
Эффект положения генов это в биологии кратко
Обновлено: 07.07.2024
Комплиментарность - это способ взаимодействия неаллельных генов, при котором они взаимно дополняют друг друга,(несколько генов из различных аллелей вместе формируют признак).
Большинство сложных признаков, являясь результатом целой цепи биохимических и структурных преобразований, требует участия многих генов, влияющих на разные звенья этого процесса. Отсутствие нормального первичного продукта, хотя бы одного из них, не позволяет сформировать нормальный сложный признак. Так как часто нормальный первичный продукт гена является результатом экспрессии его доминантного аллеля, сложный признак формируется при наличии в генотипе доминантных аллелей всех взаимодействующих генов.
Примеры : тестикулярная феминизация, или синдром Морриса. Процесс формирования половой принадлежности организма у человека. Развитие признаков пола у человека, так же как у большинства животных, определяется сочетанием гетерохромосом в его кариотипе. Наличие Х- и Y-хромосом обусловливает возможность формирования мужского пола, а двух Х-хромосом —женского. Однако, для развития организма мужского пола не достаточно присутствия одного лишь Y-сцепленного гена, который определяет дифференцировку половых желез по мужскому типу и синтез гормона тестостерона. Для этого необходим также продукт другого гена — белок-рецептор, обеспечивающий проникновение гормона в клетки тканей-мишеней. За синтез такого белка отвечает особый ген, расположенный в Х-хромосоме. Его мутация, нарушающая образование нормального белка-рецептора, делает ткани-мишени невосприимчивыми к гормону, направляющему их развитие по мужскому типу. Не использовав такую возможность на определенном этапе онтогенеза, организм осуществляет развитие по женскому типу. В результате появляется особь с кариотипом XY, но внешне более сходная с женщиной. Такие субъекты не способны иметь потомство, их половые железы недоразвиты, а их выводные протоки часто формируются по женскому типу. Вторичные половые признаки также характерны для женского пола. Достижение конечного результата при формировании у человека признаков мужского пола является следствием взаимодополняющего действия нескольких генов, определяющих возможность синтеза мужского полового гормона и белка-рецептора.
Формирование слуха у человека зависит от двух неаллельных генов, один из которых отвечает за развитие улитки, а другой – за развитие слухового нерва. Если имеются два неаллельных доминатных гена (A_B_) то развивается нормальный (слышащий) организм, если по одной или двум парам генов организм является рецессивной гомозиготой, то он страдает глухотой 1 типа, 2 типа или и 1,и 2 типов одновременно.
Пигментация волос зависит от черного и от красного пигментов.
M bw – среднее кол-во черного пигмента
M bk - малое кол-во черного пигмента
M bd - большое кол-во черного пигмента
R1 – не вырабатывается красный пигмент
R2 - вырабатывается красный пигмент
Различные комбинации дают весь спектр.
Эпистаз – это способ взаимодействия неаллельных генов, при котором одна пара генов может подавлять активность другой пары генов.
Эпистатический ген – ген, который подавляет.
Гиостатический ген – ген, который подавляется.
Доминантный эпистаз – взаимодействие, при котором проявляется подавляющее действие в гомо и гетерозиготном состоянии.
Рецессивный эпистаз - проявляется подавляющее действие только в гомозиготном состоянии.
Пример: Бомбейский феномен, рецессивный эпистаз.
При изучении генотипов было обнаружено, что на формирование группы крови по системе AB0 кроме известных аллелей (I A , I B , I 0 ), влияет эпистатический ген H, который подавляет действие других аллелей в рецессивном гомозиготном состоянии.
Эффект положения генов – это вид взаимодействия неаллельных генов, при которомпроявление действия гена зависит от качества рядом расположенных генов. Является частным случаем эпистаза.
Особый вид представляет взаимодействие, обусловленное местом положения гена в системе генотипа,— эффект положения. Непосредственное окружение, в котором находится ген, может сказываться на характере его экспрессии. Изменение активности гена, наблюдаемое при хромосомных перестройках, нередко связано с перемещением его в другую группу сцепления при транслокациях или изменением его положения в своей хромосоме при инверсиях.
Явление открыто А. Стёртевантом в 1925 . Различают два типа эффекта положения гена — стабильный и нестабильный. Стабильный эффект положения гена наблюдается при перемещении гена между эухроматиновыми участками хромосом. Возможный механизм этого явления — вовлечение перемещённого гена в систему регуляции других генов. Нестабильный эффект положения гена обычно наблюдается при перемещении гена из эухроматина в гетерохроматин. В случае перемещения гена из эухроматина в гетерохроматин в некоторых клетках происходит инактивация транскрипции гена, причём инактивированное состояние гена наследуется в клеточных поколениях. Вследствие такой инактивации появляется мозаицизм признака в соматических тканях.
Пример : резус. 3 пара генов C, D, E отвечают за резус-фактор - за образование антигенов C, D, E. Если в генотипе в одной из гомологичных хромосом локализованы доминантные аллели, а в другой – рецессивные, то ген C подавляет действие гена E, поэтому на мембране – присутствуют D и C, а антигена E – нет. Если в одной из гомологичных хромосом локализованы аллели C, d,e, а в другой – c, d, E, то ген C не подавляет действие гена E, поэтому на мембране – присутствуют C, D и E.
Полимерия – способ взаимодействия неаллельных генов, при котором несколько генов влияют на развитие одного признака. Каждый ген определяет развитие определенной дозы признака: доминантный – большую, рецессивный – меньшую дозу признака. Взаимодействие генов проявляется в сумме их эффектов. Совместное действие полигенов обусловливает различную экспрессивность — степень выраженности признака, зависящую от дозы соответствующих аллелей.
В основе появления в геноме таких генов, лежит их дупликация или амплификация. Это позволяет увеличить синтез соответствующего продукта в клетках организма.
Все гены обозначаются одной буквой (так кат влияют на формирование одного признака), рядом с буквой ставится цифровой индекс, указывающий на расположение гена в хромосоме.
Полимерные факторы были открыты шведским генетиком Нильсоном-Эле при изучении наследования окраски семян у пшеницы.
Пример: определение интенсивности окраски кожных покровов, зависящей от уровня отложения в клетках пигмента меланина. В геноме человека имеется четыре гена, отвечающих за этот признак. В генотипе все они представлены в двойной дозе. Наличие в генотипе восьми доминантных аллелей в системе полигенов, определяющих цвет кожи, обусловливает максимальную ее пигментацию, наблюдаемую у африканских негров (P1P1P2P2P3P3P4P4). Полное отсутствие доминантных аллелей у. рецессивных гомозигот (р1р2р2р3р3р4р4) проявляется в виде минимальной пигментации у европеоидов. Большее или меньшее количество доминантных аллелей, колеблющееся от 8 до 0, обеспечивает разную интенсивность окраски кожи.
Полимерное взаимодействие генов лежит в основе определения главным образом количественных признаков (рост, масса организма, возможно, интеллект).
Модифицирующее действие генов — усиление или ослабление действия главных генов неаллельными им генами-модификаторами, которые в первом случае называются интенсификаторами, а во втором — супрессорами (ингибиторами). Один и тот же ген может быть главным в отношении контролирования развития одного признака и модификатором в отношении развития др. признака. При изучении явления взаимодействия были открыты гены, которые сами по себе не определяют какую-либо качественную реакцию или признак, а лишь усиливают или ослабляют проявление действия основного гена. Это гены-модификаторы, а их действие - модифицирующее.
Одни из генов-модификаторов могут усиливать эффект основного гена, другие ослаблять. Это явление пенетрантности. Она выражается долей особей, у которых проявляется исследуемый признак среди особей одинакового генотипа, по контролируемому этот признак гену. Пенетрантность определяет частоту соответствия фенотипа определенному генотипу.
Плейотропия – явление зависимости нескольких признаков от одного гена.
Первичная – ген одновременно проявляет свое множественное действие, так как измененный белок мутантного аллеля взаимодействует с цитоплазмой различных клеточных систем или изменяет свойства мембран в клетках нескольких генов. Пример: синдром Марфана обусловлен аутосомным доминантным геном коллагена. Поэтому одновременно поражается скелет, сосуды, хрусталик глаза.
Вторичная плейотропия типичное первичное фенотипическое проявление вслед за которым развиваются ступенчатые вторичные процессы. Пример, при серповидно-клеточной анемии, существует одно первичное место действия мутантного гена — гемоглобин в эритроцитах, а все остальные наблюдаемые при ней симптомы, такие, как нарушение умственной и физической деятельности, сердечная недостаточность, местные нарушения кровообращения, увеличение и фиброз селезенки и многие другие, возникают как следствие аномального гемоглобина.
Для анализа генетического контроля необходимо, кроме того, знать место первичного действия данного гена, т.е. следует различать случаи относительной, или зависимой, плейотропии от прямой, или истинной, плейотропии. В случае относительной плейотропии, как, При прямой плейотропии все разнообразные дефекты, возникающие в различных тканях или органах, вызываются непосредственным действием одного и того же гена именно в этих разных местах.
пенетрантность и экспрессивность
Фенотипическое проявление информации, заключенной в генотипе, характеризуется показателями пенетрантности и экспрессивности.
Пенетрантностъ отражает частоту проявления определенного аллеля в признак. Она соответствует проценту особей, у которых доминантный аллель гена проявился в признак, по отношению ко всем носителям этого аллеля. Неполная пенетрантность доминантного аллеля гена может быть обусловлена системой генотипа, в которой функционирует данный аллель и которая является своеобразной средой для него. Взаимодействие неаллельных генов в процессе формирования признака может привести при определенном сочетании их аллелей к непроявлению доминантного аллеля одного из них.
Наличие в генотипе одного из генов в рецессивном гомозиготном состоянии не дает возможности проявиться доминантному аллелю другого гена (альбинизм, бомбейский феномен). Известны также случаи, когда фенотипическому проявлению определенного аллеля препятствуют факторы окружающей организм среды.
В медицинской генетике если у всех носителей патологичного гена наблюдается его клиническое (фенотипическое) проявление, то можно говорить о полной пенетрантности (ген хореи Гентингтонна, синдром Марфана, ахондроплазия). Если действие мутантного гена проявляется не у всех его носителей – неполная пенетрантность. В этом случае носитель патологичного гена может быть клинически здоров, а в родословной наблюдается пропускание поколения.
Пенетрантность зависит от 1) взаимодействия генов (эпистаз, эфф. положения), 2) от особенностей генотипа- гены признаков зависящих от пола, 3) от факторов среды.
Экспрессивность также является показателем, характеризующим фенотипическое проявление наследственной информации. Она характеризует степень выраженности признака и, с одной стороны, зависит от дозы соответствующего аллеля гена при моногенном наследовании или от суммарной дозы доминантных аллелей генов при полигенном наследовании, а с другой — от факторов среды.
Эти отличия в основном состоят в следующем.
В мейозе профаза I более продолжительна. В ней происходит конъюгация (соединение гомологичных хромосом) и обмен генетической информацией. В анафазе I центромеры, скрепляющие хроматиды, не делятся, а к полюсам отходит одна из гомологмейоза
Митоз и его фазы
митоза и ичных хромосом. Интерфаза перед вторым делением очень короткая, в ней ДНК не синтезируется. Клетки (галиты), образующиеся в результате двух мейотических делений, содержат гаплоидный (одинарный) набор хромосом. Диплоидность восстанавливается при слиянии двух клеток — материнской и отцовской. Оплодотворенную яйцеклетку называют зиготой.
Митоз, или непрямое деление, наиболее широко распространен в природе. Митоз лежит в основе деления всех неполовых клеток (эпителиальных, мышечных, нервных, костных и др.). Митоз состоит из четырех последовательных фаз (см. далее таблицу). Благодаря митозу обеспечивается равномерное распределение генетической информации родительской клетки между дочерними. Период жизни клетки между двумя митозами называют интерфазой. Она в десятки раз продолжительнее митоза. В ней совершается ряд очень важных процессов, предшествующих делению клетки: синтезируются молекулы АТФ и белков, удваивается каждая хромосома, образуя две сестринские хроматиды, скрепленные общей центромерой, увеличивается число основных органоидов цитоплазмы.
В профазе спиралируются и вследствие этого утолщаются хромосомы, состоящие из двух сестринских хроматид, удерживаемых вместе центромерой. К концу профазы ядерная мембрана и ядрышки исчезают и хромосомы рассредоточиваются по всей клетке, центриоли отходят к полюсам и образуют веретено деления. В метафазе происходит дальнейшая спирализация хромосом. В эту фазу они наиболее хорошо видны. Их центромеры располагаются по экватору. К ним прикрепляются нити веретена деления.
В анафазе центромеры делятся, сестринские хроматиды отделяются друг от друга и за счет сокращения нитей веретена отходят к противоположным полюсам клетки.
В телофазе цитоплазма делится, хромосомы раскручиваются, вновь образуются ядрышки и ядерные мембраны. В животных клетках цитоплазма перешнуровывается, в растительных — в центре материнской клетки образуется перегородка. Так из одной исходной клетки (материнской) образуются две новые дочерние.
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа). Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты.
ДНК-полимераза может присоединять нуклеотид только к 3'-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3'-конца к 5'-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по-разному и в противоположных направлениях. На цепи 3'–5' синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей. На цепи 5'–3' — прерывисто, фрагментами (фрагменты Оказаки), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстающей).
Репликация протекает сходно у прокариот и эукариот. Скорость синтеза ДНК у прокариот на порядок выше (1000 нуклеотидов в секунду), чем у эукариот (100 нуклеотидов в секунду). Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК. Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов. Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой. Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Пример – наследование белка Rh-фактора (резус-фактора). У 85% европейцев резус-фактор имеется (Rh+), у 15% – его нет (Rh-). Определяется резус-фактор тремя доминантными генами (С, D, E), расположенными в хромосоме рядом друг с другом.
Два человека с одинаковым генотипом CcDDEe будут иметь разные фенотипы в зависимости от варианта расположения аллельных генов в паре гомологичных хромосом: в варианте А – много антигена Е, но мало антигена С; в варианте В – мало антигена Е, но много антигена С.
Смотреть что такое "ЭФФЕКТ ПОЛОЖЕНИЯ ГЕНА" в других словарях:
эффект положения гена — geno padėties efektas statusas T sritis augalininkystė apibrėžtis Geno veikimo pakitimas dėl jo padėties chromosomoje pakeitimo kitų genų atžvilgiu. atitikmenys: angl. gene position effect rus. эффект положения гена … Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
Эффект положения гена — влияние расположения Генов в хромосоме на. проявление их активности. Явление открыто американским генетиком А. Стёртевантом в 1925. Наблюдается при структурных перестройках хромосом (транслокациях), в результате которых гены активных зон… … Большая советская энциклопедия
эффект положения гена — зависимость фенотипического проявления гена от его локализации в хромосоме … Большой медицинский словарь
эффект положения — position effect эффект положения. Изменение экспрессии гена в результате изменения его локализации на хромосоме (в своей группе сцепления), возникающего при хромосомных перестройках и при рекомбинации; различают стабильный Молекулярная биология и генетика. Толковый словарь.
эффект Дубинина — Dubinin effect эффект Дубинина. Вариант эффекта положения
, связанный не с удвоением гена (как в случае с геном Bar у дрозофилы), а с переносом определенного гена в новое генетическое “окружение”; Э.Д. впервые… … Молекулярная биология и генетика. Толковый словарь.
ПОЛОЖЕНИЯ ЭФФЕКТ — Явление изменения действия гена в зависимости от его положения в системе целой хромосомы … Термины и определения, используемые в селекции, генетике и воспроизводстве сельскохозяйственных животных
Выключение гена — Выключение гена, сайленсинг генов, англ. gene silencing это общий термин, описывающий эпигенетический процесс регуляции генов. При этом последовательность нуклеотидов не изменяется, а лишь прекращается экспрессия соответствующего гена. Для… … Википедия
Сайленсинг гена — Выключение гена, Сайленсинг генов, Gene silencing это общий термин, описывающий эпигенетический процесс регуляции генов. При этом последовательность нуклеотидов не изменяется, а лишь прекращается экспрессия соответствующего гена. Сайленсинг генов … Википедия
Вы уже знаете о том, что гены могут взаимодействовать друг с другом по типу полного и неполного доминирования. Однако, в генетике встречается масса других примеров взаимодействия генов. В этой статье мы затронем те, которые ранее не обсуждались.
Кодоминирование
Кодоминирование - взаимодействия аллельных генов, при котором в гетерозиготном состоянии могут оказаться два доминантных гена одновременно, при этом каждый ген отвечает за свой признак.
Наиболее распространенным примером кодоминирования является наследование групп крови у человека.

Решим пару задач, которые укрепят понимание темы.
Пример решения задачи №1
"Родители имеют II и III группы крови, гетерозиготны. Какие группы крови можно ожидать у их детей?"
Гетерозиготный генотип матери - I A i 0 и генотип отца - I B i 0 . Составим схему решения для такого случая.

Итак, в результате такого брака может получиться ребенок с любой группой крови, в чем мы убедились.
Пример решения задачи №2
"Дигетерозиготная по B (III) группе и положительному резус-фактору вступила в брак с таким же мужчиной. Какое расщепление по фенотипу можно ожидать у детей?"
Сходу понятно, что гетерозиготы по III (B) группе крови будут записаны I B i 0 . Резус-фактор для нас новое понятие - это белок, находящийся на поверхности эритроцита (тогда говорят, что резус-фактор положителен), или отсутствующий (тогда у человека резус-фактор считается отрицательным). Генотипы записываются так:
- Резус-фактор положителен: Rh + Rh + , Rh + rh -
- Резус-фактор отрицателен: rh - rh -
В данной задаче сказано, что "дигетерозиготна по . и положительному резус-фактору" - значит, резус фактор будет записывать в генотипе - Rh + rh - .

Обратите внимание, что ошибкой является записать рецессивный ген перед доминантным. За такое могут снять балл на экзамене: aA, bB, i 0 I A , rh - Rh + . Правильный вариант записи: Aa, Bb, I A i 0 , Rh + rh - .
- 9 потомков : положительный резус-фактор, III (B) группа крови
- 3 потомка : положительный резус-фактор, I (0) группа крови
- 3 потомка : отрицательный резус-фактор, III (B) группа крови
- 1 потомок : отрицательный резус-фактор, I (0) группа крови
Расщепление по фенотипу в данном случае получилось: 9:3:3:1. Здесь проявляется III закон Менделя - закон независимого наследования, так как гены, отвечающие за группу крови и резус-фактор, находятся в разных хромосомах.
Комплементарность
Комплементарность - тип взаимодействия неаллельных генов, при котором развитие признака определяется не одной, а двумя или более парами неаллельных генов, располагающихся в разных хромосомах.
Неаллельные гены - это гены, расположенные в разных локусах хромосом, которые отвечают за разные признаки. В генетике случается такое, что один неаллельный ген может влиять на другой (ген a подавляет действие гена B). В этом разделе статьи мы подробно разберемся с подобным взаимодействием и рассмотрим задачи, которые могут встретиться.
Таким образом, развитие признака определяется именно сочетанием генов друг с другом. Здесь логичнее подчеркнуть совместное действие генов, нежели чем сказать, что доминантный ген подавляет рецессивный - при комплементарности это не совсем так.
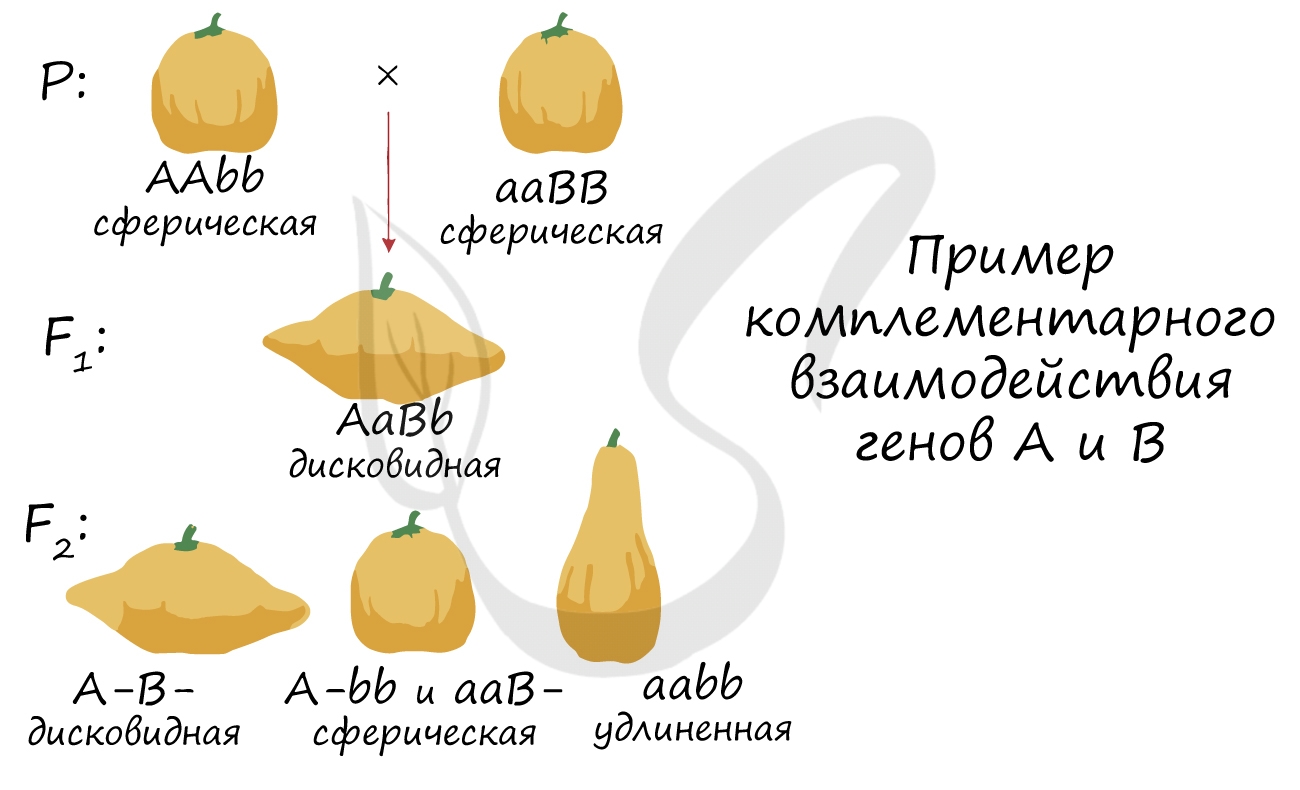
В каждой задаче свой случай комплементарного взаимодействия генов. Чтобы успешно их решать, надо помнить, что такое явление, как комплементарность, в принципе, возможно, и быть внимательным при написании генотипов особей и их гамет.
Пример решения задачи №3
Наследование слуха у человека определяется двумя доминантными генами из разных аллельных пар, один из которых детерминирует развитие слухового нерва, а другой – улитки. Определить вероятность рождения глухих детей, если оба родителя глухие, но по разным генетическим причинам (у одного отсутствует слуховой нерв, у другого улитка). По генотипу оба родителя являются дигомозиготными.

Здесь проявляется I закон Менделя - закон единообразия гибридов первого поколения. Возможен только один вариант генотипа ребенка от такого брака. У ребенка будет развит и слуховой нерв, и улитка - ребенок не будет глухим, в отличие от родителей.
Эпистаз
Эпистаз (противоположное действие генов) - явление, при котором один ген аллельной пары (супрессор) в доминантном (доминантный эпистаз) или рецессивном (рецессивный эпистаз) состоянии может подавлять развитие признака, за развитие которого отвечает другая пара генов.
Широко известным примером рецессивного эпистаза является Бомбейский феномен, названный так в результате зафиксированного случая в индийском городе Бомбеи. Доктор Бхенде обнаружил, что у людей рецессивных по гену h (hh) на поверхности эритроцитов не синтезируются агглютиногены - в результате этого они могут быть универсальными донорами.
Говоря проще о Бомбейском феномене: у людей с генотипом hh всегда обнаруживается первая группа крови при любом генотипе - I A I A , I B I B , I A I B . Ген h подавляет гены I A и I B - на поверхности эритроцитов не образуются агглютиногены A и B.
Пример решения задачи №4
"Редкий рецессивный ген (h) в гомозиготном состоянии обладает эпистатическим действием по отношению к генам I A , I B и изменяет их действие до I группы крови (бомбейский феномен). Определите возможные группы крови у детей, если у мужа II гомозиготная, у жены IV и оба родителя гетерозиготны по эпистатическому гену"
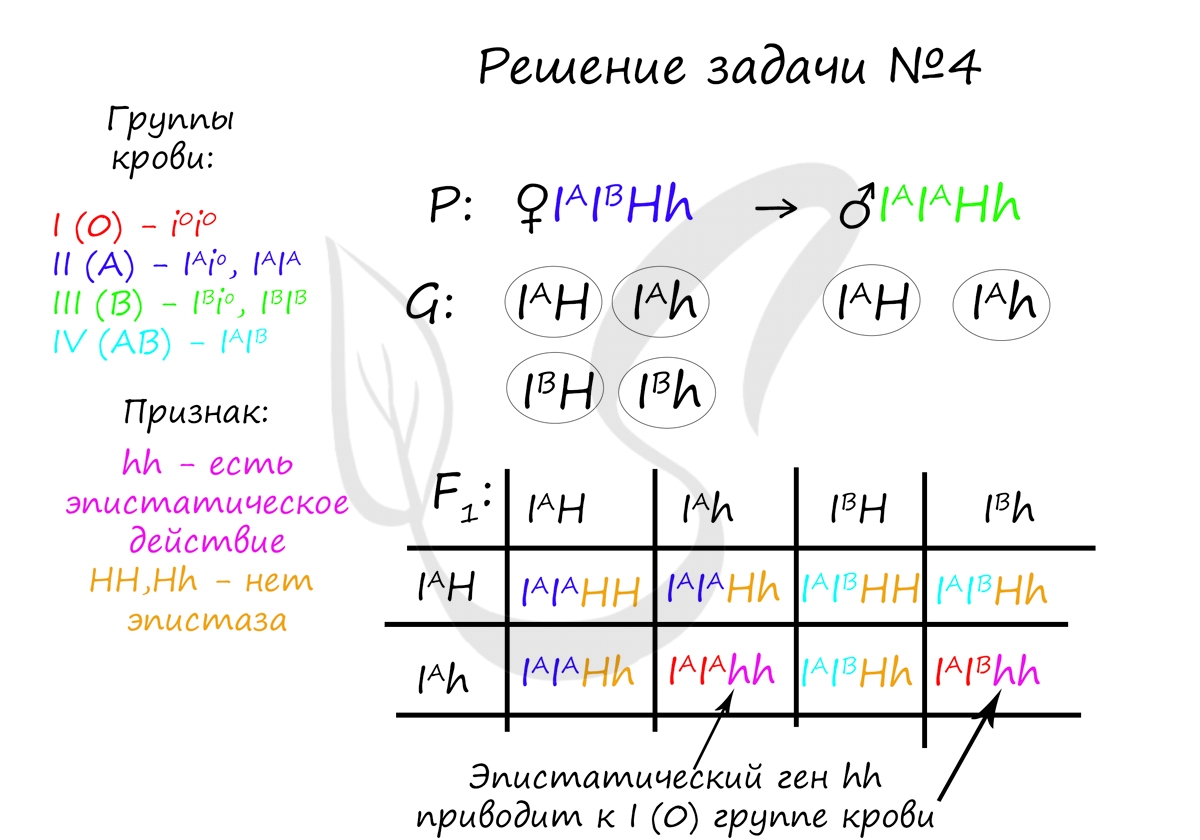
Вероятность рождения детей с i(0) группой крови в данном случае равна 2/8, или 1/4 (25%). Генотипами, у которых будет i(0) группа крови являются: I A I A hh и I A I B hh. Эпистатический рецессивный ген hh в гомозиготном состоянии всегда приводит к i(0) группе крови.
Полимерия
Полимерией называют зависимость определенного признака организма от нескольких пар аллельных генов, обладающих схожим действием. Такие гены называются полимерными. Часто выраженность признака зависит от соотношения доминантных и рецессивных аллелей - то есть чем больше доминантных генов, тем более выражен признак.

У человека полимерное действие генов заложено в наследовании количественных признаков (вес, рост, цвет кожи, давление).
Пример решения задачи №5
"Цвет кожи у мулатов наследуется по типу полимерии. При этом данный признак контролируется 2 аутосомными несцепленными генами. Сын белой женщины и негра женился на белой женщине. Может ли этот ребенок быть темнее своего отца?"
В данном случае полимерия проявляется в том, что чем больше доминантных генов в генотипе (A и B), тем более темный цвет кожи имеет человек. Это правило мы и применим для решения.
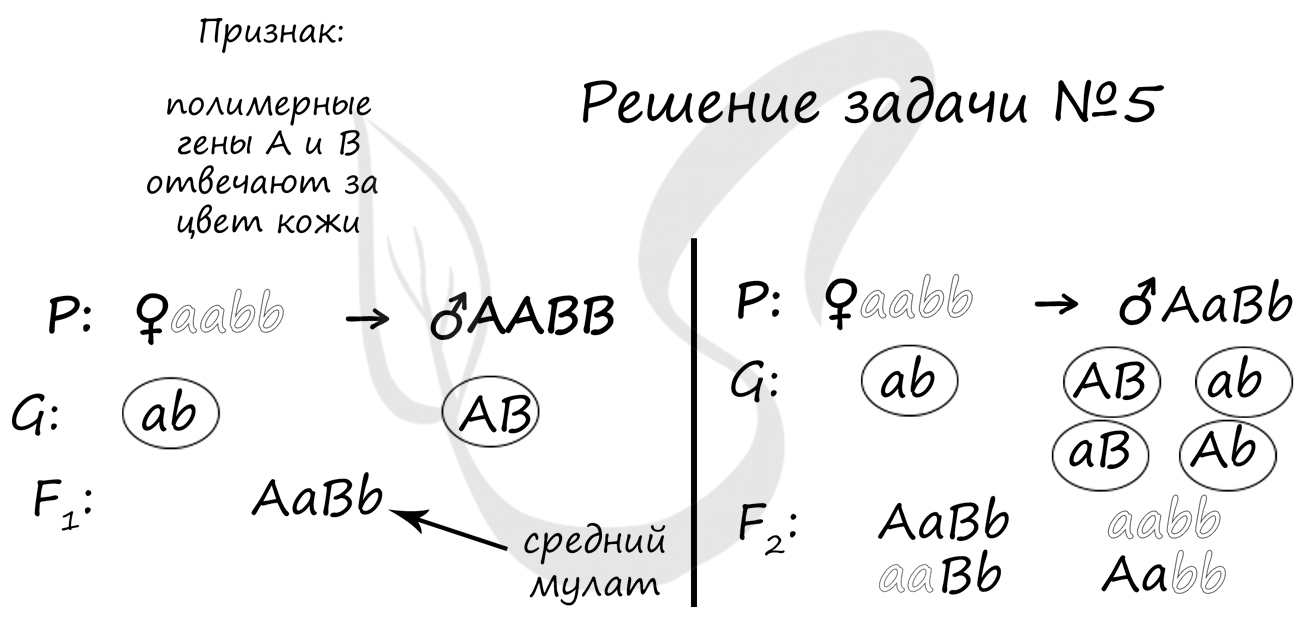
В результате первого брака (вспоминаем закон единообразия Менделя) получается AaBb - средний мулат. По условиям задачи он берет в жены белую женщину aabb. Очевидно, что в этой семье ребенок не может быть темнее своего отца: дети могут быть средними мулатами (AaBb), как отец, светлыми мулатами (aaBb, Aabb), либо белыми, как мать (aabb).
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Теория корпускулярности генов, представляющая их как морфологические и функциональные отдельности, оказалась в противоречии с большой группой фактов, говорящих о наличии так называемого эффекта положения генов. Было установлено, что влияние гена на процессы развития особи зависит не только от его свойств, но и от того, какие гены расположены рядом с ним в хромосоме. Впервые эффект положения генов был открыт на дрозофиле Стертевантом на примере гена Ва r (полосковидные глаза); однако этот пример долгое время оставался уникальным, будучи, кроме того, связан со спецификой неравного кроссинговера. После того как было открыто мощное мутагенное действие ионизирующих излучений, оказалось возможным в неограниченном количестве экспериментально получать разнообразнейшие реорганизации в структуре хромосом. Это привело к получению многообразных случаев изменения взаиморасположения генов в хромосомах, изучение которых показало широкую распространенность явления эффекта положения генов. Проведенные исследования показали, что явление эффекта положения генов представляет собой одну из главных проблем теоретической генетики, которая должна пролить свет на организацию хромосом и на природу первичных функций генов.
Однако, поскольку изменение положения связано со структурными преобразованиями в хромосомах, факты эффекта положения могут быть истолкованы с двух точек зрения. Для хромосомных перестроек характерно возникновение на границе пункта реорганизации (в местах разрывов исходных хромосом и нового соединения фрагментов) таких генетических эффектов, как летальность или изменения в действиях генов этих участков. Эти изменения могут быть результатом как мутаций, так и изменений в функциях генов в силу воздействия на них нового генного окружения. Летальные мутации могут появляться в результате выпадений отдельных локусов или их групп во время возникновения структурных перестроек. В результате долгое время явление эффекта положения во всех этих случаях оставалось возможным, но не обязательным истолкованием полученных фактов. Прямое доказательство существования явления эффекта положения было дано на дрозофиле Н. П. Дубининым и Б. Н. Сидоровым при изучении полученной ими под воздействием рентгеновых лучей транслокации между третьей и четвертой хромосомами, связанной с изменением действия гена hairy (дополнительные щетинки). Микроскопическое изучение гигантских хромосом слюнных желез и опыты по сцеплению показали, что в этой транслокации ген hairy , изменивший свое действие, локализован на некотором расстоянии от точки разрыва в третьей хромосоме. Поэтому удалось получить перекрест между этим геном и точкой разрыва и вывести его из хромосомной перестройки. Во всех случаях, когда ген hairy переводили в нормальную систему хромосом, он приобретал обычную нормальную функцию. Когда он оказывался в системе транслокации, он проявлял измененные свойства. Такая обратимость изменения функции изученного гена исключала мутационную природу изменений; эти эксперименты дали прямые доказательства существования эффекта положения, они исключительно ясно показали, что изменения функции гена могут наступить в результате изменений межгенных связей.
Глубокое своеобразие явлений эффекта положения проявилось в случае изменений функции гена cubitus interruptus у дрозофилы, открытого Н. П. Дубининым и Б. Н. Сидоровым. Было обнаружено, что при разрывах в проксимальной части микрохромосомы (4-я хромосома), где расположен нормальный аллель cubitus interruptus , этот аллель теряет свойство быть доминантным, и у особей, гетерозиготных по рецессивному аллелю, проявляется характерное недоразвитие кубитальной жилки на крыле. Однако этот аллель, дающий в гетерозиготе эффект положения, в гомозиготном состоянии никаких изменений в развитии особей не вызывал.
К настоящему времени проблема эффекта положения гена стала одной из центральных в современной генетике. Природа самого явления еще не ясна. Возможно, что эффект положения генов зависит от изменения в химических взаимоотношениях между первичными продуктами генов, появляющихся на поверхности хромосом. Главный вывод из исследований по эффекту положения генов состоит в установлении того фундаментального положения, что хромосома отнюдь не представляет собой нить с нанизанными на нее отдельными независимыми друг от друга корпускулами — генами, но представляет собой систему, в которой пространственное расположение ее отдельных частей (генов) играет существенную роль. Естественный отбор создает исторические системы новых генотипов, не только опираясь на мутации отдельных генов, но и на новые формы связей между генами внутри хромосомы.
Новые исследования по регуляции генов на молекулярном уровне (см. ниже) раскрыли важнейшие причины эффекта положения в нарушении регуляции действия сложных генов.
Читайте также: