактерий E. coli , как модельную систему.
Они начали с выращивания E. coli в среде из питательного бульона, содержащего "тяжелый" изотоп азота — 15N (изотоп - это просто версия химического элемента, которая отличается от других версий количеством нейтронов содержащихся в ядре.) При выращивании на среде, содержащей тяжелые изотопы 15N, бактерии берут азот и используют его для синтеза новых биологических молекул, в том числе ДНК.
Мезельсон и Сталь знали, с какой частотой делились клетки E. coli, и они могли собирать небольшие образцы каждого поколения бактерий, а затем извлекать и очищать их ДНК. После чего они измеряли плотность ДНК (и, косвенно, содержание в ней изотопов 15N), используя центрифугирование в градиенте плотности.
Метод центрифугирования в градиенте плотности разделяет молекулы, такие как ДНК, на фрагменты, с помощью вращения их на высоких скоростях в присутствии другой молекулы, такой как хлорид цезия, которая создает градиент плотности от верха до низа вращающейся пробирки. Центрифугирование в градиенте плотности позволяет обнаруживать очень малые различия в плотности, такие, как, например, между ДНК помеченной тяжелым изотопом 15N и легким изотопом 14N.

,\" автор Mariana Ruiz Villareal (public domain)"]">Рисунок 2. Изображение модифицировано, источник: "Meselson-Stahl experiment diagram en," автор Mariana Ruiz Villareal (public domain)
Результаты эксперимента
Анализ молекул ДНК первых четырех поколений E. coli выявил картину расположение полос, показанную на рисунке ниже:

О чем эти результаты рассказали Мезельсону и Шталь? Давайте пройдемся по первым нескольким поколениям, которые предоставляют ключевую информацию.
Поколение 0
Поколение 1
ДНК, выделенная из бактерий первого поколения (после одного цикла репликации ДНК), также показывала одну полосу при центрифугировании. Однако эта полоса располагалась уже выше, что указывало на промежуточную плотность между ДНК с тяжелым изотопом 15N и ДНК с легким изотопом 14N.
Эта промежуточная полоса рассказала Мезельсону и Сталю, что молекулы ДНК, полученные в первом раунде репликации, представляют собой гибрид легкой и тяжелой ДНК. И этот результат соответствует дисперсионной и полуконсервативной моделям, но не соответствует консервативной модели.
Консервативная модель предсказала бы две отдельные полосы в этом поколении (полоса для тяжелой исходной молекулы ДНК и полоса для легкой, вновь созданной молекулы ДНК).
Поколение 2
Информация из второго поколения позволила Мезельсону и Сталю определить, какая из оставшихся моделей (полуконсервативная или дисперсионная) была действительно правильной.
Когда ДНК бактерий второго поколения была центрифугирована, она показала две полосы. Первая полоса находилась в той же позиции, что и промежуточная полоса из первого поколения, а вторая была выше (указывая на то, что ДНК помечена только легким изотопом 14N).

Поколения 3 и 4
В полуконсервативной модели ожидается, что каждая гибридная молекула ДНК из второго поколения будет давать одну гибридную молекулу и одну легкую молекулу в третьем поколении, тогда как каждая легкая молекула ДНК будет давать при репликации только легкие молекулы.
Таким образом, в третьем и четвертом поколениях мы ожидаем, что гибридная полоса будет становиться всё более и более узкой (поскольку она будет представлять всё меньшую и меньшую долю всей ДНК), а светлая полоса будет становиться всё шире (поскольку она будет представлять всё большую долю).
Как мы можем видеть на рисунке, Мезельсон и Шталь получили именно такую картину в своих результатах, подтвердив таким образом полуконсервативную модель репликации.
Что если дисперсионная модель была бы верна?
Дисперсионная модель предсказывает, что каждая реплицируемая молекула ДНК должна представлять собой набор фрагментов из родительской ДНК (из предыдущего поколения) и дочерней ДНК (вновь синтезированной во время репликации), что можно сравнить с лоскутным одеялом. Механистически это похоже на то, как будто старая и новая ДНК были разрезаны во время репликации, перемешаны с друг другом, а затем собраны вместе в спираль. Таким образом, каждый раунд репликации в дисперсионной модели будет производить лоскутные молекулы с тяжелыми и легкими участками. Такая лоскутная-ДНК будет содержать все большую и большую долю легкого изотопа 14N при каждой последующей репликации, поэтому плотность ДНК со временем будет постепенно уменьшаться, что приведет к появлению одной полосы (или нечеткой полосы/мазка), которая бы сдвигалась выше с каждым поколением.
Что если консервативная модель была бы верна?
Этот паттерн иллюстрирует, что при консервативной модели тяжелая ДНК никогда полностью не исчезает, но вновь синтезированная, легкая ДНК очень быстро превосходит её по количеству. Вы также можете увидеть, что при консервативной репликации никогда не образуются гибридные молекулы, которые можно видеть в двух других моделях. Это различие позволило Мезельсону и Сталю исключить консервативную модель уже после первого поколения.
Выводы
Эксперимент, проведенный Мезельсоном и Сталем, продемонстрировал, что ДНК реплицируется полуконсервативно, а это означает, что каждая цепь в молекуле ДНК служит шаблоном для синтеза новой комплементарной цепи.
Хотя Мезельсон и Шталь проводили свои эксперименты на бактериях E. coli, сегодня мы знаем, что полуконсервативная репликация ДНК является универсальным механизмом, общим для всех организмов на планете Земля. Некоторые из ваших клеток прямо сейчас полуконсервативно реплицируют свою ДНК!
Друзья, если вам понравился эта статья, и вы хотели бы видеть еще больше качественных, образовательных материалов, то пожалуйста 🙏, поддержите наш проект, поделившись 📢 этой статьей с друзьями и подписавшись на нас:
Цитируемые работы
4. Watson, J. D., and Crick, F. H. C. (1953). Genetical implications of the structure of deoxyribonucleic acid. Nature, 171, 740-741.Davis, T. H. (2004).
Читайте также: