
Экология высокогорных растений кратко
Обновлено: 30.06.2024
В анатомическом строении высокогорных растений есть ряд черт, которые отчасти способствуют защите от избыточной радиации, отчасти связаны с особенностями водного режима и некоторых сторон обмена веществ в высокогорьях: утолщение покровных тка=-ней, опушение, усиленное развитие механических тканей, придающих устойчивость к сильным ветрам, и т. д. Однако в горах довольно обычны и растения с листьями, лишенными опушения и воскового налета. С увеличением высоты местности, как правило, умень1ДШ01СЯ_р.азмеры клеток и возрастает штгность тканей, увеличивается число устьиц на единицу поверхности листа и уменьшаются их размеры; иными словами, наблюдаются изменения в сторону ксероморфоза. Особенно четко они выражены у растений, растущих на скалах; напротив, у видов, обитающих вблизи талых вод или других источников увлажнения, листья крупнее, а ксеро-морфные черты выражены гораздо слабее.
Низкие температуры и сильная освещенность способствуют образованию больших количеств антоциана, отсюда —. глубокие-, насыщенные тона окраски цветков. Сочетание крупных яркоокра-ш'енных цветков и небольших листьев при малом (иногда совсем крохотном) росте — характерная черта многих альпийских растений
Основные физиологические процессы у растений в высокогорных условиях характеризуются повышенной интертсивтюстдю. Прежде всего это относится к газообмену. На больших высотах отмечены очень высокие величины фотосинтеза (у некоторых видов до 50— 100 мг СОг на 1 г листа в 1 ч). Правда, в гумидных высокогорьях (Альпы, Гиссарский хребет) фотосинтез довольно умеренный. Тем не менее в целом по мере поднятия в горы прослеживается тенденция к увеличению фотосинтеза. Она отмечается и при сопоставлении интенсивности фотосинтеза разных видов в высотных поясах,

и у одного и того же вида, растущего (или пересаженного) н"а разной высоте. Сильнее всего эти высотные различия фотосинтеза и его температурных оптимумов заметны у видов со сравнительно слабой ассимиляцией. В соответствии с сильной освещенностью фотосинтез горных растений имеет высокий уровень светового насыщения. Так, в высокогорьях Памира он составляет 50— 60 тыс. лк (40—50% от полной освещенности), а у ряда видов световое насыщение отсутствует совсем, и фотосинтез продолжает повышаться с увеличением освещенности до максимально возможной (см. рис. 40). Снижение содержания углекислоты в воздухе, по-видимому, не отражается на фотосинтезе высокогорных растений: согласно теоретическим расчетам, уменьшение парциального давления СОг на больших высотах должно компенсироваться повышением скорости диффузии ее в лист.
Приход "солнечной радиации в горах увеличен отчасти в связи с некоторой разреженностью атмосферы, но главным образом из- за ее большой прозрачности.
В высокогорьях Памира освещенность в дневные часы составляет около 130 тыс. лк, т. е. лишь немногим меньше, чем на границе земной атмосферы. Ультрафиолетовая радиация здесь значительно сильнее, чем на равнинах: так, на высотах 2500—4000 м интенсивность радиации в области

Рис. 156. Соотношение вертикальной и широтной зональности на североамериканском континенте (схема; по Wolcott R. Н.; 1946)
Районы \Гагринспо -БзыВский Подрайоны(

Рис. 157. Схема вертикальной поясности растительности на Кавказе (Западная Грузия, Гагринско-Бзыбский район) (По Сохадзе Е. В. и Сохадзе М. Е., 1964):
/—2а — леса нижнего горного пояса с листопадным подлеском (с преобладанием; / — дуба грузинского, 2 и 2а — граба кавказского), 3 — смешанные леса с участием вечнозеленого подлеска. 4—6 — буковые леса среднего и верхнего горного пояса (разные типы), 7—10 — буково- гемнохвойные и темнохвойные леса (разные типы), 11 — высокогорные луга и скальная растительность, 12— граница известняков 290—310 нм (у границы видимого света) в десятки раз больше, чем на уровне моря (рис. 158).
Другие характерные черты высокогорных условий — пониженные температуры (в частности, дочные заморозки, действующигГиа растения в некоторых горных районах в течение большей части

158. Кривые распределения облученности в коротковолновой части спектра в полуденное время на разных высотах (по данным разных авторов):
1—уровень моря (Москва), 2—уровень моря (Вашингтон), 3 — 2320 м (Памир), 4— 1789 м (Маунт-Вильсон, Калифорния), 5 — 3860 м (Памир), 6 — 3700 м (Памир), 7 —4450 м (Памир), 8 — за пределами атмосферы
или даже всего вегетационного периода), сильные ветры, значительно укороченный _вегета- ционный сезон. На больших высотах снижено содержание углекислоты в воздухе: например, на Памире на высоте 3800 м концентрация С02 всего 0,012— 0,020%. Счт\, г С
Особо своеобразны условия в нивальном (приснежном) поясе, в непосредственной близости от массивов снега и льда, возле границы вечных снегов, тающих ледников и снежников.
„Для высокогорных растений характерен приземистый рост. Во всех "высокогорных областях земного шара преобладают низкорослые стелющиеся кустарники и кустарнички, подушковидные и ро- зеточные многолетние травы, дерновинные злаки и осоки, мхи и лишайники. Однако есть и более крупные, весьма своеобразные формы, например распространенные в высокогорьях Южной Америки (Анды) и Африки древовидные розеточные растения из родов Senecio, Espeletia, Lobelia с высоким колонновидным стеблем, несущим наверху крупную розетку мясистых, часто сильно опушенных листьев.
Характерная морфологическая черта многих высокогорных приземистых растений (кустарников, кустарничков)— значительное преобладание подземной массы по сравнению с надземной.
В анатомическом строении высокогорных растений есть ряд черт, которые отчасти способствуют защите от избыточной радиации, отчасти связаны с особенностями водного режима и некоторых сторон обмена веществ в высокогорьях: утолщение покровных тка=- ней, опушение, усиленное развитие механических тканей, придающих устойчивость к сильным ветрам, и т. д. Однако в горах довольно обычны и растения с листьями, лишенными опушения и воскового налета. С увеличением высоты местности, как правило, уменьшаются размеры клеток и возрастает плогность тканей, увеличивается число устьиц на единицу поверхности листа и уменьшаются их размеры; иными словами, наблюдаются изменения в сторону ксероморфоза. Особенно четко они выражены у растений, растущих на скалах; напротив, у видов, обитающих вблизи талых вод или других источников увлажнения, листья крупнее, а ксеро- морфные черты выражены гораздо слабее.
Низкие температуры и сильная освещенность спосббствуют образованию больших количеств антоциана, отсюда — глубокие-.
насыщенные тона окраски цветков. Сочетание крупных яркоокра- шенных цветков и небольших листьев при малом (иногда совсем крохотном) росте — характерная черта многих альпийских растений.
Основные физиологические процессы у растений в высокогорных условиях характеризуются повышенной интенсивностью. Прежде всего это относится к газообмену. На больших высотах отмечены очень высокие величины фотосинтеза (у некоторых видов до 50— 100 мг С02 на 1 г листа в 1 ч). Правда, в гумидных высокогорьях (Альпы, Гиссарский хребет) фотосинтез довольно умеренный. Тем не менее в целом по мере поднятия в горы прослеживается тенденция к увеличению фотосинтеза. Она отмечается и при сопоставлении енсивности фотосинтеза разных видов в высотных поясах,

и у одного и того же вида, растущего (или пересаженного) н“а разной высоте. Сильнее всего эти высотные различия фотосинтеза и его температурных оптимумов заметны у видов со сравнительно слабой ассимиляцией. В соответствии с сильной освещенностью фотосинтез горных растений имеет высокий уровень светового насыщения. Так, в высокогорьях Памира он составляет 50— 60 тыс. л к (40—50% от полной освещенности), а у ряда видов световое насыщение отсутствует совсем, и фотосинтез продолжает повышаться с увеличением освещенности до максимально возможной (см. рис. 40). Снижение содержания углекислоты в воздухе, по-види- мому, не отражается на фотосинтезе высокогорных растений: согласно теоретическим расчетам, уменьшение парциального давления СОг на больших высотах должно компенсироваться повышением скорости диффузии ее в лист.

Рис. 159. Содержание углеводов в листьях злака Clinelymus nutans на разной высоте над уровнем моря (по Рейнус Р.
Б — на Памире (3860 м).
Фазы развития: о, — кущение, б — цветение, в — созревание семян. Углеводы: 1 — маннозы, 2—сахароза, 3 — углеводы типа мальтозы и декстринов, 4 — крахмал, 5 — гемнцеллюлозы
Влияние низких температур сказывается на характере накопления продуктов фотосинтеза (рис. 159):двели- чйвается содержание растворимых углеводов (сахаров), играющих большую роль в холодостойкости растений (см. с. 88). Особенно возрастает ОШУ куосени, перед наступлением морозов. В высокогорьях Памира (3800 м и более) содержание растворимых углеводов наиболее велико у растений верхней части альпийского пояса, где безморозный период отсутствует (например, в листьях примулы Primula тоог- kroftiana в начале вегетации 13%, к осени — до 23%)- Не только у видов местной флоры, но и у растений, пересаженных в горы из более низких поясов, содержание сахаров возрастает (например, при пересадке из равнинных районов Франции в Альпы на высоту более 2500 м — почти в 3 раза). И наоборот, у типично высокогорных видов содержание сахаров заметно снижается после пересадки в равнинные условия. На больших высотах в растениях увеличивается концентрация важйых для тлетаболизма веществ (например, ас- корбйНовои'й других органических кислот), а также ароматических соединений и пр. Недаром ценятся питательные и вкусовые свойства высокогорных кормовых растений и молочных продуктов.
Характерная черта физиологии и биохимии высокогорных растений — повышение интенсивности окислительно-восстановительных процессов, увеличение активности участвующих в них ферментов (каталазы, пероксидазы и др.), более ниашге^ чемшу равнинных растений, температурные оптимумы их работы Во многих исследованиях отмечено усиление дыхания растений на больших высотах

7350 1900 2400 2700 3000м
/ — Primula macrocalyx, 2—Primula ruprechtii, 3—Onobtychis bieberstelntl. 4 — Sca- biosa caucasica. 3a 100% принята величина дыхания на высоте 1350 м. Температура
(рис. 160) (а следовательно, увеличение энергии, освобождающейся при распаде сложных соединений). По современным представлениям это связано с тем, что в труднвіх условиях горной среды у растений появляются такие пути усиленного потребления энергии дыхания в обмене веществ, каких нет на равнинах. Один из них — использование энергии для синтеза веществ в процессе репарации '(їїбсстановления), обеспечивающей сохранение растений при частых" повреждениях (например, от ночных заморозков). Дыхание высокогорных растений устойчиво к неблагоприятным воздействиям, что является одной из физиологических основ приспособленности растений к этим крайним условиям.
Общая экологическая черта различных физиологических процессов у высокогорных растений — снижение температурных оптиму-
-8 -6 -4 -2 8 10 20 30. 40 50 °С
J 1 1 1 1 1 1 I I 1—
Рис. 161. Температурные пределы выносливости некоторых цветковых растений на разных высотах над уровнем моря в Альпах (по Pisek А.
/—летняя минимальная температура, 2—холодостойкость, 3—область фотосинтеза, 4 — оптимум фотосинтеза, 5 — летияя максимальная температура, 6 — жаростойкость. Виды растений и высота над уровнем моря: а — Oxyria digyna (2500 м), 6 — Picea excelsa (900 м), в — Fagus sylvatica (600 м), г — Olea europaea (240 м). д — Citrus limonum (80 м)
мов, хорошо заметное при сравнении с температурными адаптация- ми растений низкогорных или равнинных местообитаний (рис. 161, ,62).
В целом сопоставление основных физиологических показателей у горных и равнинных видов (а для широкораспространенных видов — у соответствующих популяций) показывает, что на больших

Высота над уровнем моря, м
Рис. 162. Температурные оптимумы фотосинтеза пихты бальзамической — Abies balsamea, происходящей из высокогорных местообитаний на разной высоте над уровнем моря (по Fryer J. Н. a. Ledig F. Т., 1972)
высотах жизнедеятельность растений значительно интенсивнее. Очевидно, эволюция высокогорных растений шла в направлении наиболее полного использования всех возможностей короткого и холодного вегетационного периода.
Существенно изменяется при поднятии в горы сезонное развитие растений. Чем выше, тем позже тает снег весной и раньше выпадает осенью, тем короче вегетационный период,. тем~позже начинается весеннее развитие и раньше наступает осень. Весной, поднимаясь в горы, можно видеть сезонное развитие одного и того же вида в обратной последовательности (например, в низкогорном поясе — цветение, в среднем — бутонизацию, еще выше — начало вегетации и, наконец, только появление из-под снега). Напротив, осенью при подъеме в горы можно наблюдать ускоренное наступление осенних фенофаз (осеннее расцвечивание листвы, листопад, отмирание надземных частей). Сокращение вегетационного сезона с высотой хорошо иллюстрируется рис. 163.
В связи со сжатыми сроками вегетации в высокогорьях темпы сезонного развития здесь значительно ускорены. Это хорошо заметно по скорости прохождения фенологических фаз на разных высотах в горах Кавказа (табл. 19, А): например, фаза бутонизации сокращается на 5—7 дней с поднятием на 100 м (на больших высотах это сокращение меньше), что подтверждается и в экспериментальных пересадках растений на разные высоты (табл. 19, Б).

Разные виды растений неодинаково относятся к высотной зональности. Одни имеют широкий высотный диапазон и растут в разных поясах, но при этом у них сильно изменяются внешний облик и основные стороны жизнедеятельности. Примером могут служить черника и голубика в Карпатах, распространенные от низкогорного до альпийского пояса (рис. 164), или типчак в горах Кавказа и др. Однако широкий высотный диапазон не всегда связан с большой экологической пластичностью растений: так, встречающийся в Тянь-Шане на разных высотах шафран Королькова — Crocus korolkovii во всех высотных поясах вегетирует только вскоре после таяния снега, т. е. в одинаковых гидротермических условиях (хотя сроки его развития и приходятся на разное время, с января по июнь). Это пример очень узкой экологической амплитуды при широком высотном диапазоне.
Рис. 163. Схема изменения продолжительности фенологических периодов над уровнем моря с увеличением высоты (по Гамсу Г. и Вальтеру Г.).
Заштрихованная часть — период покоя, незаштрихован- ная — период вегетации. Фенологические периоды: / — ранняя весна, 2—весна, 3 — поздняя весна, 4 — первая
половина лета, 5 — вторая
половина лета, б — ранняя
осень. 7 — осень, 8 — поздняя осень
Другие виды распространены в ограниченных пределах нескольких или одного (иногда довольно узкого) высотного пояса и исчезают при переходе в соседние, сменяясь замешаюшими видами. Так, в горах на разных высотах растут близкие виды — герань холмовая — Geranium collinum и герань скальная — G. saxatile, имеющие еще внутривидовые высотные формы с морфологическими и биохимическими различиями и узкой экологической амплитудой. Иногда вид выходит за пределы определенной высотной зоны, но при этом поселяется уже в совсем иных
Рис. 164. Голубика — Vaccinium uliginosum в разных высотных поясах Восточных Карпат.
А — ннзкогорный пояс (800 м над уровнем моря), поляна в еловом лесу; Б — альпийский пояс (1600 м над уровнем моря)
1 На высоты, указанные в графах таблицы.
Таким образом, отношения между высотной и экологической амплитудами растений сложны и многообразны.

Рис. 165. Различие угла падения солнечных лучей на склоны южной и северной экспозиции
Экспозиция и крутизна склона. Наряду с высотой над уровнем моря условия для жизни растений в горах в большой мере определяются экспозицией и крутизной склонов. Известно, что на склонах южной экспозиции угол падения солнечных лучей ближе к прямому чем на горизонтальной поверхности (за исключением экваториальных областей) (рис. 165).
В связи с неодинаковыми условиями на склонах разной экспозиции заметно различается состав растительности, облик и состояние растений. Известно, что на южных склонах граница древесной растительности поднимается гораздо выше, чем на северных. В целом границы всех зон сдвигаются кверху, а в составе растительных группировок преобладают более южные и теплолюбивые элементы.
Различны в зависимости от экспозиции и морфофизиологические особенности растений одного и того же вида. Так, туркестанский можжевельник арча (Juniperus turkestanica) в субальпийском поя- .се гор Киргизии обычно имеет стланиковую форму (в возрасте

Рис. 166. Температура поверхности почвы на горных склонах разной экспозиции. Западный Тянь-Шань, 3000 м над уровнем моря (по Культиасову И. М., 1955). I — южный склон, II — северный склон.
Температура: 1 — максимальная, 2 — среднесуточная, 3 — минимальная


Рис. 167. Распределение листоватых лишайников на гранитном валуне (по Ниценко А. А., 1951).
А — на южной стороне, Б — на северной стороне
а водному режиму — более гигрофильные (меньшая транспирация, большая оводненность листьев и т. д.). Однако в более напряженных условиях водоснабжения и температуры могут иметь место иные соотношения: так, растения злаково-разнотравных лугов Кавказа больше транспирируют на северных склонах.
Влияние различной экспозиции отражается на составе растительности не только в случае крупных элементов рельефа; оно хорошо заметно и на небольших холмах, повышениях, валунах (рис. 167).
Влияние крутизны склона на условия жизни растений сказывается главным образом через особенности почвенной среды, водного и температурного режима. Сильный сток воды и смыв почвы с крутых склонов создают трудные условия для поселения растений. Преимущество здесь имеют виды литофильного характера с глубокой и цепкой корневой системой, экономно расходующие воду. На склонах с более мягким грунтом (например, крутые стенки глубоких оврагов) хорошо закрепляются растения-пионеры с поверхностной и разветвленной корневой системой (мать-и-мачеха и др.).
В горных странах с весьма сложным рельефом создается очень сложное переплетение влияний — широтных факторов, вертикальной зональности, различий экспозиции склонов, их крутизны, степени изрезанности, а также особенностей почвенно-грунтовых условий, водного режима и т. д. Поэтому картина распределения экологических факторов и растительности может быть сильно усложнена. Так, в замкнутых котловинах, даже на небольшой высоте, скапливается более холодный воздух, чем в вышележащих поясах; в таких случаях возможны инверсии (обращения) зональности. Например, очень наглядны инверсии в предгорьях Низких Татр в Словакии со сложным рельефом в области карстовых явлений: высокогорные виды спускаются здесь очень низко по холодным и влажным ущельям, а многие теплолюбивые виды низин, напротив, поднимаются по хорошо освещенным и прогретым известняковым хребтам выше своего обычного высотного ареала.
А горная экосистема Это совокупность биотических (живые организмы) и абиотических (климат, почва, вода) факторов, которые развиваются в горном рельефе. В горной местности фактор высоты имеет решающее значение, поскольку он создает градиент условий окружающей среды, особенно температуры.
При подъеме в высокие горы температура падает, и это влияет на присутствующую растительность и фауну. Таким образом, существует предел высоты, выше которого деревья больше не высаживаются, что, в свою очередь, зависит от широты.
С другой стороны, горы - это естественные препятствия, вызывающие усиление ветра и конденсацию влаги, вызывающую дожди. Точно так же ориентация склонов влияет на падение солнечной радиации.
Все эти элементы влияют на ряд экосистем, сформировавшихся в горах, от лесов и джунглей до альпийской тундры. В высокогорье последовательность экосистем зависит от высоты и аналогична тем, которые возникают из-за широтных колебаний.
В тропиках с высотным градиентом наиболее распространенными горными экосистемами являются сезонные леса в предгорьях, за которыми следуют облачные леса на больших высотах. Впоследствии за пределами деревьев появляются холодные кустарники и луга, холодные пустыни и, наконец, вечный снег.
И в умеренном, и в холодном поясе высотная последовательность идет от горных экосистем умеренного лиственного леса, субальпийского хвойного леса, альпийской тундры и вечного снега.
Характеристики горной экосистемы
Гора как физическая опора горных экосистем определяет ряд элементов, которые влияют на их характеристики и их распределение.
Высота и температура
Когда вы поднимаетесь на высокую гору, температура окружающей среды снижается, что называется вертикальным температурным градиентом. В горах умеренного пояса температура понижается на 1 ° C на каждые 155 м высоты, а в тропической зоне, с более высокой солнечной радиацией, на 1 ° C на каждые 180 м высоты.
На эти различия в температурном градиенте также влияет тот факт, что атмосфера в тропиках толще, чем в умеренных и холодных широтах. Это оказывает решающее влияние на распределение горных экосистем по высотному градиенту.
Климатические условия, создаваемые высотой, определяют, что на нижнем и среднем уровнях горы есть леса, а на более высоких - редкая, травянистая или кустарниковая растительность.
Ограничение по дереву
Снижение температуры и доступность воды определяют предел высоты, выше которого деревья не развиваются в горах. Следовательно, оттуда присутствующие экосистемы будут кустарниками или лугами.
Этот предел ниже по мере увеличения широты, то есть дальше на север или юг. В тропических зонах этот предел достигается на высоте от 3500 до 4000 метров над уровнем моря.
Орографический дождь
Гора определенной высоты представляет собой физическое препятствие для циркуляции воздушных потоков, которые при столкновении с ней поднимаются. Эти поверхностные течения теплые и влажные, особенно если они движутся над океаническими массами.
Когда они сталкиваются с горами и поднимаются, воздушные массы охлаждаются, а влага конденсируется в облака и осадки.
Влияние ориентации склона
В горных экосистемах присутствует эффект наклона, то есть роль, которую играет ориентация склонов по отношению к Солнцу. Таким образом, грани горы получают солнечное излучение в разное время суток, что влияет на тип присутствующей растительности.
Точно так же существуют различия во влажности между двумя склонами горного хребта, параллельного морскому побережью. Это связано с тем, что влажность с моря, переносимая ветрами, остается на наветренном склоне (обращенном к ветру).
В то время как подветренный склон (противоположная сторона) принимает ветры, преодолевшие гору и потерявшие большую часть влажности.
Солнечная радиация
В высокогорье атмосфера менее плотная, что способствует большему падению солнечного излучения, особенно ультрафиолетовых лучей. Это излучение оказывает негативное воздействие на живые ткани, поэтому растениям и животным необходимы стратегии для его предотвращения.
Многие высокогорные растения имеют мелкие твердые листья, обильное опушение или особые пигменты.
Эффект гравитации
Характерным действием гор является фактор силы тяжести, так как растительность должна компенсировать силу тяжести на крутых склонах. Точно так же сила тяжести влияет на сток дождевой воды, инфильтрацию и доступность воды, что определяет тип присутствующей растительности.
Последовательность горной экосистемы
В высоких горах экосистемы меняются от основания до вершины, в основном в зависимости от температуры и влажности. Нечто похожее на то, что происходит между экватором Земли и полюсами Земли, где возникает широтное изменение растительности.
Высокая умеренная и холодная гора
В горах умеренного и холодного пояса лиственные леса умеренного пояса встречаются в нижних частях, подобные тем, что встречаются в умеренных широтах. За более высокими высотами следуют субальпийские хвойные леса, похожие на бореальную тайгу в субполярных широтах.
Наконец, на возвышенностях появляется альпийская тундра, похожая на арктическую тундру, а затем - зону вечного снега.
Высокая тропическая гора
В случае с тропическими горами последовательность горных экосистем включает ряд типов тропических лесов в нижних частях. Позже, на большей высоте, луга, похожие на альпийскую тундру, и, наконец, зона вечного снега.
Горы мексиканской Сьерра-Мадре - хороший пример соответствия между последовательностью горных и широтных экосистем. Это потому, что они являются переходными регионами между умеренным и тропическим поясами.
В них расположены экосистемы горных тропических лесов в нижних частях, а затем смешанные леса из умеренных и тропических покрытосеменных растений вместе с хвойными. Выше хвойные леса, затем альпийская тундра и, наконец, вечный снег.
Флора
Флора в горных экосистемах очень разнообразна, в зависимости от высоты, на которой они развиваются.
Тропические горные экосистемы
Лиственные или полуолиственные леса встречаются в тропических Андах в предгорьях и на нижних склонах. Затем при восхождении развиваются влажные леса и даже туманные леса, а на больших высотах - болотистые или холодные луга.
В этих тропических горах есть экосистемы горных джунглей с разнообразными слоями и многочисленными эпифитами и альпинистами. Mimosaceae бобовые, виды рода Фикус, lauraceae, пальмы, орхидеи, раковые и бромелиевые.
Во влажных горных лесах растут деревья высотой более 40 м, такие как синица (Альбиция карбонария) и ребенок или ложка (Gyranthera caribensis). В то время как в болотах много трав и кустарников компоста, вересковых и бобовых.
Горные экосистемы умеренного пояса
В горах умеренного пояса наблюдается переход от лиственного леса умеренного пояса к хвойному лесу, а затем к альпийской тундре. Вот такие умеренные покрытосеменные, как дуб (Quercus robur), бук (Fagus sylvatica) и березы (Betula виды).
А также хвойные породы вроде сосны (Pinus spp.) и лиственницы (Ларикс децидуа). В то время как альпийская тундра изобилует розоцветными, травами, а также мхами и лишайниками.
Горные экосистемы приполярных широт
На нижних склонах развиваются хвойные леса и смешанные леса между хвойными и покрытосеменными. В высокогорных районах развивается альпийская тундра с редкой травянистой и кустарниковой растительностью.
Фауна
Фауна также меняется с высотой, в основном из-за температуры и типа растительности. В джунглях или лесах в нижней и средней частях обычно наблюдается большее разнообразие, чем в горных экосистемах на больших высотах.
Тропические горные экосистемы
В тропических горах существует большое биологическое разнообразие, с многочисленными видами птиц и насекомых, а также рептилий и мелких млекопитающих. Точно так же обитают представители семейства кошачьих, среди которых выделяется этот род. Пантера с ягуаромPanthera onca) в Америке леопард (Panthera pardus) в Африке и Азии, а тигр (Panthera tigris) в Азии.
В горных лесах центральной Африки водится вид горной гориллы (Горилла beringei beringei). С другой стороны, в горах Анд, в Южной Америке очковый медведь (Tremarctos ornatus).
Умеренные и холодные горные экосистемы
В горных экосистемах этих территорий обитает бурый медведь (Ursus arctos), черный медведь (Ursus americanus) и кабана (Sнам скрофа). Как лиса (Vязвы vulpes), волк (Canis lupus) и разные виды оленей.
Также есть разные виды птиц, например, тетерев (Тетраус урогаллус) в Пикос-де-Эуропа и бородатый стервятник (Gypaetus barbatus) в Пиренеях. В смешанных лесах северного Китая большая панда (Ailuropoda melanoleuca), символический вид сохранения.
Примерыгорная экосистема
Облачный лес Ранчо Гранде (Венесуэла)
Этот облачный тропический лес расположен в горном хребте Коста в северо-центральном регионе Венесуэлы, на высоте от 800 до 2500 метров над уровнем моря. Для него характерен густой подлесок из крупных злаков и кустарников, а также два древесных слоя.
Первый слой составляют небольшие деревья и пальмы, за ним следуют деревья до 40 метров в высоту. Затем эти деревья изобилуют плетистыми и бигнониевыми, а также эпифитными орхидеями и бромелиями.
Погода
Фауна
Вы можете найти такие виды, как ягуар (Panthera onca), пекари с воротником (Таяссу пекари), обезьяна арагуато (Аллуата старческая) и ядовитые змеи (Ботроп атрокс, B. venezuelensis).
В этом районе он считается одним из самых разнообразных в мире по количеству птиц, выделяя турпиаль (Желтушная желтуха), коното (Псароколий декуманский) и сорокуа (Трогон воротничок). Такое высокое разнообразие объясняется наличием естественного прохода, через который проходят миграции птиц с севера на юг Америки, известного как Пасо Портачуэло.
Тропическая пустошь
Это растительное образование в высоких тропических горах Анд в Эквадоре, Колумбии и Венесуэле на высоте более 3500 метров над уровнем моря до предела вечного снега. Он состоит из розовых трав и мягких листьев с плюшевыми листьями, а также невысоких кустов с жесткими листьями.
Наиболее характерным семейством растений является соединение (Asteraceae) с многочисленными эндемичными родами, такими как Эспелетия (frailejones).
Погода
Это холодный климат высоких тропических гор, с низкими температурами и морозами ночью и высокой солнечной радиацией днем. Дожди в páramo обильные, но вода не всегда доступна, потому что она замерзла до основания, а уровень эвапотранспирации высок.
Фауна
Представлены различные виды насекомых, рептилий и птиц, среди которых характерен андский кондор (Vultur gryphus). Также можно получить Очкового или Фронтинного медведя (Tremarctos ornatus) и андский олень матакан (Mazama bricenii).
Особенности , зоны высокогорных растений
Схема, показывающая соответствие между последовательными
вертикальными и горизонтальными растительными зонами:
1 — тропическая, зона (зона тропических лесов); 2 — умеренная зона (зона лиственных и хвойных лесов); 3 – альпийская зона (зона травянистой растительности, мхов и лишайников); 4 - полярная зона (зона снегов и льдов)
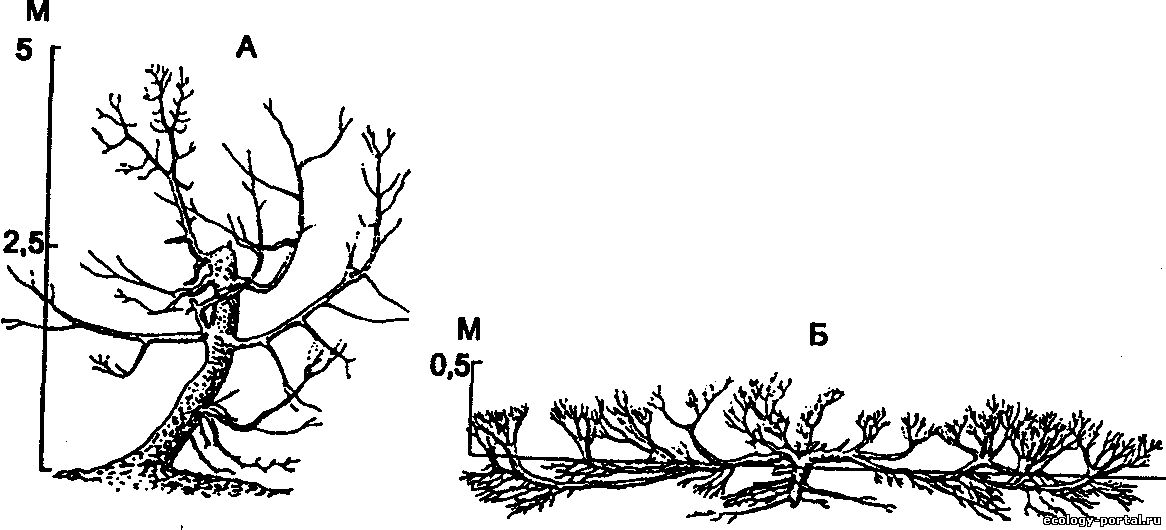
Можжевельник туркестанский - на склонах хребта
Терекей-Алатау (по И. Г. Серебрякову, 1955):
А – древовидная форма (лугово-лесной пояс, 2900 м над уровнем моря); Б – стланник (субальпийский пояс, 3200 м над уровнем моря)
Читайте также: