
Эффект пастера биохимия кратко
Обновлено: 08.07.2024
Эффект Пастера – это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода. Биохимический механизм эффекта заключается в конкуренции за пируват между пируватдегидрогеназой, превращающей пируват в ацетил-S-KoA, и лактатдегидрогеназой, превращающей пируват в лактат.
У пируватдегидрогеназы сродство гораздо выше и в обычных аэробных условиях она окисляет большую часть пировиноградной кислоты. Как только поступление кислорода уменьшается (недостаток кровообращения, тромбоз и т.п.) происходит следующее:
- внутримитохондриальные процессы дыхания не идут и НАДН в дыхательной цепи не окисляется,
- моментально накапливающийся в митохондриях НАДН тормозит цикл трикарбоновых кислот.
- ацетил-S-KoA не входит в ЦТК и ингибирует ПВК-дегидрогеназу.
В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную.
При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
Вовлечение других углеводов в гликолиз
Помимо глюкозы энергетически значимыми являются и другие моносахариды: фруктоза, галактоза и манноза с помощью разных ферментативных реакций включаются в гликолитический путь.
Фруктоза является основным источником энергии в диетах, богатых фруктами и сахарозой. Метаболизм фруктозы в мышцах требует одной дополнительной ферментативной реакции. Её катализирует фермент гексокиназа. Продукт этой реакции — фруктозо-6-фосфат напрямую включается в гликолиз.
В печени присутствует гомолог гексокиназы - глюкокиназа. Однако этот фермент фосфорилирует только глюкозу. Печень вовлекает фруктозу в гликолиз с помощью 7 дополнительных реакций, результатом которых является образование дигидроксиацетонфосфата, который изомеризуется с помощью триозофосфатизомеразы в глицеральдегид-3- фосфат. Последний, в свою очередь, также вовлекается в гликолиз.
Галактоза - моносахарид, входящий в состав молочного сахара (лактозы) и присутствующий в молочных продуктах. Галактоза включается в гликолиз с помощью 4 ферментативных реакций, в результате которых образуется глюкозо-6-фосфат.
Манноза - моносахарид, входящий в состав гликопротеинов и полисахаридов. Она вовлекается в гликолиз с помощью двух ферментативных реакций:
1) гексокиназа фосфорилирует маннозу до маннозо-6-фосфата.
2) Фосфоманнозоизомераза изомеризует маннозо-6-фосфат во фруктозо-6-фосфат, вовлекаемый в 3-ю реакцию гликолиза.
Регуляция скорости гликолитических реакций.
Поскольку основное значение гликолиза состоит в синтезе АТФ, его скорость должна коррелировать с затратами энергии в организме.
Большинство реакций гликолиза обратимы (они находятся в термодинамическом равновесии, их скорость очень высока, поэтому они едва ли могут ограничивать скорость гликолиза в целом), за исключением трёх, катализируемых гексокиназой (или глюкокиназой), фосфофруктокиназой и пируваткиназой. Регуляторные факторы, изменяющие скорость гликолиза, а значит и образование АТФ, направлены на необратимые реакции. Показателем потребления АТФ является накопление АДФ и АМФ. Даже небольшой расход АТФ ведёт к заметному увеличению АМФ. Отношение уровня АТФ к АДФ и АМФ характеризует энергетический статус клетки, а его составляющие служат аллостерическими регуляторами скорости как общего пути катаболизма (ОПК), так и гликолиза. Механизмы регуляции скорости гликолиза могут быть следующие:
Эффект Пастера – это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода. Биохимический механизм эффекта заключается в конкуренции за пируват между пируватдегидрогеназой, превращающей пируват в ацетил-S-KoA, и лактатдегидрогеназой, превращающей пируват в лактат.
У пируватдегидрогеназы сродство гораздо выше и в обычных аэробных условиях она окисляет большую часть пировиноградной кислоты. Как только поступление кислорода уменьшается (недостаток кровообращения, тромбоз и т.п.) происходит следующее:
- внутримитохондриальные процессы дыхания не идут и НАДН в дыхательной цепи не окисляется,
- моментально накапливающийся в митохондриях НАДН тормозит цикл трикарбоновых кислот.
- ацетил-S-KoA не входит в ЦТК и ингибирует ПВК-дегидрогеназу.
В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную.
При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
Вовлечение других углеводов в гликолиз
Помимо глюкозы энергетически значимыми являются и другие моносахариды: фруктоза, галактоза и манноза с помощью разных ферментативных реакций включаются в гликолитический путь.
Фруктоза является основным источником энергии в диетах, богатых фруктами и сахарозой. Метаболизм фруктозы в мышцах требует одной дополнительной ферментативной реакции. Её катализирует фермент гексокиназа. Продукт этой реакции — фруктозо-6-фосфат напрямую включается в гликолиз.
В печени присутствует гомолог гексокиназы - глюкокиназа. Однако этот фермент фосфорилирует только глюкозу. Печень вовлекает фруктозу в гликолиз с помощью 7 дополнительных реакций, результатом которых является образование дигидроксиацетонфосфата, который изомеризуется с помощью триозофосфатизомеразы в глицеральдегид-3- фосфат. Последний, в свою очередь, также вовлекается в гликолиз.
Галактоза - моносахарид, входящий в состав молочного сахара (лактозы) и присутствующий в молочных продуктах. Галактоза включается в гликолиз с помощью 4 ферментативных реакций, в результате которых образуется глюкозо-6-фосфат.
Манноза - моносахарид, входящий в состав гликопротеинов и полисахаридов. Она вовлекается в гликолиз с помощью двух ферментативных реакций:
1) гексокиназа фосфорилирует маннозу до маннозо-6-фосфата.
2) Фосфоманнозоизомераза изомеризует маннозо-6-фосфат во фруктозо-6-фосфат, вовлекаемый в 3-ю реакцию гликолиза.
Регуляция скорости гликолитических реакций.
Поскольку основное значение гликолиза состоит в синтезе АТФ, его скорость должна коррелировать с затратами энергии в организме.
Большинство реакций гликолиза обратимы (они находятся в термодинамическом равновесии, их скорость очень высока, поэтому они едва ли могут ограничивать скорость гликолиза в целом), за исключением трёх, катализируемых гексокиназой (или глюкокиназой), фосфофруктокиназой и пируваткиназой. Регуляторные факторы, изменяющие скорость гликолиза, а значит и образование АТФ, направлены на необратимые реакции. Показателем потребления АТФ является накопление АДФ и АМФ. Даже небольшой расход АТФ ведёт к заметному увеличению АМФ. Отношение уровня АТФ к АДФ и АМФ характеризует энергетический статус клетки, а его составляющие служат аллостерическими регуляторами скорости как общего пути катаболизма (ОПК), так и гликолиза. Механизмы регуляции скорости гликолиза могут быть следующие:

Папиллярные узоры пальцев рук - маркер спортивных способностей: дерматоглифические признаки формируются на 3-5 месяце беременности, не изменяются в течение жизни.

Опора деревянной одностоечной и способы укрепление угловых опор: Опоры ВЛ - конструкции, предназначенные для поддерживания проводов на необходимой высоте над землей, водой.

Организация стока поверхностных вод: Наибольшее количество влаги на земном шаре испаряется с поверхности морей и океанов (88‰).
Эффект Пастера – это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода.
Луи Пастер, занимавшийся вопросами виноделия, наблюдал подобный феномен при производстве вина. Забегая вперед, отметим, что спиртовое брожение весьма похоже на гликолиз, только вместо молочной кислоты из пирувата образуется спирт.
Биохимический механизм эффекта Пастера заключается в конкуренции между пируватдегидрогеназой, превращающей пируват в ацетил-S-КоА, и лактатдегидрогеназой, превращающей пируват в лактат. При отсутствии кислорода внутримитохондриальные процессы дыхания не идут, цикл трикарбоновых кислот тормозится и накапливающийся ацетил-S-КоА ингибирует ПВК‑дегидрогеназу. В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную. При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
Важно то, что пировиноградная кислота является для клетки токсичным веществом, и клетке необходимо избавиться от нее каким угодно образом. Так как через мембраны она не проходит, то обезвреживание достигается превращением пирувата 1) в лактат; 2) в ацетил-S-КоА; 3) в аланин (см "Аланинаминотрансфераза"), 4) в оксалоацетат.
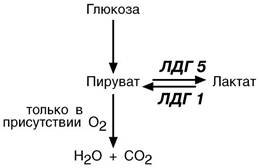
Иллюстрацией к сказанному служит отличие изоферментов лактатдегидрогеназ (ЛДГ) друг от друга. Сердечный изофермент ЛДГ-1 обладает высоким сродством к молочной кислоте и "стремится" поднять концентрацию пирувата с целью его включения в ЦТК и получения энергии для деятельности миокарда. Большое количество митохондрий и поступление сюда лактата из других органов обеспечивает работу сердца при аэробных условиях. При нехватке кислорода свойства ЛДГ-1 не изменятся, он по-прежнему будет сдвигать реакцию в сторону продукции пировиноградной кислоты. Изофермент скелетной мышцы ЛДГ-5 обладает высоким сродством к пирувату, при отсутствии кислорода в клетке быстро и эффективно превращает его в лактат, легко проникающий сквозь мембраны. Таким образом, в анаэробных условиях сильнее будет страдать сердечная мышца, что, собственно говоря, и наблюдается в медицинской практике.
Эффект Пастера – это снижение потребления глюкозы и прекращение продукции молочной кислоты клеткой в присутствии кислорода.
Луи Пастер, занимавшийся вопросами виноделия, наблюдал подобный феномен при производстве вина. Забегая вперед, отметим, что спиртовое брожение весьма похоже на гликолиз, только вместо молочной кислоты из пирувата образуется спирт.
Биохимический механизм эффекта Пастера заключается в конкуренции между пируватдегидрогеназой, превращающей пируват в ацетил-S-КоА, и лактатдегидрогеназой, превращающей пируват в лактат. При отсутствии кислорода внутримитохондриальные процессы дыхания не идут, цикл трикарбоновых кислот тормозится и накапливающийся ацетил-S-КоА ингибирует ПВК‑дегидрогеназу. В этой ситуации пировиноградной кислоте не остается ничего иного как превращаться в молочную. При наличии кислорода ингибирование ПВК-дегидрогеназы прекращается и она, обладая большим сродством к пирувату, выигрывает конкуренцию.
Важно то, что пировиноградная кислота является для клетки токсичным веществом, и клетке необходимо избавиться от нее каким угодно образом. Так как через мембраны она не проходит, то обезвреживание достигается превращением пирувата 1) в лактат; 2) в ацетил-S-КоА; 3) в аланин (см "Аланинаминотрансфераза"), 4) в оксалоацетат.
Иллюстрацией к сказанному служит отличие изоферментов лактатдегидрогеназ (ЛДГ) друг от друга. Сердечный изофермент ЛДГ-1 обладает высоким сродством к молочной кислоте и "стремится" поднять концентрацию пирувата с целью его включения в ЦТК и получения энергии для деятельности миокарда. Большое количество митохондрий и поступление сюда лактата из других органов обеспечивает работу сердца при аэробных условиях. При нехватке кислорода свойства ЛДГ-1 не изменятся, он по-прежнему будет сдвигать реакцию в сторону продукции пировиноградной кислоты. Изофермент скелетной мышцы ЛДГ-5 обладает высоким сродством к пирувату, при отсутствии кислорода в клетке быстро и эффективно превращает его в лактат, легко проникающий сквозь мембраны. Таким образом, в анаэробных условиях сильнее будет страдать сердечная мышца, что, собственно говоря, и наблюдается в медицинской практике.
Снижение скорости потребления глюкозы и прекращение накопления лак-тата в присутствии кислорода носит название эффекта Пастера. Впервые это явление наблюдал Л. Пастер во время своих широко известных исследований роли брожения в производстве вина. В дальнейшем было показано, что эффект Пастера наблюдается также в животных и растительных тканях, где кислород тормозит анаэробный гликолиз. Значение эффекта Пастера, т.е. перехода в присутствии кислорода от анаэробного гликолиза или брожения к дыханию, состоит в переключении клетки на наиболее эффективный и экономичный путь получения энергии. В результате скорость потребления субстрата, например глюкозы, в присутствии кислорода снижается. Молекулярный механизм эффекта Пастера заключается, по-видимому, в конкуренции между системами дыхания и гликолиза (брожения) за АДФ, используемый для образования АТФ. Как известно, в аэробных условиях значительно эффективнее, чем в анаэробных, происходят удаление Piи АДФ, генерация АТФ, а также регенерирование НАД + , окисленного из восстановленного НАДН. Иными словами, уменьшение в присутствии кислорода количества Рi и АДФ и соответствующее увеличение количества АТФ ведут к подавлению анаэробного гликолиза.
Эффект Пастера является ингибирование брожения процессов с помощью кислорода .
Резюме
Открытие
Этот эффект был открыт в 1857 или 1861 году Луи Пастером , который обнаружил, что аэрирование дрожжевого бульона вызывает усиление развития дрожжей и уменьшение производства спирта при спиртовом брожении .
Механизм
Поскольку дрожжи являются факультативными анаэробами , они могут получать энергию с помощью различных метаболических путей. Когда концентрация кислорода низкая, продукт гликолиза ( пируват ) превращается в этанол и диоксид углерода , и выход при производстве АТФ остается низким (2 моля АТФ на моль использованной глюкозы). Если концентрация кислорода увеличивается, пируват превращается в ацетил-КоА, который может использоваться циклом Кребса , что увеличивает выход до 38 моль АТФ на моль глюкозы .
В анаэробных условиях скорость использования глюкозы выше, а количество продуцируемого АТФ ниже. В аэробных условиях (с кислородом) скорость гликолиза снижается, поскольку повышенное производство АТФ действует как аллостерический ингибитор на этом метаболическом пути.
Практическое значение
Все процессы производства алкоголя должны осуществляться в условиях без кислорода и, следовательно, без воздуха: производство вина , производство пива, производство биоэтанола . И наоборот, для эффективного производства дрожжей рекомендуется насыщать кислородом середину с помощью вентиляции.
Читайте также: