
Древние вымершие папоротникообразные кратко
Обновлено: 25.06.2024
Никогда не прекращайте искать вокруг себя удивительное и интересное, ведь когда вам кажется, что ничего такого для вас нигде больше нет, то это лишь значит, что вы слишком глупы и невнимательны. Один обычный куст, который растет где-то в горшке, либо же около подъезда, может скрывать в себе больше тайн и загадок, чем Египетские пирамиды или целый сезон Гравити фоллз.Приготовьтесь узнать об одном из потрясающих древних растений, у которого на самом деле нет ни цветов(!), ни стеблей (!!), ни листьев (. ), ни даже мелочи на проезд(?). Поехали.
Начнем обо всем по порядку. Как я уже писал выше, папоротники — древнейший отдел высших растений, которые растут по всему земному шару. Раньше, когда по земле шастали динозавры (даже раньше, лет так на 200 миллионов), папоротники занимали огромные площади и формировали собой первые лесные территории. Привычных нам деревьев тогда и в помине не было, зато были древовидные папоротники, которых сегодня почти не осталось. Повсюду росли только плауны, хвощи и папоротники, Земля была молодой, свежей и луна была немного ближе. Нигде не валялось ни одной пустой банки от балтики, ни упаковок от сухариков из ашана, ни даже пластиковых стаканчиков.
Орляк обыкновенный. Растет в нашем климате, срез ветки напоминает государственного орла. Поэтому Орляк.
Орляк обыкновенный. Растет в нашем климате, срез ветки напоминает государственного орла. Поэтому Орляк.
Вопреки славянским легендам, что огненно-красный цветок папоротника обладает волшебными свойствами, ни одно папоротниковое растение не умеет делать цветы. Папоротники произошли от Риниофитов — вымершего около 400 млн лет назад отдела первых примитивных сухопутных растений, и размножаются не семенами, — до них растения додумаются гораздо позже.
Один из немногих сохранившихся сегодня видов древовидных папоротников — Cyathea cooperi. Настоящий небольшой кусочек Палеозоя.
Один из немногих сохранившихся сегодня видов древовидных папоротников — Cyathea cooperi. Настоящий небольшой кусочек Палеозоя.
Папоротники размножаются вегетативно, а еще. спорами. Да-да, не только грибы так умеют! Эта способность дает папоротникам огромный буст к размножению и позволяет путешествовать на огромные расстояния, заселяя светлые леса, вроде хвойных и березовых, и образовывать там крупные заросли. Лучше всего растут там, где тепло и сыро — в тропических и субтропических зонах. Легко адаптируются к любым изменениям среды, могут расти при низкой освещенности и переувлажнении почвы, что делает их хорошими комнатными растениями. Даже если условия слишком суровые, а света почти нет, папоротник не перестанет существовать, а просто побледнеет и потеряет пару очков декоративности за счет небольшого облысения.
Так как папоротникам не нужны цветы, чтобы размножаться, одним из их скиллов является невероятная плодовитость. Одно растение образует дофига спор, которые хранятся в специальных мешочках на нижней части листиков. После созревания мешочки раскрываются, и легкие, как пыль, споры разлетаются по ветру. Из них вырастают гаметофиты — мелкие листочки, в которых происходит оплодотворение. На нижней части этой непонятной зеленой штуки, похожей на сердечко, образуются сперматозоиды и яйцеклетки, и, когда проходит дождь, под листочком образуется влажная среда, по которой сперматозоиды переплывают к яйцеклеткам. В итоге образуется бесполое растение, через пару лет вырастающее в большой красивый папоротник.
От папоротников через пару сотен миллионов лет произошли первые голосеменные, а от них покрытосеменные (цветковые), с появлением которых число папоротников сильно поубавилось. В отличие от своих молодых потомков, наш папоротник настоящих листьев не имеет. Их роль у него заменяет Вайя — листоподобный орган, что-то среднее между веткой, побегом и листом, но не являющееся ни тем, ни другим, и ни третьим. Четкое разграничение между стеблем и листом начало появляться только у первых голосеменных, у папоротников же все это смешано и слабо выражено. "Ветвь" папоротника в эмбриональном состоянии напоминает улитку, которая растет очень медленно и раскрывается только через несколько лет после прорастания. Мелкие разделенные листовые пластинки, которые имеются у некоторых видов, растут в одной плоскости и напоминают настоящие листья, как у современных растений.
Папоротники — самая большая группа споровых растений, и включает в себя 48 семейств и больше 10 000 видов . В диком виде растут в лесах — в нижнем и верхнем ярусах, на ветвях и стволах крупных деревьев — как эпифиты (растения, растущие на других растениях), в расщелинах скал, на болотах, в реках и озерах, на стенах городских домов, на сельскохозяйственных землях как сорняки, и по обочинам дорог. Ты, возможно, их не замечаешь, но на самом деле папоротники везде. Стоит лишь поглядеть чуть-чуть внимательнее, и ты сам убедишься в этом. А гуляя в лесу по зарослям папоротника можешь представлять себя динозавром, потому что именно так, скорее всего, и выглядели леса в те времена.
ПÁПОРОТНИКИ ИСКОПÁЕМЫЕ, вымершие представители класса Папоротники ( Filicopsida ). Возникли в начале среднего девона, предшественники точно не известны; вероятными предками считаются либо плауновидные, либо непосредственно проптеридофиты. Наряду с травянистыми для П. и. характерны и древовидные формы, особенно для позднего девона и юры. Группа пережила расцвет в конце карбона и относительный в юре – раннем мелу.
Представители класса отличаются рассечёнными листьями (вайями), представленными перьями нескольких порядков. Спорангии располагаются на нижней стороне листа или по краю.

Ископаемые папоротники (реконструкция): 1 – Psaronius (сем. Marattiaceae), карбон ; 2 – Phlebopteris (сем. Matoniaceae) , юра ; 3 – Todites (сем. Osmundaceae), юра .
Мараттиевые (Marattiaceae) – семейство папоротников, у которых спорангии размещены вдоль главной жилки и срастаются в синангии. Множество древовидных форм в палеозое и мезозое. Конец девона – ныне.
Pecopteris – формальный род (ф. р.) позднепалеозойских древовидных папоротников сем. Мараттиевые . Листья крупные, состоящие из небольших прямых листовых пластинок, закруглённых на конце. Пластинки приросшие к черешку 2-го порядка, который в свою очередь прикреплен к черешку 1-го порядка, образуя форму расчёски. Поздний девон – поздняя пермь.
Матониевые ( Matoniaceae ) – семейство папоротников с крупными дланевидными вайями. Сорусы (сгруппированные спорангии) располагаются в 2 ряда вдоль главной жилки пёрышка. Поздний триас – ныне.
Диптериевые (Dipteridaceae) – семейство, представители которого отличаются крупными листьями и чётким сетчатым жилкованием. Спорангии располагаются внутри петель жилок последнего порядка. Известны с начала триаса.
Hausmannia – род сем. Диптериевые (Dipteridaceae). Крупные наземные папоротники с длинными ползучими волосистыми корневищами и неглубоко-лопастными листьями. Пластинка сидячая, разделена на 2 симметричных половинки, посередине с полостью. Жилкование разветвлённое и полуразделённое дихотомически. Конец триаса – ранний мел.
Схизейные (Schizaeaceae) – семейство травянистых папоротников, преимущественно жарких климатических зон. Стебли прямостоячие, реже ползучие, листья перисто- и пальчаторассечённые. В ископаемом состоянии встречаются главным образом споры. Стерильные листья иногда относят к ф.р. Sphenopteris. Карбон – ныне.
Глейхениевые ( Gleicheniaceae ) – семейство травянистых папоротников, иногда вьющихся. Вайи повторно-вильчатые или вееровидные. С порангии довольно крупные, продольно раскрывающиеся, с косым кольцом. Споры трилетные. Поздний триас – ныне.
Gleichenia – род папоротников одноимённого семейства с тонкими дихотомически разветвлёнными корневищами. Листья дважды или трижды дихотомические. Поздний триас – ныне.
Cladophlebis – ф. р. папоротников с неопределённым систематическим положением , выделенный только по стерильным листьям и их частям. Листья дважды или четырежды перистые, линейные или близкие к треугольным очертаниям. Пермь – верхний мел всего мира.









Pecopteris sp. П еро последнего порядка . Пермь, казанский ярус. Исаклинский флористический комплекс, 2013.





Находки остатков П. и. на территории Самарской области отмечаются довольно редко, однако вертикальное их распространение охватывает почти весь срок существования данной группы. В девонских континентальных отложениях региона остатки вегетативных органов П. и. неизвестны. Тем не менее, уже в бийском горизонте эйфельского яруса среднего девона обнаружены предположительно относящиеся к П. и. споры Granulatisporites minor. Для эйфельского и живетского ярусов характерны споры ф. р. Leiotriletes, до некоторой степени сопоставляемого с папоротниками: L. simplex, L. laevis, L. nigratus. Из кизеловского горизонта турне (нижний карбон) определён комплекс спор, содержащий L. cibotioformis, L. subintortus, L. laevis. Отложения визейского возраста (радаевский горизонт) заключают углистые прослои с растительными остатками, в которых обнаружены споры Stenozonotriletes reticulatus, S. rauseri и Knoxisporites literatus. Из терригенных толщ московского яруса (верейский горизонт) известны G. [= Microreticulatisporites? ] minor, S. conformis, S. cycloides, Ambitisporites pumilus, L. inermis . К сожалению, судить о таксономическом составе папоротниковой флоры карбона, опираясь только на данные спорово-пыльцевых комплексов (СПК), нельзя.
Отпечатки и фитолеймы фрагментов перьев П. и. известны в отложениях казанского яруса пермской системы на северо-востоке Самарской обл. В видовом составе флористических комплексов определены П. и. , остатки которых в известных местонахождениях представлены только стерильными листьями Pecopteris helenaeana (принадлежат мараттиевым), P. micropinnata и ближе не определённых видов того же ф. р. Эти растения, скорее всего, принадлежали мезофильной (промежуточной между влаголюбивой и засухоустойчивой) растительной ассоциации и занимали подчинённое положение в растительном сообществе.
После длительного континентального перерыва П. и. известны для среднеюрского времени из песчаников, относимых ныне к байосу, на Самарской Луке и в Чапаевске. Здесь были встречены остатки диптериевых папоротников Hausmannia crenata, H. volgensis, матониевый папоротник Phlebopteris sp., Coniopteris sp . из сем. циатейные (Cyatheaceae) , папоротники ближе не установленного систематического положения Sphenopteris sp., Cladophlebis sp.
Помимо находок вегетативных частей, в байосских отложениях известны СПК, в составе которых безусловно присутствуют П. и. С юго-востока региона (с. Ореховка) имеется определение спор глейхениевых – Gleichenia stellata. Для СПК в палеокарстовых образованиях близ Переволокского перешейка приведён целый ряд приблизительных определений, выполненных специалистами по современной флоре из Казанского университета, из которых можно сделать вывод о присутствии осмундовых (сем. Osmundaceae), глейхениевых, представителей Acrostichum и Adiantum из птериевых (сем. Pteridaceae), диптериевых (вероятно, преобладающий род Hausmannia), схизейных. Также для СПК байоса–бата указывают ф. р. Tripartina и Leiotriletes.

Споры папоротников из комплекса у с. Печерск ( Сызранский р-н ). Юра, байосский ярус, глуб. 27 м: 1 – Osmundacidites, 2 – Gleicheniidites, 3, 4 – Pteridaceae (3 – Adiantum, 4 – Acrostichum), 5 – Dipteridaceae, 6 – Schizaeaceae, 7 – Tripartina, 8 – Leiotriletes .
Только в составе СПК известны папоротники в пограничных верхнеюрско-нижнемеловых (от волжских до нижнебарремских) отложениях Предволжья. В это время происходит расцвет глейхениевых (ксерофиты, до 60% в спектрах) и, в несколько меньшей степени, схизейных (влаголюбивые, до 7%). Оба семейства особенно широко и разнообразно представлены в рязанском региоярусе (= берриасский ярус), в дальнейшем же разнообразие схизейных и количество других влаголюбивых групп (мараттиевые, осмундовые) значительно уменьшается. В биостратиграфии на данной территории используются формальные виды: из глейхениевых – Gleicheniidites senonicus (с конца волжского века, массовый вид), Plicifera delicata (с начала рязанского века), G . toriconcavus , Ornamentifera granulata (готерив); из схизейных – Trilo b osporites asper (волжский региоярус), Cicatricosisporites ludbrooki , C . exilis (рязанский региоярус), Concavisporites dubia (готерив). Помимо них, для этого времени известны представители других ф. р. : из глейхениевых – Clavifera ; схизейных – Impardecispora , Plicatella , Anemia , Appendicisporites , Lygodiumsporites ; матониевых – Dictyophyllidites ; мараттиевых – Marattisporites ; осмундовых – Todisporites ; циатейных – Cyathidites ; диптериевых.
Из палеогена Самарской обл. имеется единственная (2015) находка представителя вымершего рода Speirseopteris телиптерисовых папоротников (сем. Thelypteridaceae) в виде стерильных пёрышек листа папоротника в песчанике. Род был ранее известен лишь из палеоценовых отложений Канады.
Сведения о П. и. акчагыльского века неполны; по СПК известны неопределённые представители рода Gleichenia и сем. Многоножковые (Polypodiaceae). В четвертичные межледниковья папоротники играли существенную роль в лесных сообществах. В СПК мелекесского аллювия (перед началом донского оледенения) присутствуют споры Botrychium lunarium (сем. Гроздовниковые), а СПК жигулёвского аллювия (между донской и окской эпохами оледенения), помимо этого вида, содержат Osmunda cinnamomea, O. claytoniana (сем. Осмундовые), страусник обыкновенный Struthiopteris filicastrum (сем. Оноклеевые), Azolla filiculoides (сем. Сальвиниевые). Позднее, в красноярском аллювии (между днепровским и московским оледенениями), определён только B. boreale.
Древовидные папоротники десятки миллионов лет были доминирующими растениями на большей части поверхности Земли. Они процветали, поскольку в древности (в частности, в Юрском периоде) климат на планете был равномерно теплым и более влажным, т.е. были созданы все подходящие условия для роста древовидных папоротников. Однако потом материки стали отдаляться от экватора, поэтому климат на Земле изменился. Начался процесс оледенения, в некоторых местах выпадало очень мало осадков. Именно из-за климатических изменений древовидные папоротники вымерли везде, кроме современного экваториального пояса.
Папоротники, впервые появившись середине девона, широко распространились и достигали расцвета в каменноугольном периоде.
Уже в девоне, вместе с настоящими папоротниками появились первые семенные папоротники. Среди папоротников были как мелкие травянистые растения, так и крупные древовидные формы.
Папоротники вместе с лепидодендронами, сигилляриями, каламитами составляли тенистые леса.
Настоящие папоротники размножаются спорами. У некоторых развиваются специальные спороносные листья.
Первые папоротники появились в девоне. По внешнему виду они напоминали псилофиты: отсутствовали настоящие листья,одиночные спорангии располагались на концах листоподобных ветвей. Позднее у них появились настоящие листья – вайи. Среди папоротников встречались крупные растения, которые сходны с современными древовидными папоротниками.
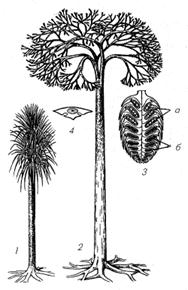
Лепидодендрон (лат. Lepidodendron) — род вымерших древоподобных плауновидных растений, существовавший в каменноугольном периоде и составлявший часть флоры угольных лесов. Высота растения, по данным палеобиологии, составляла от 10 до 35 метров, диаметр ствола — до одного метра.

Археоптерис (лат. Archaeopteris, от др.-греч. ἀρχαῖος — древний, πτέρις — папоротник) – древнейшее крупное папоротникообразное растение, которое существовало с девона по карбон. Высота 20 м. листьями был покрыт лишь вверху, напоминали ствол пальмы.
Каламиты(Calamités) — ископаемые растения, близкие к ныне живущим хвощам. Это были древовидные растения, имевшие мощные членистые, полые внутри стебли, перегороженные поперечными диафрагмами. Междоузлия были неодинаковой длины, в нижней части стебля короче, нежели в верхней; поверхность междоузлий покрыта продольными ребрами, чередующимися в двух последних междоузлиях. Стебли ветвились большею частью только в верхней части; ветви располагались рассеянно или мутовчато. Листьев, по-видимому, не было, или они были слабо развиты. Сосудистые пучки на поперечном разрезе клиновидные, узкие, разделенные полосками паренхиматической ткани; кора сильно развита, с большими пустотами. Изогнутым, клиновидным основанием стебли прикреплялись к подземному корневищу. Там, где эти стебли встречаются в их первоначальном положении, они образуют группу, отходящую от общего центра, т. е. корневища, откуда можно заключить, что стебли были однолетние, подобно таким же высоким стеблям хвощей из Триаса. К. росли повсеместно на земле во времена каменноугольной формации. Остатки их встречаются в виде тонких пластинок из каменного угля или в виде сдавленных кусков стебля, состоящего из минеральной массы, соответствующей центральной полости стебля, и из слоя каменного угля, соответствующего коре и древесине; эта каменноугольная покрышка легко обваливается; обнаженная минеральная масса имеет более сильные ребра, нежели кора; ребра соответствуют желобкам внутренней поверхности древесины, а желобки — выступам сосудов. Спорангии у К. были собраны в колосья, на которых плодящие мутовки листьев чередовались с бесплодными; споры были одного рода (isosporae).


У ихтиостег были ноги, но они, возможно, не использовались для ходьбы по суше. Ихтиостеги имели хвостовой плавник и некоторые органы чувств, функционирующие только в воде. Тело их было покрыто мелкими чешуйками.
По мнению некоторых учёных, ихтиостеги могут считаться переходными формами между кистеперыми рыбами и наземными позвоночными.
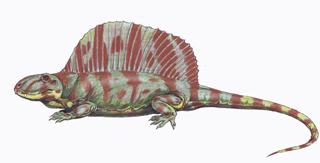
Пеликозавры (лат. Pelycosauria) — отряд наиболее примитивных синапсид. Сохраняли типично рептильную физиологию и внешний облик (за исключением того, что не имели достаточно развитых чешуйных покровов). еликозавры в целом находились на уровне организации, характерном для ранних амниот. Внешностью большинство из них походило на ящериц; они имели двояковогнутые позвонки и хорошо сохранившиеся брюшные рёбра. Однако зубы их сидели в альвеолах (как у терапсид и млекопитающих), а у некоторых пеликозавров (например, у сфенакодона) намечалась — пусть и слабовыраженная — дифференциация зубов. Среди пеликозавров встречались как крупные (длиной 3—6,5 м), так и существенно более мелкие (1—2 м) животные — хищные и растительноядные. В конце карбона и начале перми (300—270 млн лет тому назад) пеликозавры доминировали в наземных экосистемах.

Папоротники, впервые появившись середине девона, широко распространились и достигали расцвета в каменноугольном периоде.
Уже в девоне, вместе с настоящими папоротниками появились первые семенные папоротники. Среди папоротников были как мелкие травянистые растения, так и крупные древовидные формы.
Папоротники вместе с лепидодендронами, сигилляриями, каламитами составляли тенистые леса.
Настоящие папоротники размножаются спорами. У некоторых развиваются специальные спороносные листья.
Первые папоротники появились в девоне. По внешнему виду они напоминали псилофиты: отсутствовали настоящие листья,одиночные спорангии располагались на концах листоподобных ветвей. Позднее у них появились настоящие листья – вайи. Среди папоротников встречались крупные растения, которые сходны с современными древовидными папоротниками.
Лепидодендрон (лат. Lepidodendron) — род вымерших древоподобных плауновидных растений, существовавший в каменноугольном периоде и составлявший часть флоры угольных лесов. Высота растения, по данным палеобиологии, составляла от 10 до 35 метров, диаметр ствола — до одного метра.
Археоптерис (лат. Archaeopteris, от др.-греч. ἀρχαῖος — древний, πτέρις — папоротник) – древнейшее крупное папоротникообразное растение, которое существовало с девона по карбон. Высота 20 м. листьями был покрыт лишь вверху, напоминали ствол пальмы.
Каламиты(Calamités) — ископаемые растения, близкие к ныне живущим хвощам. Это были древовидные растения, имевшие мощные членистые, полые внутри стебли, перегороженные поперечными диафрагмами. Междоузлия были неодинаковой длины, в нижней части стебля короче, нежели в верхней; поверхность междоузлий покрыта продольными ребрами, чередующимися в двух последних междоузлиях. Стебли ветвились большею частью только в верхней части; ветви располагались рассеянно или мутовчато. Листьев, по-видимому, не было, или они были слабо развиты. Сосудистые пучки на поперечном разрезе клиновидные, узкие, разделенные полосками паренхиматической ткани; кора сильно развита, с большими пустотами. Изогнутым, клиновидным основанием стебли прикреплялись к подземному корневищу. Там, где эти стебли встречаются в их первоначальном положении, они образуют группу, отходящую от общего центра, т. е. корневища, откуда можно заключить, что стебли были однолетние, подобно таким же высоким стеблям хвощей из Триаса. К. росли повсеместно на земле во времена каменноугольной формации. Остатки их встречаются в виде тонких пластинок из каменного угля или в виде сдавленных кусков стебля, состоящего из минеральной массы, соответствующей центральной полости стебля, и из слоя каменного угля, соответствующего коре и древесине; эта каменноугольная покрышка легко обваливается; обнаженная минеральная масса имеет более сильные ребра, нежели кора; ребра соответствуют желобкам внутренней поверхности древесины, а желобки — выступам сосудов. Спорангии у К. были собраны в колосья, на которых плодящие мутовки листьев чередовались с бесплодными; споры были одного рода (isosporae).
У ихтиостег были ноги, но они, возможно, не использовались для ходьбы по суше. Ихтиостеги имели хвостовой плавник и некоторые органы чувств, функционирующие только в воде. Тело их было покрыто мелкими чешуйками.
По мнению некоторых учёных, ихтиостеги могут считаться переходными формами между кистеперыми рыбами и наземными позвоночными.
Пеликозавры (лат. Pelycosauria) — отряд наиболее примитивных синапсид. Сохраняли типично рептильную физиологию и внешний облик (за исключением того, что не имели достаточно развитых чешуйных покровов). еликозавры в целом находились на уровне организации, характерном для ранних амниот. Внешностью большинство из них походило на ящериц; они имели двояковогнутые позвонки и хорошо сохранившиеся брюшные рёбра. Однако зубы их сидели в альвеолах (как у терапсид и млекопитающих), а у некоторых пеликозавров (например, у сфенакодона) намечалась — пусть и слабовыраженная — дифференциация зубов. Среди пеликозавров встречались как крупные (длиной 3—6,5 м), так и существенно более мелкие (1—2 м) животные — хищные и растительноядные. В конце карбона и начале перми (300—270 млн лет тому назад) пеликозавры доминировали в наземных экосистемах.
Читайте также: