
Ауксины это в биологии кратко
Обновлено: 28.06.2024
АУКСИ́НЫ (от греч. αὔ ξω – увеличивать, выращивать, расти), гормоны растений, регулирующие их рост и развитие. К синтезу А. способны также некоторые фитопатогенные грибы и бактерии. А. усиливают рост растений за счёт стимуляции растяжения клеток. К природным А. относятся индолилуксусная кислота (ИУК), индолилмасляная кислота (ИМК) и некоторые др. производные индола, фенилуксусная кислота. Они интенсивно образуются в активно делящихся клетках верхушек побегов, в молодых листьях и в меньшей мере – в корнях и транспортируются к клеткам-мишеням. В стебле, напр., поток А. направляется от его верхушки к кончику корня (со скоростью ок. 1 см/ч). В создании направленного потока А. участвуют спец. белки-переносчики. При изменении факторов внешней среды (направления освещения или силы тяжести) поток А. быстро меняется: через 10–15 мин А. направляются на более затенённую или на нижнюю часть органа (стебля, корня и др.), из-за чего меняется скорость роста на освещённой (затенённой) или верхней (нижней) части органа, что вызывает ростовой изгиб (тропизм). Действуя на уровне клетки, молекула А. связывается с соответствующим клеточным рецептором (напр., белком ABP 1), вызывая поступление ионов $\ce>$ в клетку. Затем происходит активизация протонных насосов плазматич. мембраны, в результате чего ионы $\ce$ поступают из цитоплазмы в клеточную стенку, приводя к её размягчению и растяжению (удлинению) клетки под действием тургорного давления. В целом растении А. направляют транспорт питательных веществ в интенсивно растущие побеги, листья, корни и плоды. А. являются ключевым фактором органогенеза: образования зачатков листьев, боковых и придаточных корней, дифференцировки проводящей системы, завязывания и роста плодов, подавления роста боковых почек и др. процессов. В растениях А. взаимодействуют с др. фитогормонами; в частности они индуцируют образование этилена, который в свою очередь тормозит транспорт и усиливает распад А. Препараты А. (гетероауксин – смесь ИУК и ИМК) используют для стимуляции корнеобразования у черенков. Синтетич. аналоги А. – 1-нафтил- и 2-нафтоксиуксусные кислоты, хлорированные производные бензойной, феноксиуксусной и пиколиновой кислот, используют в качестве регуляторов роста и гербицидов (в борьбе с двудольными сорными растениями). См. также Фитогормоны .
группа гормонов растений. Регулируют на разных этапах жизни растения его рост, дифференцировку органов, ростовые реакции на свет и силу тяжести. По химической природе — производные индола. Основной представитель — индолилуксусная кислота.
АУКСИ́НЫ, группа гормонов растений. Регулируют на разных этапах жизни растения его рост, дифференцировку органов, ростовые реакции на свет и силу тяжести. По химической природе — производные индола. Основной представитель — индолилуксусная кислота (см. ИНДОЛИЛУКСУСНАЯ КИСЛОТА) .
* * *
АУКСИ́НЫ (от греч. auxano — расту), группа фитогормонов (см. ФИТОГОРМОНЫ) . Активируют метаболизм, необходимы для роста и развития растений, дифференциации органов, ориентации по отношению к свету и силе тяжести. По химической природе — производные индола (см. ИНДОЛ) . Исследование эффектов ауксинов начато Ч. Дарвином (см. ДАРВИН Чарлз Роберт) и его сыном в 1880, продолжено П. Бойсен-Йенсеном (см. БОЙСЕН-ЙЕНСЕН Петер) (1913), А. Паалем (1919), Н. Г. Холодным (см. ХОЛОДНЫЙ Николай Григорьевич) и Ф. Вентом (1924—28) и др.
Основным ауксином является индолил-3-уксусная кислота (см. ИНДОЛИЛУКСУСНАЯ КИСЛОТА) (ИУК). Обнаружены и другие природные вещества с ауксиновой активностью, такие как индолил-3-ацетонитрил, 4-хлор-3-индолилуксусная и фенилуксусная кислоты, однако их ауксиновая активность существенно слабее. В растениях часто обнаруживают конъюгаты ИУК с аминокислотами, сахарами и спиртами, представляющие, как полагают, запасные формы ауксинов. Искусственно синтезирован ряд соединений с высокой ауксиновой активностью (2,4-дихлорфеноксиуксусная-, 2,4,5-трихлорфеноксиуксусная-, индолил-3-масляная-, 1-нафтилуксусная кислоты и другие), часто применяемых для научных и практических целей. ИУК синтезируется из триптофана в верхушках побега и перемещается сверху вниз по паренхимным клеткам со скоростью 10—15 мм/час благодаря особому механизму полярного транспорта. Возможно также более быстрое передвижение ауксинов по транспортным каналам растения. Ауксины обладают многообразным физиологическим действием и жизненно важны для роста и развития растений. Ауксины необходимы для деления и растяжения клеток, для формирования проводящих пучков и корней, способствуют разрастанию околоплодника. Ауксины обусловливают явление апикального доминирования, т. е. тормозящее действие апикальной почки на рост пазушных почек. Ауксины играют первостепенную роль в ростовых движениях: фото- и геотропизме и настиях. Ауксины усиливают аттрагирующее действие органов и тканей (т. е. их способность притягивать питательные вещества) и во многих случаях задерживают их старение. При реализации многих физиологических программ ауксины взаимодействуют с цитокининами и другими фитогормонами. При высоких концентрациях ауксины повышают образование своего антагониста — фитогормона этилена. Первичное действие ауксинов направлено на изменение активности (активацию или репрессию) определенного набора компетентных генов, характерного для данной ткани. ИУК также активирует АТФазу плазмалеммы, вызывая выкачивание протонов из клетки и закисление клеточной стенки. Это приводит к размягчению матрикса стенки, что делает возможным рост клеток растяжением. На практике ауксины и их синтетические аналоги применяют для размножения клеток и растений в стерильной культуре и для получения трансгенных растений (совместно с цитокининами). Их часто используют для предотвращения предуборочного опадения плодов древесных культур, получения бессемянных плодов томатов, огурцов, баклажанов, перца и др., а также как стимуляторы корнеобразования у черенков; в высоких дозах — как гербициды и дефолианты (2,4-дихлорфеноксиуксусная кислота и др.). Ауксины синтезируются многими фитопатогенными и симбиотическими микроорганизмами, что помогает последним воздействовать на клетки растения-хозяина.
Литературу см. при ст. Фитогормоны (см. ФИТОГОРМОНЫ) .
Энциклопедический словарь . 2009 .
Полезное
Смотреть что такое "ауксины" в других словарях:
АУКСИНЫ — группа гормонов растений. Регулируют на разных этапах жизни растения его рост, дифференцировку органов, ростовые реакции на свет и силу тяжести. По химической природе производные индола. Основной представитель индолилуксусная кислота … Большой Энциклопедический словарь
АУКСИНЫ — гормоны растений, производные индола. Образуются в апикальных меристемах и стимулируют клеточное растяжение. Один из наиболее распространённых А. Р индолил 3 уксусная к та (ИУК), или гетероауксин, биохимич. предшественником к рого является… … Биологический энциклопедический словарь
ауксины — группа гормонов растений. Регулируют на разных этапах жизни растения его рост, дифференцировку органов, ростовые реакции на свет и силу тяжести. По химической природе – производные индола. Образуются в апикальных меристемах. Синтезируются также… … Словарь микробиологии
Ауксины — * аўксіны * auxins растительные гормоны, которые индуцируют рост клеток и их деление, способствуют аттрагирующему действию тканей, активируют протонную помпу, регулируют рост растяжением и устьичные движения. Естественным А. является производное… … Генетика. Энциклопедический словарь
Ауксины — Индолил 3 уксусная кислота (гетероауксин) … Википедия
ауксины — ((гр. auxano увеличиваю; расту) группа вырабатываемых растениями фи тогормонов, стимулирующих ростовые процессы; в больших дозах действуют угнетающе; синтетически изготовленные а. (напр., гетероауксин) примен. в растениеводстве. Новый словарь… … Словарь иностранных слов русского языка
Ауксины — (от греч. аuхо выращиваю, увеличиваю) вырабатываемые в клетках растений вещества, стимулирующие ростовые процессы (рост корней у черенков, растяжение клеток у отрезков стеблей, деление клеток в культуре растительной ткани); группа… … Большая советская энциклопедия
АУКСИНЫ — (от греч. аихо выращиваю, увеличиваю), одна из групп прир. регуляторов роста растений ( фитогормонов). Влияют на рост, деление и дифференциацию клеток; играют важную роль в явлениях гео и фототропизма. Прир. А. производные индола, напр. 3 (3… … Химическая энциклопедия
АУКСИНЫ — группа гормонов р ний. Регулируют на разных этапах жизни р ния его рост, дифференцировку органов, ростовые реакции на свет и силу тяжести. По хим. природе производные индола. Осн. представитель индолилуксусная кислота … Естествознание. Энциклопедический словарь
Растения представлены самыми разнообразными по внешнему виду, форме и размерам организмами. Это могут быть как миниатюрные особи, вроде мохообразных, так и гиганты размером в 110 метров. Для координированного управления таким гигантским телом необходим постоянный обмен сигналами между различными его частями.
Для этих и многих других случаев у растений, как и у животных, есть специализированные системы передачи информации: электрическая, действующая локально путем изменения мембранных потенциалов клеток; трофическая, обусловленная потоками различных синтезированных внутри растения (эндогенных) веществ; и самая важная – гормональная, где ключевую роль играют сигнальные молекулы, которые называются фитогормонами. Фитогормоны (дословно с греческого – растительные возбудители) являются низкомолекулярными регуляторными эндогенными веществами, которые способны транспортироваться по растительному организму и вызывать различные тканеспецифичные ответы, но не участвующие во всевозможных метаболических путях непосредственно. Важным свойством всех гормонов является способность действовать в очень небольших концентрациях (примерно 1*10 -5 моль/л и меньше).
Действие фитогормонов заключается в изменении программы развития, что выражается комплексным изменением активности клеток и тканей: рост, растяжение, синтез пептидов и всевозможных метаболитов. Любая растительная клетка способна к синтезу всего разнообразия гормонов. Как и гормоны животных, растительные гормоны очень тесно взаимодействуют между собой. Небольшое изменение баланса в гормональной системе серьезно сказывается на различных уровнях организации тела растения.
Гормоны являются первичными мессенджерами в сигнальных путях. Они связываются с рецепторами на клеточной мембране или внутри клетки, и дальше по каскаду вторичных мессенджеров информация передается до конкретной эффекторной системы.
Очевидно, что светочувствительность и способность расти по направлению к свету (положительный фототропизм) является очень важной особенностью растений. Опыты по поиску светочувствительной части растений и были поставлены Чарльзом и Фрэнсисом Дарвинами (рис.1)
Деление клеток у растений происходит в особых тканях, называемых меристемами, одной из таких меристем побегов является апикальная (верхушечная) меристема. В некоторой области (субапикальной) под меристемой новообразованные клетки удлиняются и наблюдается рост побега. Было обнаружено, что если освещать проростки с одной стороны, то растения будут поворачиваться к источнику света, при этом изгиб будет формироваться как раз в зоне под меристемой.
В ходе самого опыта проростки канареечной травы освещали с одного бока и в результате наблюдали изгиб верхушки. Изгиб наблюдался и в случае, когда субапикальная зона была закрыта плотным материалом. А вот если закрыть макушки проростков, изгиба наблюдаться не будет. На основе данных эксперимента можно сделать вывод о том, что за световосприятие у растений ответственна меристема, и она, передавая какие-то сигналы нижележащей области, заставляет клетки в ней неравномерно удлиняться, что и обуславливает изгиб.
То, что этот сигнал является химическим веществом, было выявлено в опыте Ф.Вента. Похожие опыты независимо ставили Тиманн и Холодный.
В ходе этого эксперимента у растений отрезали меристему и переносили ее на агар. Из меристемы в агар выделялись разнообразные вещества, в том числе и интересующее исследователя. Затем эти агаровые блоки переносились на побеги с удаленной меристемой. И в том случае, когда использовали агаровый блок с веществами из меристемы, проростки начинали расти. Если же поместить блок только на край побега, то наблюдается изгиб, причем в сторону, противоположную блоку.
Вызывающее данный эффект химическое вещество, названное ауксином (от латинского слова расти), было выделено Ф.Кёглем из мочи вегетарианцев. Ауксины - это группа разнообразных химических веществ, основное из которых – гетероауксин или β-индолилуксусная кислота (ИУК), синтезируются в основном в апикальной меристеме побега и в растущих листьях, являются гормонами роста в небольших концентрациях, а также ингибиторами роста - в высоких. Последний эффект позволяет использовать синтетические аналоги ауксинов в качестве гербицидов. Применяемыми синтетическими аналогами часто являются 2,4-дифеноксиуксусная кислота, 1-натилуксусная кислота, индолмаслянная кислота. Эти вещества в растении могут превращаться в ИУК.
Есть несколько путей синтеза ИУК из триптофана (рис.4). В основном пути L-Trp под действием трансаминазы превращается в индол-2-пируват, который после декрабоксилирования и окисления превращается в ИУК. Интересно, что часть ИУК в растениях может иметь бактериальное происхождение. Такое наблюдается в случае заражения бактериями корончатого галла (Agrobacterium tumefaciens). В ходе заражения происходит трансформация растительных клеток плазмидой, несущей гены, отвечающие за неконтролируемый растениями синтез ИУК.
Инактивация и депонирование избытка гормона происходит путем взаимодействия со специфическими сахарами и белками. Распад идет путем окисления в присутствии молекулярного кислорода.
Ауксины по растению могут в незначительной степени транспортироваться по флоэме, основной же транспорт идет от клетки к клетке, что определяется наличием специальных трансмембранных переносчиков. Протонированная форма активно закачивается в клетку белком AUX1, депротонированная АУК выкачивается из клетки белком PIN. При этом AUX и PIN расположены на разных полюсах клетки, что обеспечивает полярный транспорт гормона. Это особенно важно при развитии и закладке разных частей растения. В присутствии ингибиторов полярного транспорта ауксина наблюдаются аномальные разрастания частей растения и изменение общего габитуса.
Ауксины имеют широкий спектр действия на растения, проявляющийся во множестве физиологических эффектов. Перечислим некоторые из них.
- Аттрагирующий эффект. Места синтеза и накопления ауксинов привлекают потоки питательных веществ, необходимых для деления и роста клеток.
- Растяжение клеток. Клеточная стенка растений содержит сшивочные гликаны (устаревшее: гемицеллюлоза). Эти полисахаридные молекулы сшивают между собой фибриллы целлюлозы, создавая таким образом жесткий каркас по типу матраса. Внутри клеток тонопласт создает тургорное давление, которое сдерживается жесткой структурой клеточной стенки. Во внеклеточном пространстве содержатся ферменты (экспансины, ксилоглюканэндотрансликозилазы) которые при закислении внеклеточной среды расщепляют связи между сшивочными гликанами, и фибриллы целлюлозы под действием давления расходятся.
Этот процесс вызывается ауксином. У растительной клетки обнаружено два рецептора связывания ИУК. Ранее считалось, что ключевым рецептором в данном случае может быть Auxin Binding Protein1 (ABP1), который располагается в клеточной мембране и мембране ЭПР. В последнее время было обнаружено, что ABP1 располагается в апопласте, и появились данные, оспаривающие необходимую роль ABP1 в процессах развития взрослого растения. Второй рецептор (а точнее - рецепторная система) внутриядерный. Это многокомпонентная систама, работающая по принципу ингибирования ингибирования - при связывании ауксина с белками-корецепторами (TIR1/AFB-Aux/IAA) происходит убиквитинилирование ингибитора генов ответа на ауксины (Aux/IAA) и его деградация в протеасоме. При этом активируется транскрипция генов ответа на ИУК. В частности, активируется транскрипция гена протонной помпы, которая встраивается в мембрану ЭПР и затем транспортируется в везикуле (как ранее полагали - усиленно за счет ABP1) к плазмалемме и встраивается в нее. Протонная помпа за счет гидролиза АТФ перекачивает протоны, закисляет апопласт, что приводит к активации в апопласте специальных белков (экспансины, ксилоглюканэндотрансгликозилазы), которые нарушают прочность и жесткость клеточной стенки, позволяя ей растянутся под воздействием внутриклеточного давления.
Ауксин влияет на дифференцировку клеток в растениях. Потоками ауксина размечаются места закладки прокамбия, т.е. проводящих пучков. Пучок формируется там, где поток ауксинов разворачивается внутрь меристемы.
Он также отвечает за закладку и развитие ксилемы. Под действием ауксинов происходит вторичное утолщение, пролиферация камбия.Ауксин влияет на дифференцировку клеток в растениях. Потоками ауксина размечаются места закладки прокамбия, т.е. проводящих пучков. Пучок формируется там, где поток ауксинов разворачивается внутрь меристемы.
- Ауксины отвечают за эффект апикального доминирования, заключающийся в том, что с потоками гормона боковые почки получают сигнал о благополучии апикальной и не растут. Чем дальше от верхушки, тем слабее сигнал, поэтому почки начинают расти, и наблюдается базитонное ветвление (рис.7Б). При удалении апекса побега (снятии эффекта апикального доминирования) первыми трогаются в рост почки, находящиеся выше всего, т.е. первые, до которых перестал доходить сигнал, это акротонное ветвление (А).
Эффект апикального доминирования также снимается при цветении. Этим можно объяснить существование соцветий типа дихазий.
5. Будучи ответственны за закладку проводящих пучков в меристеме, ауксины также определяют листорасположение. Согласно одной из гипотез, для закладки проводящего пучка необходимо определенное количество гормона, поступающего из клеток меристемы. Таким образом, существует некая фиксированная площадь, под которой потом будут закладываться проводящие ткани. Ближе, чем позволяют границы этой площади, соседний проводящий пучок не сможет возникнуть. А, поскольку каждый проводящий пучок принадлежит определенному листу, расположение примордиев (зачатков) листа будет соответствовать расположению этой площади. В зависимости от индивидуальных характеристик растения, от размеров его меристемы (количества этих площадей в одном круге) будет зависеть филлотаксис.
6. Увеличение листовой массы растения вызывает увеличение площади транспирации, что должно сопровождаться более интенсивным поглощением почвенных растворов. Соответственно, ауксины стимулируют ризогенез, заключающийся в закладке боковых и придаточных корней. На этом основано действие сельскохозяйственных препаратов типа Корневина.
Индол-3-уксусная кислота (ИУК) является наиболее распространенным и основным ауксином, который естественным образом встречается и функционирует в растениях. Он вызывает большинство эффектов ауксина у интактных растений и является наиболее сильным нативным ауксином.
В растениях есть еще четыре эндогенно синтезируемых ауксина. [1] [2]
Все ауксины представляют собой соединения с ароматическим кольцом и группой карбоновой кислоты: [2] [3]
Ауксины (множественное число от ауксина / ˈ ɔː k s ɪ п / ) являются классом гормоны растений (или регуляторы роста растений) с некоторыми морфоген-подобные характеристики. Ауксины играют кардинальную роль в координации многих процессов роста и поведения в жизненных циклах растений и необходимы для развития организма растений. Голландский биолог Фриц Вармолт пошел впервые описал ауксины и их роль в росте растений в 1920-х годах. [4] Кеннет В. Тиманн (1904-1997) стал первым, кто выделил один из этих фитогормоны и определить его химическую структуру как индол-3-уксусная кислота (IAA). Вент и Тиманн написали в соавторстве книгу о растительных гормонах. Фитогормоны, в 1937 г.
Содержание
Обзор
(Динамический и адаптивный к окружающей среде) шаблон Распределение ауксина в растении является ключевым фактором для роста растения, его реакции на окружающую среду и, в частности, для развития органов растения. [5] [6] (Такие как листья или же цветы). Это достигается за счет очень сложных и слаженных активный транспорт молекул ауксина от клетки к клетке по всему растительному организму - так называемым полярный транспорт ауксина. [5] Таким образом, растение может (в целом) реагировать на внешние условия и приспосабливаться к ним, не требуя нервная система. Ауксины обычно действуют вместе с другими растительными гормонами или против них. Например, отношение ауксина к цитокинин в определенных тканях растений определяет зарождение корневых почек, а не побегов.
На молекулярном уровне все ауксины представляют собой соединения с ароматическим кольцом и карбоновая кислота группа. [3] Самый важный член семейства ауксинов - это индол-3-уксусная кислота (IAA), [7] который генерирует большинство эффектов ауксина в интактных растениях и является наиболее мощным нативным ауксином. А как нативный ауксин, его равновесие у растений контролируется разными способами, от синтеза до возможных спряжение на разложение его молекул, всегда в соответствии с требованиями ситуации.
- Пять природных (эндогенных) ауксинов в растениях включают индол-3-уксусную кислоту, 4-хлориндол-3-уксусная кислота, фенилуксусная кислота, индол-3-масляная кислота, и индол-3-пропионовая кислота. [1][2] Однако большая часть знаний, описанных до сих пор в биологии ауксинов и описанных в следующих разделах, в основном применима к ИУК; остальные три эндогенных ауксина, по-видимому, имеют незначительное значение для интактных растений в естественной среде. Помимо эндогенных ауксинов, ученые и производители разработали множество синтетических соединений с ауксиновой активностью.
- Синтетические аналоги ауксина включают: 1-нафталинуксусная кислота, 2,4-дихлорфеноксиуксусная кислота (2,4-Д), [1] и много других.
Некоторые синтетические ауксины, такие как 2,4-Д и 2,4,5-трихлорфеноксиуксусная кислота (2,4,5-Т), продаются как гербициды. Широколиственные растения (двудольные), Такие как одуванчики, гораздо более восприимчивы к ауксинам, чем узколистные растения (однодольные) Такие как травы и каша сельскохозяйственных культур, что делает эти синтетические ауксины ценными гербицидами.
Открытие ауксина
Чарльз Дарвин
В 1881 г. Чарльз Дарвин и его сын Фрэнсис проводил эксперименты на колеоптили- влагалища, закрывающие молодые листья у прорастающих проростков травы. Эксперимент выставил колеоптиль на свет от однонаправленного источника и обнаружил, что они изгибаются навстречу свету. [8] Накрыв различные части колеоптилей светонепроницаемой непрозрачной крышкой, Дарвины обнаружили, что свет улавливается концом колеоптиля, но изгиб происходит в гипокотиль. Однако проростки не проявляли признаков развития к свету, если верхушка была закрыта непрозрачной крышкой или если верхушка была удалена. Дарвины пришли к выводу, что кончик колеоптиля отвечает за восприятие света, и предположили, что посланник передается вниз от кончика колеоптиля, заставляя его изгибаться. [9]
Питер Бойзен-Йенсен
В 1913 году датский ученый Питер Бойзен-Йенсен продемонстрировал, что сигнал не является фиксированным, а подвижным. Он отделил кончик от остальной части колеоптиля кубиком желатина, который предотвращал клеточный контакт, но позволял химическим веществам проходить через него. Сеянцы ответили нормально, наклонившись к свету. Однако, когда наконечник был отделен непроницаемым веществом, кривизны стержня не было. [10]
Фриц пошел
В 1928 году голландский ботаник Фриц Вармолт пошел показали, что химический посланник распространяется от кончиков колеоптилей. Эксперимент Вента установил, как химическое вещество, способствующее росту, заставляет колеоптиль расти к свету. Пошел срезал кончики колеоптилей и поместил их в темноту, положив несколько кончиков на блоки агара, которые, как он предсказывал, поглотят химическое вещество, стимулирующее рост. На контрольных колеоптилях он поместил блок, в котором не было химического вещества. На других он разместил блоки, содержащие химикат, либо по центру на вершине колеоптиля, чтобы распределить химикат равномерно, либо смещенными, чтобы увеличить концентрацию с одной стороны. [9]
Когда химическое вещество, способствующее росту, было распределено равномерно, колеоптиль рос прямо. Если химическое вещество распределялось неравномерно, колеоптиль изгибался в сторону от куба, как будто рос к свету, даже если он был выращен в темноте. Позже Вент предположил, что субстанция-посредник представляет собой гормон, способствующий росту, который он назвал ауксином, который асимметрично распределяется в области изгиба. Вент пришел к выводу, что ауксин находится в более высокой концентрации на затемненной стороне, способствуя удлинению клеток, что приводит к изгибу колеоптилей в направлении света. [10]
Гормональная активность
Ауксины помогают развитию растений на всех уровнях, начиная с сотовый уровне, через органы и, в конечном итоге, на все растение.
Молекулярные механизмы
Когда растительная клетка вступает в контакт с ауксином, она вызывает резкие изменения экспрессия гена, со многими генами с повышенной или пониженной регуляцией. Точные механизмы, с помощью которых это происходит, все еще являются областью активных исследований, но в настоящее время существует общий консенсус по крайней мере по двум сигнальным путям ауксина. [11] [12]
Восприятие
Другой ауксин-связывающий белок, ABP1, в настоящее время часто рассматривается как рецептор ауксина ( апопласт), но обычно считается, что он играет гораздо более незначительную роль, чем путь передачи сигналов TIR1 / AFB, и о передаче сигналов ABP1 известно гораздо меньше. [12]
Модули сигнализации Aux / IAA и ARF
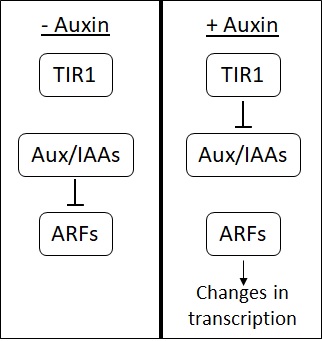
Факторы ответа на ауксин (ARF) представляют собой большую группу факторы транскрипции которые действуют в передаче сигналов ауксина. В отсутствие ауксина ARF связываются с классом репрессоров, известным как Aux / IAA. Aux / IAA подавляют способность ARF усиливать транскрипцию генов. Кроме того, связывание Aux / IAA с ARF приводит Aux / IAA в контакт с промоутеры генов, регулируемых ауксином. Находясь на этих промоторах, Aux / IAA подавляет экспрессию этих генов за счет привлечения других факторов для внесения изменений в ДНК структура.
Связывание ауксина с TIR1 / AFB позволяет им связываться с Aux / IAA. Когда они связаны TIR1 / AFB, Aux / IAAs помечаются для деградации. Распад Aux / IAA высвобождает белки ARF, которые затем могут активировать или репрессировать гены, с промоторами которых они связаны. [11] [12]
Считается, что большое количество возможных пар связывания Aux / IAA и ARF и их различное распределение между типами клеток и возрастом развития объясняют удивительно разнообразные ответы, которые производит ауксин.
В июне 2018 года было продемонстрировано, что ткани растений могут чрезвычайно быстро реагировать на ауксин TIR1-зависимым образом (вероятно, слишком быстро, чтобы это можно было объяснить изменениями в экспрессии генов). Это заставило некоторых ученых предположить, что существует еще не идентифицированный TIR1-зависимый путь передачи сигналов ауксина, который отличается от хорошо известного транскрипционного ответа. [13]
На клеточном уровне
На клеточном уровне ауксин необходим для рост клеток, затрагивая как деление клеток и клеточная экспансия. Уровень концентрации ауксина вместе с другими местными факторами способствует дифференциация клеток и уточнение клеточной судьбы.
В зависимости от конкретной ткани ауксин может способствовать осевому удлинению (как в побегах), латеральному расширению (как при набухании корней) или изодиметрическому расширению (как при росте плодов). В некоторых случаях (рост колеоптилей) клеточная экспансия, стимулируемая ауксином, происходит в отсутствие клеточного деления. В других случаях клеточное деление, стимулируемое ауксином, и рост клеток могут быть точно секвенированы в пределах одной и той же ткани (зарождение корня, рост плода). В живом растении ауксины и другие гормоны растений почти всегда взаимодействуют, определяя паттерны развития растения.
Узоры органов
Рост и деление растительных клеток вместе приводят к росту ткань, а специфический рост тканей способствует развитию растений органы.
Рост клеток влияет на размер растения, неравномерно локализованный рост вызывает изгиб, поворот и направление органов - например, стебли, поворачивающиеся к источникам света (фототропизм), корни, растущие под действием силы тяжести (гравитропизм), и другие тропизмы возникло потому, что клетки с одной стороны растут быстрее, чем клетки с другой стороны органа. Таким образом, точный контроль распределения ауксина между различными клетками имеет первостепенное значение для конечной формы роста и организации растений.
Транспорт ауксина и неравномерное распределение ауксина
Чтобы вызвать рост в необходимых доменах, ауксины обязательно должны быть активны преимущественно в них. Локальные максимумы ауксина могут быть образованы активным биосинтезом в определенных клетках тканей, например, через триптофан-зависимые пути, [14] но ауксины синтезируются не во всех клетках (даже если клетки сохраняют потенциальную способность делать это, синтез ауксина в них активируется только при определенных условиях). Для этой цели ауксины должны быть не только перемещены в те участки, где они необходимы, но также должны иметь установленный механизм для обнаружения этих участков. Транслокация осуществляется по всему телу растения, в основном из пики всходов к вершинам корней (сверху вниз).
На большие расстояния перемещение происходит через поток жидкости в флоэма сосудов, но для транспортировки на короткие расстояния используется уникальная система скоординированного полярного транспорта непосредственно от клетки к клетке. Этот активный транспорт на короткие расстояния демонстрирует некоторые морфогенетический характеристики.
Этот процесс, полярный транспорт ауксина, является направленным, очень строго регулируемым и основан на неравномерном распределении переносчиков оттока ауксина на плазматической мембране, которые направляют ауксины в нужном направлении. Хотя белки в форме PIN (PIN) жизненно важны для полярного транспорта ауксина, [6] [15] семейство генов AUXIN1 / LIKE-AUX1 (AUX / LAX) кодирует неполярных носителей притока ауксина. [16]
Регуляция локализации белка PIN в клетке определяет направление транспорта ауксина из клетки, и концентрированное усилие многих клеток создает пики ауксина или максимумы ауксина (области, в которых клетки с более высоким содержанием ауксина - максимум). [6] Правильные и своевременные максимумы ауксина в развивающихся корнях и побегах необходимы для организации развития органа. [5] [17] [18] PIN регулируются несколькими путями как на транскрипционном, так и на посттрансляционном уровнях. Белки PIN могут фосфорилироваться с помощью PINOID, который определяет их апикобазальную полярность и тем самым направленность потоков ауксина. Кроме того, другие киназы AGC, такие как D6PK, фосфорилируют и активируют переносчики PIN. Киназы AGC, включая PINOID и D6PK, нацелены на плазматическую мембрану посредством связывания с фосфолипидами. Перед D6PK 3'-фосфоинозитид-зависимая протеинкиназа 1 (PDK1) действует как главный регулятор. PDK1 фосфорилирует и активирует D6PK на базальной стороне плазматической мембраны, выполняя активность PIN-опосредованного полярного транспорта ауксина и последующего развития растений. [19] Вокруг максимума ауксина находятся клетки с минимумом ауксина или минимумом ауксина. Например, в Арабидопсис плода, было показано, что минимальное количество ауксина важно для развития его тканей. [20]
Ауксин оказывает значительное влияние на пространственную и временную экспрессию генов во время роста апикальных меристем. Эти взаимодействия зависят как от концентрации ауксина, так и от пространственной ориентации во время первичного позиционирования. Ауксин полагается на PIN1, который работает как переносчик оттока ауксина. Расположение PIN1 на мембранах определяет направленный поток гормона от более высоких концентраций к более низким. [21] Зарождение зачатков в апикальных меристемах коррелирует с повышенными уровнями ауксина. [22] Гены, необходимые для определения идентичности клеток, располагаются и экспрессируются в зависимости от уровней ауксина. STM (SHOOTMERISTEMLESS), который помогает поддерживать недифференцированные клетки, подавляется в присутствии ауксина. Это позволяет растущим клеткам дифференцироваться в различные ткани растений. Гены CUC (CUP-SHAPED COTYLEDON) устанавливают границы для роста тканей и способствуют росту. [23] Они активируются притоком ауксина. [24] Эксперименты с использованием визуализации GFP (GREEN FLUORESCENCE PROTEIN) на Arabidopsis подтвердили эти утверждения.
Организация завода
Важный принцип организации растений, основанный на распределении ауксина: апикальное доминирование, что означает, что ауксин, продуцируемый апикальной почкой (или верхушкой роста), диффундирует (и транспортируется) вниз и подавляет развитие роста задней боковой почки, которая в противном случае конкурировала бы с верхушкой за свет и питательные вещества. Удаление верхушечного кончика и его подавляющего действия ауксина позволяет нижним спящим боковым почкам развиваться, а почки между стеблем листа и стеблем дают новые побеги, которые конкурируют за лидерство в росте. Процесс на самом деле довольно сложен, потому что ауксин, транспортируемый вниз от верхушки свинцового побега, должен взаимодействовать с несколькими другими гормонами растения (такими как стриголактоны или же цитокинины) в процессе в различных положениях вдоль оси роста в теле растения для достижения этого явления. Такое поведение растений используется в обрезка садоводами.
Наконец, количество ауксина, поступающего от стеблей к корням, влияет на степень роста корней. Если удалить кончики побегов, растение не отреагирует только на рост боковых почек, которые, как предполагается, заменят исходный свинец. Отсюда также следует, что меньшее количество ауксина, поступающего к корням, приводит к более медленному росту корней, и, следовательно, питательные вещества в большей степени вкладываются в верхнюю часть растения, которая, следовательно, начинает расти быстрее.
Читайте также: