
Анизогамия это в биологии кратко
Обновлено: 02.07.2024
Связанные понятия
Оогамия (от др.-греч. ᾠόν — яйцо и γάμος — брак) — это вид полового процесса, при котором сливаются резко отличающиеся друг от друга половые клетки — крупная неподвижная яйцеклетка с мелкой, обычно подвижной, мужской половой клеткой (сперматозоидом или спермием).
Тип спа́ривания, или полово́й тип, — понятие, применяемое по отношению к микроорганизмам, у которых есть половой процесс (конъюгация, изогамия или анизогамия), но к которым классическое понятие биологического пола неприменимо, так как у них отсутствуют яйцеклетки и сперматозоиды как таковые (то есть гаметы, различные структурно и по размеру). Вместо этого у них может происходить передача генетической информации между клетками при их непосредственном контакте, но не сопровождающаяся слиянием клеток.
Гаметогенез или предзародышевое развитие — процесс образования половых клеток, или гамет. Поскольку в ходе гаметогенеза специализация яйцеклеток и сперматозоидов происходит в разных направлениях, обычно выделяют оогенез (устаревшее название — овогенез в современных изданиях по эмбриологии больше не употребляется) и сперматогенез.
Зооспо́ра (др.-греч. ζῷον — животное и σπορά — посев, семя), или зоогони́дий, или бродя́жка — стадия жизненного цикла многих водорослей и некоторых низших грибов. Представляют собой жгутиконосцев, перемещающихся в жидкой среде с помощью биения одного или нескольких жгутиков. Многие водоросли на этой стадии обладают хроматофором, стигмой и сократительными вакуолями. Зооспоры некоторых желто-зелёных водорослей обладают многочисленными ядрами и несколькими парами жгутиков (синзооспоры).
Гаметы, или половые клетки, — репродуктивные клетки, имеющие гаплоидный (одинарный) набор хромосом и участвующие, в частности, в половом размножении. При слиянии двух гамет в половом процессе образуется зигота, развивающаяся в особь (или группу особей) с наследственными признаками обоих родительских организмов, произведших гаметы.
Гетерогаме́тный пол — это генетически определённый пол, соответствующий наличию в клетках организма двух разных половых хромосом или одной, в удвоенной дозе приводящей к формированию альтернативного пола. Особи гетерогаметного пола дают две группы гамет (по содержанию разных половых хромосом).
Полово́й проце́сс, или оплодотворе́ние, или амфими́ксис (др.-греч. ἀμφι- — приставка со значением обоюдности, двойственности и μῖξις — смешение), или сингамия — процесс слияния гаплоидных половых клеток, или гамет, приводящий к образованию диплоидной клетки зиготы. Не следует смешивать это понятие с половым актом (встречей половых партнёров у многоклеточных животных).
Половое размножение — процесс у большинства эукариот, связанный с развитием новых организмов из половых клеток (у одноклеточных эукариот при конъюгации функции половых клеток выполняют половые ядра).
Конъюга́ция у инфузо́рий — половой процесс инфузорий, сопровождающийся переносом ядер между клетками партнёров при их непосредственном контакте. Наличие такого своеобразного полового процесса является уникальной чертой инфузорий. Половой процесс у инфузорий, в отличие от полового процесса в привычном представлении, не сопровождается образованием гамет, поэтому у них нет и зиготы. Кроме того, конъюгация инфузорий не сопровождается размножением, то есть увеличением числа клеток, поэтому конъюгация.
Оогене́з или овогене́з (др.-греч. ᾠόν — яйцо + γένεσις — возникновение) — развитие женской половой клетки — яйцеклетки (яйца). Одна из форм гаметогенеза.
Почкование — тип бесполого или вегетативного размножения животных, грибов и растений, при котором дочерние особи формируются из выростов тела материнского организма (почек). Почкование характерно для многих грибов, печёночных мхов и животных (простейшие, губки, кишечнополостные, некоторые черви, оболочники, некоторые жгутиковые, инфузории, споровики). У ряда животных — почкование не доходит до конца, молодые особи остаются соединёнными с материнским организмом. В ряде случаев это приводит к образованию.
Бесполое размножение, или агамогенез — один из способов размножения, при котором следующее поколение развивается из соматических клеток без участия репродуктивных клеток — гамет. Следует отличать бесполое размножение от однополого размножения (партеногенеза), который является особой формой полового размножения.
Метагене́з — форма жизненного цикла животных, состоящая в закономерном чередовании бесполых поколений и поколений, размножающихся половым путём.
Парасексуа́льный проце́сс, или парасексуа́льный цикл, — аналог полового процесса, свойственный грибам и одноклеточным организмам. При парасексуальном процессе происходит слияние вегетативных клеток, содержащих генетически разнородные ядра (образование гетерокариона), возникновение гетерозиготных диплоидов за счёт слияния ядер в гетерокарионах и последующего митотического расщепления диплоидов, приводящее к появлению гаплоидных или диплоидных рекомбинантов.
Дициеми́ды (лат. Dicyemida) — тип примитивно устроенных беспозвоночных животных, паразитирующих на головоногих моллюсках. Содержит 70—75 видов. Ранее дициемид из-за простоты организации объединяли с ортонектидами в группу Mesozoa, рассматривая в качестве переходной группы между протистами и многоклеточными животными. Впоследствии, когда получила широкую поддержку гипотеза об их вторичном упрощении в связи с паразитическим образом жизни, эти представления были отвергнуты.
Эмбриогенез — это физиологический процесс, в ходе которого происходит образование и развитие эмбриона. Эмбриогенез — это начальная стадия онтогенеза живых существ. Хотя явление эмбриогенеза известно как в растительном, так и в животном царствах, а также в царстве многоклеточных грибов, данная статья посвящена общим особенностям эмбриогенеза только у животных, с некоторым акцентом на особенности эмбриогенеза хордовых и позвоночных, в особенности млекопитающих.
Дробле́ние — ряд последовательных митотических делений оплодотворенного или инициированного к развитию яйца. Дробление представляет собой первый период эмбрионального развития, который присутствует в онтогенезе всех многоклеточных животных. При этом масса зародыша и его объём не меняются, оставаясь такими же, как и в начале дробления. Яйцо разделяется на все более мелкие клетки — бластомеры. Характерная особенность дробления — ведущая регуляторная роль цитоплазмы в развитии. Характер дробления зависит.
Бластоме́ры — клетки эмбрионов животных на этапе дробления зиготы. Оплодотворенная яйцеклетка (зигота) делится на две дочерние клетки, называемые бластомерами. Каждый бластомер делится на два новых дочерних бластомера. У большинства животных первые деления эмбриональных клеток не сопровождаются их ростом — каждое новое поколение клеток приблизительно в два раза меньше по размеру. В результате таких делений, количество клеток в зародыше увеличивается, но общий размер зародыша остается прежним. Этот.
Гаметангий — орган полового размножения у растений, в котором развиваются половые клетки. У водорослей и грибов гаметангием называют клетку, в которой образуются подвижные половые элементы — гаметы.
Са́ркома́стигофо́ры, или Саркожгутиконосцы (лат. Sarcomastigophora) — от греч. sarcodes — мясистый — полифилетическая группа в некоторых старых системах рассматривалась в качестве типа свободноживущих и паразитических простейших, которые передвигаются с помощью особых вре́менных выростов цитоплазмы (псевдоподий) или бичевидных выростов (жгутиков). Насчитывают около 18000 видов.
Хламидомона́да (лат. Chlamydómonas) — род одноклеточных зелёных водорослей из семейства Хламидомонадовые (Chlamydomonadaceae).
Гоноци́т (лат. gonocytus; гоно- + гист. cytus клетка) или первичная половая клетка — эмбриональная клетка, из которой впоследствии могут образоваться сперматозоиды или яйцеклетки. Также гоноцитом могут называться любые клетки, участвующие в процессе гаметогенеза, и сами гаметы.
Зиго́та (от др.-греч. ζυγωτός — удвоенный) — диплоидная (содержащая полный двойной набор хромосом) клетка, образующаяся в результате оплодотворения (слияния яйцеклетки и сперматозоида). Зигота является тотипотентной клеткой, то есть способной породить любую другую. Термин ввёл немецкий ботаник Э. Страсбургер.
Сократи́тельная вакуо́ль — мембранный органоид, осуществляющий выброс излишков жидкости из цитоплазмы. Представляет собой наиболее заметную часть согласованно работающего комплекса, в котором выступает в роли периодически опорожняющегося резервуара. Жидкость поступает в сократительную вакуоль из системы пузыревидных или трубчатых вакуолей, называемой спонгио́м. Работа комплекса позволяет поддерживать более или менее постоянный объём клетки, компенсируя постоянный приток воды через плазматическую.
Во́львокс (лат. Volvox) — род подвижных колониальных организмов, относящийся к отделу зелёных водорослей. Обитают в стоячих пресных водоёмах. При массовом размножении вызывают цветение воды, окрашивая её в зелёный цвет.
Определе́ние по́ла, или детермина́ция по́ла, — биологический процесс, в ходе которого развиваются половые характеристики организма. Большинство видов организмов имеет два пола. Иногда встречаются также гермафродиты, сочетающие признаки обоих полов. Некоторые виды имеют лишь один пол и представляют собой самок, размножающихся без оплодотворения путём партеногенеза, в ходе которого на свет появляются также исключительно самки.
Половые хромосомы, или гоносомы, — хромосомы, набор которых отличает мужские и женские особи у животных и растений с хромосомным определением пола.
Ортонекти́ды (лат. Orthonectida) — тип многоклеточных животных, который ранее вместе с типом дициемид объединяли в тип мезозои. Все ортонектиды — эндопаразиты беспозвоночных животных. Ортонектиды характеризуются довольно сложным жизненным циклом, включающим стадии плазмодия, половых особей и личинок, и крайне простой организацией половых особей, в которой, впрочем, сохраняются основные черты организации двусторонне-симметричных первичноротых животных. Тип включает 2 семейства, содержащих 6 родов.
Гетерого́ния (от др.-греч. ἕτερος — другой и γονή — поколение) — в биологии вид размножения, когда у одного и того же вида животных чередуются между собой два различных половых поколения, один из видов чередования поколений, в отличие от чередования половых поколений с бесполыми (метагенезис).
Бластоцель (муж. род, лат. blastocoelia; от др.-греч. βλαστός — зачаток, зародыш и κοῖλος — полый; Полость дробления) — полость бластулы, образующаяся между бластомерами у зародышей животных. Заполнена жидкостью, отличающейся по химическому составу от окружающей среды. Полость увеличивает площадь поверхности эмбриона, улучшая его способность усваивать питательные вещества и кислород. Достигает наибольшего размера к концу дробления, на стадии бластулы. В процессе гаструляции постепенно вытесняется.
Апикомпле́ксы (лат. Apicomplexa), или споровики́ (лат. Sporozoa) — тип простейших из группы альвеолят (лат. Alveolata). Все представители типа являются облигатными паразитами позвоночных и беспозвоночных животных. Общность плана строения апикомплекс наиболее отчетливо проявляется на стадии зоита и выражается в наличии специфического комплекса органелл — апикального комплекса. Покровы представлены характерной для альвеолят пелликулой. В жизненном цикле большинства представителей типа обнаружен половой.
Гомологи́чные хромосо́мы — пара хромосом приблизительно равной длины, с одинаковым положением центромеры и дающие одинаковую картину при окрашивании. Их гены в соответствующих (идентичных) локусах представляют собой аллельные гены — аллели, то есть кодируют одни и те же белки или РНК. При двуполом размножении одна гомологичная хромосома наследуется организмом от матери, а другая — от отца.
Хоанофлагеллаты или воротничковые жгутиконосцы (лат. Choanoflagellata) — тип одноклеточных эукариот (протистов). Характерная черта — наличие у клетки одного жгутика, окружённого воротничком из 30—40 микроворсинок. Хоанофлагеллаты представлены одиночными и колониальными формами, обитающими в морских и пресных водах. Описано около 150 видов.
Грегарины (лат. Gregarinasina) — подкласс одноклеточных паразитических протистов из типа апикомплексов (споровиков) надтипа альвеолят. Большинство представителей — внеклеточные паразиты практически всех известных на сегодняшний момент типов беспозвоночных (членистоногих, кольчатых червей, иглокожих и т. д.).
Псевдопо́дии, или ложноно́жки, — цитоплазматические выросты у одноклеточных организмов и некоторых видов клеток многоклеточных. Используются клетками для передвижения (амёбоидное движение) и ловли крупных частиц (например, пищи или материала для строительства раковины). В зависимости от особенностей строения (например, организации цитоскелета) псевдоподии разделяют на лобоподии, филоподии, аксоподии и ретикулоподии.
Плодовая мушка Drosophila melanogaster была введена в качестве модельного организма в генетические эксперименты Томасом Морганом в 1909 году и до настоящего времени является одним из самых любимых модельных организмов среди исследователей, изучающих эмбриональное развитие животных. Малый размер, быстрая смена поколений, высокая плодовитость, прозрачность эмбрионов — делают дрозофилу идеальным объектом для генетических исследований.
Сти́гма (глазо́к) — внутриклеточный органоид, имеющийся у многих протистов, в том числе одноклеточных жгутиковых водорослей. Наличие стигмы обусловливает способность организма к фототаксису (однако фототаксисом обладают и многие протисты, лишённые стигмы, например, многие виды инфузорий). Стигма обычно включает пятно ярко-красного цвета, представляющее собой скопление глобул, содержащих пигмент гематохром. Строение стигмы у разных организмов различно: в большинстве случаев у водорослей они являются.
Хитридиомице́ты (лат. Chytridiomycota) — отдел царства грибов (Fungi). Объединяет более 120 родов и около 1000 видов. Мицелий слабо развит, основная масса таллома представляет собой т. н. плазмодий, из которого вырастают ризоидные гифы. Самые примитивные представители совершенно не имеют мицелия, и тело их в вегетативном состоянии представлено одиночной клеткой, иногда лишённой жёсткой клеточной стенки. Основа клеточной стенки хитиново-глюкановая, как и у высших грибов.
Фототаксис (от др.-греч. φως / φωτος — свет и τάξις — строй, порядок, расположение по порядку) — тип таксиса, свойство клеток и микроорганизмов ориентироваться и двигаться по направлению к или от источника света, характерное прежде всего фототрофным организмам. Существует как положительный фототаксис (движение происходит в направлении к источнику света; осуществляется за счёт светочувствительных глазок и жгутиков), так и отрицательный фототаксис — движение идёт в противоположном направлении.
Пиреноид — особое включение (дифференцированная область) внутри хлоропласта. Встречаются у некоторых групп эукариотических водорослей и у антоцеротовых мхов. Функционально представляет собой место запасания фермента рибулозобисфосфаткарбоксилаза (рубиско) и/или центр синтеза сахаров при помощи этого фермента. Вокруг пиреноида у зелёных водорослей откладываются запасные продукты (крахмал).
Диминуция хроматина (от лат. diminutio — уменьшение) — общее название клеточных генетических процессов, в ходе которых при эмбриональном развитии некоторых многоклеточных животных (в основном, беспозвоночных) соматические клетки запрограммировано теряют часть генетического материала, присутствовавшего в зиготе и остающегося нетронутым в клетках зародышевой линии. Диминуция хроматина наблюдается у некоторых представителей двукрылых насекомых, паразитических круглых червей (нематод), веслоногих ракообразных.
Полярное тельце (син.: направительное тельце, от англ. polar body) - образуется в процессе овогенеза в результате первого и второго мейотического деления. Полярное тело имеет гаплоидный набор хромосом. Используется в технологии искусственного оплодотворения (ЭКО), как материал для анализа потенциального качества яйцеклетки.
Бла́стула (зародышевый пузырь, бластосфера) — это многоклеточный зародыш, имеющий однослойное строение (один слой клеток), стадия в развитии зародыша, которую проходят яйца большинства животных — окончательный результат процесса дробления яйца. При своем дроблении (см. Дробление яйца) яйцо рядом последовательных делений распадается на комплекс клеток, именуемых шарами дробления, сегментационными шарами, или бластомерами, относительная величина и взаимное расположение которых бывают различны, смотря.
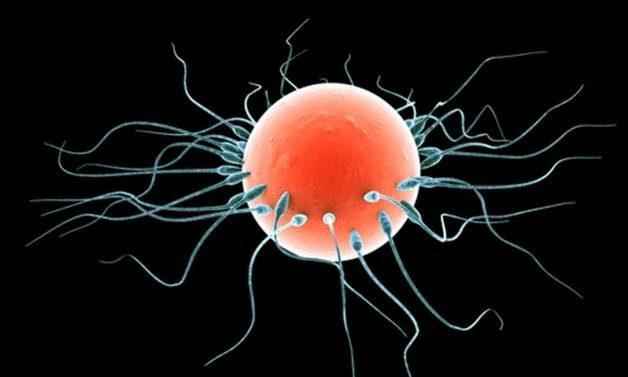
Анизогамия, также называемая гетерогамией является формой полового размножения, включающей объединение или слияние двух разнородных гамет (отличающихся по размеру или форме). Меньшие гаметы считаются мужским (сперматозоиды), тогда как более крупные гаметы являются женскими (яйцеклетки).
Существует несколько видов анизогамии. Обе гаметы могут иметь жгутики и, следовательно, подвижны. Также может быть, что обе гаметы не подвижны. Такая ситуация возникает, например, у некоторых водорослей и растений. У красных водорослей полисифония (Polysiphonia) большие неподвижные яйцеклетки оплодотворяются маленькими неподвижными сперматозоидами. У цветущих растений, гаметы являются не подвижными половыми клетками в гаметофитах.
Один из видов гетерогамии, который встречается у животных, называется оогамия. При оогамии большая и неподвижная яйцеклетка оплодотворяется маленьким подвижным сперматозоидом. Крупная яйцеклетка оптимизирована для долголетия, а небольшие сперматозоиды довольно подвижны и быстры. Размер и ресурсы яйцеклетки позволяют производить феромоны, которые привлекают сперматозоидов.
Эволюция анизогамии
Анизогамия – это процесс оплодотворение мелкими мужскими гаметами (сперматозоиды) крупных женских гамет (яйцеклетки). Разница в размере гамет является принципиальным различием между самцами (мужчинами) и самками (женщинами). Гетерогамия впервые развилась у многоклеточных гаплоидных видов, когда была установлена дифференциация различных типов спаривания.
Для эволюции анизогамии были предложены две теории. Основное внимание уделяется теории конкуренции спермы; другой акцент делается на взаимодействие различных типов спаривания для борьбы с рисками оплодотворения. Обе теории предполагают, что анизогамия возникла из-за дизруптивного (разрывающего) отбора, действующего на предковую изогамную популяцию, и что существует компромисс между большим числом гамет и меньшим приспособлением каждой отдельной гаметы, поскольку общий ресурс, который один организм может вложить в воспроизведение фиксируется.
Теория конкуренции спермы была первой теорией, объясняющей эволюцию анизогамии, предложенную Джеффом Паркером. Он предположил, что, поскольку размер зиготы определяется размерами обоих гамет, то меньший размер гаметы, уменьшит размер зиготы не менее чем на половину. Поэтому существует большая выгода для уменьшения размера гамета, чтобы значительно увеличить их количество.
Однако конкуренция существует не только среди спермы, но также и среди яйцеклеток. Паркер не дал объяснений, почему именно сперма стала меньше, а не яйцеклетки, или что привело к тому, что прото-сперма и прото-яичники стали неравными. Фактически, недавняя математическая модель показывает, что конкуренция сперматозоидов не является ни необходимой, ни достаточной для эволюции анизогамии. Конкуренция сперматозоидов приводила только к тому, что сперматозоиды становились все меньше и меньше.
Предполагается, что реальной причиной эволюции анизогамии являются риски оплодотворения. Важной проблемой для размножения многоклеточных организмов является перенос гамет с места на место для оплодотворения. Однако транспортировка имеет вероятность отказа и создает риски оплодотворения. Когда риск возрастает, гаметы одного типа спаривания должны уменьшаться, чтобы увеличить число для преодоления этого риска, а гаметы другого типа спаривания должны стать больше, чтобы компенсировать потери питательных веществ у зигот.
Таким образом, риски оплодотворения и взаимодействие различных типов спаривания могли вызвать эволюцию анизогамии. Сперма развивается, чтобы увеличить вероятность оплодотворения, а яйцеклетка развиваются, чтобы компенсировать потерю питательных веществ. Кроме того, сперматозоиды могут становится еще меньше, если яйцеклетки станут более крупными, чтобы обеспечить питательными веществами зиготы. Это взаимодействие может служить причиной того, что многие организмы имеют микроскопических сперматозоидов и чрезвычайно крупные яйцеклетки.
Существует несколько видов изогамии. Обе половые клетки могут быть жгутиконосцами и, следовательно, подвижны. Этот тип встречается, например, у водорослей, таких как некоторые, но не все виды хламидомонада (Chlamydomonas).
С неподвижными клетками
Конъюгация
Другая, более сложная форма — это конъюгация (подобно обмену генетическим материалом через цитоплазматический мостик при конъюгации у бактерий, но с вовлечением репродукции). Такой тип спаривания наблюдается у некоторых зеленых водорослей, например, Спирогира (Spirogyra). Эти водоросли растут как нити клеток. Когда две нити противоположных типов спаривания сближаются, клетки образуют конъюгационные трубки между нитями. Как только трубки сформированы, протопласт из одной клетки перемещается по трубке в другую, чтобы слиться с ней, образуя зиготу.
У инфузорий деление клеток может сопровождаться самооплодотворением (аутогамия), или переносом ядер между двумя клетками без образования зиготы.
У грибов зигомицетов (Zygomycota) две гифы противоположных спаривающих типов образуют специальные структуры, называемые гаметангиями, где они касаются друг друга. Затем гаметангия сливается с зигоспорангием. У других грибов клетки из двух гиф с противоположными типами спаривания происходит слияния цитоплазмы (плазмогамия). Два ядра не сливаются, что приводит к образованию дикарион (клетки с двумя гаплоидными ядрами), образовывающих мицелий. Кариогамия (слияние ядер) встречается в спорангии и приводит к образованию диплоидных клеток (зигот), которые сразу же подвергаются мейозу, образуя споры.
Во многих случаях изогамовое оплодотворение используется организмами, которые также могут размножаться бесполым путем, через бинарное деление, почкование или бесполые споры. Переход к режиму полового размножения часто вызван изменением от благоприятных к неблагоприятным условиям среды. Оплодотворение часто приводит к образованию зиготических покоящихся спор с толстыми стенками, которые могут выдержать суровые условия и прорастают, как только они становятся снова благоприятными.
Эволюция анизогамии
Анизогамия — это процесс оплодотворение мелкими мужскими гаметами (сперматозоиды) крупных женских гамет (яйцеклетки). Разница в размере гамет является принципиальным различием между самцами (мужчинами) и самками (женщинами). Гетерогамия впервые развилась у многоклеточных гаплоидных видов, когда была установлена дифференциация различных типов спаривания.
Для эволюции анизогамии были предложены две теории
Основное внимание уделяется теории конкуренции спермы; другой акцент делается на взаимодействие различных типов спаривания для борьбы с рисками оплодотворения. Обе теории предполагают, что анизогамия возникла из-за дизруптивного (разрывающего) отбора, действующего на предковую изогамную популяцию, и что существует компромисс между большим числом гамет и меньшим приспособлением каждой отдельной гаметы, поскольку общий ресурс, который один организм может вложить в воспроизведение фиксируется
Теория конкуренции спермы была первой теорией, объясняющей эволюцию анизогамии, предложенную Джеффом Паркером. Он предположил, что, поскольку размер зиготы определяется размерами обоих гамет, то меньший размер гаметы, уменьшит размер зиготы не менее чем на половину. Поэтому существует большая выгода для уменьшения размера гамета, чтобы значительно увеличить их количество.
Однако конкуренция существует не только среди спермы, но также и среди яйцеклеток. Паркер не дал объяснений, почему именно сперма стала меньше, а не яйцеклетки, или что привело к тому, что прото-сперма и прото-яичники стали неравными. Фактически, недавняя математическая модель показывает, что конкуренция сперматозоидов не является ни необходимой, ни достаточной для эволюции анизогамии. Конкуренция сперматозоидов приводила только к тому, что сперматозоиды становились все меньше и меньше.
Предполагается, что реальной причиной эволюции анизогамии являются риски оплодотворения
Важной проблемой для размножения многоклеточных организмов является перенос гамет с места на место для оплодотворения. Однако транспортировка имеет вероятность отказа и создает риски оплодотворения
Когда риск возрастает, гаметы одного типа спаривания должны уменьшаться, чтобы увеличить число для преодоления этого риска, а гаметы другого типа спаривания должны стать больше, чтобы компенсировать потери питательных веществ у зигот.
Таким образом, риски оплодотворения и взаимодействие различных типов спаривания могли вызвать эволюцию анизогамии. Сперма развивается, чтобы увеличить вероятность оплодотворения, а яйцеклетка развиваются, чтобы компенсировать потерю питательных веществ. Кроме того, сперматозоиды могут становится еще меньше, если яйцеклетки станут более крупными, чтобы обеспечить питательными веществами зиготы. Это взаимодействие может служить причиной того, что многие организмы имеют микроскопических сперматозоидов и чрезвычайно крупные яйцеклетки.
Встречается ли изогамия у людей?
В результате изогамии могут появляться однополые организмы, имеющие в кариотипе хромосомы исключительно подтипа Х. У живых клеток именно Х считаются эволюционно более старыми. Высшие растения и абсолютно все животные не могут использовать изогамию для продолжения рода: у зверей, высших растений и людей она сменяется оогамией, когда женские и мужские нуклеопротеидные структуры дифференцированы, и в результате появляется особь с ХУ-хромосомами (мужской пол) и с ХХ-хромосомами (женский пол).
Наблюдение за растениями помогает разобраться, почему пол малыша в момент зачатия определяют именно сперматозоиды, а не яйцеклетка. Женская репродуктивная клетка в процессе созревания, деления и выхода в полость для оплодотворения способна получать только две хромосомы Х от матери, тогда как сперматозоид во время деления может взять от организма отца как Х, так и У-хромосому.
Следовательно, если соединятся две Х, получится девочка, а если Х и У – мальчик.
У людей изогамии не бывает – это попросту невозможно. В человеческих клетках при адекватном генотипе могут быть только две одинаковые хромосомы – у женского пола, и две разные – у мужского. Таким образом, процесс деления клеток в теле сопровождается митозом (это деление, когда получается клетка с одинаковым количеством хромосом) и мейозом (происходит редукция количества хромосом, когда половина оказывается Х или У).
Различные формы изогамия:
А) изогамия подвижных клеток, Б) изогамия неподвижных клеток, В) конъюгация.
Различные формы анизогамии:
А) анизогамия подвижных клеток, Б) оогамия (яйцеклетка и сперматозоид), В) анизогамия неподвижных клеток (яйцеклетка и сперматозоиды).
Анизогамия (также называемый гетерогамия) является формой половое размножение что включает в себя объединение или слияние двух гаметы, которые различаются по размеру и / или форме. (Родственные прилагательные: анизогамный и анизогамный.) [1] Меньшая гамета считается мужской ( сперматозоид), тогда как более крупная гамета считается женской (обычно яйцеклетка, если неподвижный). [2]
Есть несколько видов анизогамии. Обе гаметы могут быть бичеванный и поэтому подвижный. Альтернативно, обе гаметы могут быть не флагеллированными. Последняя ситуация встречается в некоторых водоросли и растения. в красная водоросль Полисифониянеподвижные яйца удобренный неподвижной спермой. В цветущие растения, гаметы - неподвижные клетки внутри гаметофиты.
Форма анизогамии, встречающаяся в животные, включая люди, является оогамия, где большая неподвижная яйцеклетка (яйцеклетка) оплодотворяется маленькой подвижной спермой (сперматозоид). Яйцо оптимизировано для долголетия, в то время как маленькие сперматозоиды оптимизированы для подвижности и скорости. Размер и ресурсы яйцеклетки позволяют производить феромоны, которые привлекают плавающие сперматозоиды. [3]
Содержание
Половой диморфизм
Анизогамия - фундаментальная концепция половой диморфизм это помогает объяснить фенотипические различия между полами. [4] У большинства видов существует мужской и женский пол, оба из которых оптимизированы для репродуктивного потенциала. Из-за разного размера и формы гамет как у мужчин, так и у женщин развились физиологические и поведенческие различия, которые оптимизируют индивидуальные плодовитость. [4] Поскольку большинство самок, откладывающих яйца, как правило, должны нести потомство и иметь более ограниченный репродуктивный цикл, это обычно делает самок ограничивающий фактор в репродуктивном успехе самцов вида. Этот процесс справедлив и для женщин, выбирающих самцов, и если предположить, что самцы и самки выбирают партнеров по разным признакам, это приведет к фенотипическим различиям между полами на протяжении многих поколений. Эта гипотеза, известная как Принцип Бейтмана, используется для понимания эволюционного давления, оказываемого на мужчин и женщин из-за анизогамии. [5] Хотя это предположение подвергается критике, это общепринятая модель полового отбора у анизогамных видов. Отбор по разным признакам в зависимости от пола в пределах одного и того же вида известен как отбор по признаку пола и объясняет различные фенотипы, обнаруживаемые между полами одного и того же вида. Этот специфичный для пола выбор между полами с течением времени также приводит к развитию вторичные половые признаки, которые помогают мужчинам и женщинам в репродуктивном успехе.
У большинства видов оба пола выбирают себе партнеров на основе имеющихся фенотипов потенциальных партнеров. [5] Эти фенотипы видоспецифичны, что приводит к различным стратегиям успешного полового размножения. Например, у морских слонов сексуально отбираются крупные самцы, поскольку их большой размер помогает самцам отбиваться от других самцов, а у пауков сексуально отбираются маленькие самцы, поскольку они могут быстрее спариваться с самкой, избегая при этом сексуальный каннибализм. [6] Однако, несмотря на широкий диапазон фенотипов, отобранных по половому признаку, большинство анизогамных видов следуют набору предсказуемых желаемых черт и избирательного поведения, основанного на общих моделях репродуктивного успеха.
Женские фенотипы
За внутренние удобрения, женщины вкладывают большие средства в воспроизводство, поскольку они обычно тратят больше энергии в течение одного репродуктивного события. Это видно уже оогенез, поскольку самка жертвует числом гамет ради размера гамет, чтобы лучше увеличить шансы на выживание потенциальной зиготы; процесс более энергетически требовательный, чем сперматогенез у мужчин. [7] Оогенез происходит в яичник, особый женский орган, который также вырабатывает гормоны, чтобы подготовить другие специфические для женщин органы к изменениям, необходимым в репродуктивных органах, чтобы облегчить доставку яиц во внешних удобрениях и развитие зигот во внутренних удобрениях. Образовавшаяся яйцеклетка не только большая, но иногда даже неподвижная, что требует контакта с более подвижной спермой, чтобы вызвать оплодотворение. [7]
Поскольку этот процесс требует больших затрат энергии и времени для женщин, выбор партнера часто интегрируется в поведение женщины. [4] Самки часто очень избирательны по отношению к самцам, с которыми они выбирают для размножения, поскольку фенотип самца может указывать на физическое здоровье самца и его наследственные особенности. Самки используют выбор партнера, чтобы оказывать давление на самцов, заставляя их проявлять свои желаемые черты перед самками. ухаживание, и в случае успеха самец получает возможность размножаться. Это побуждает самцов и самок определенных видов вкладывать средства в ухаживания, а также в черты, которые могут показать физическое здоровье потенциальному партнеру. Этот процесс, известный как половой отбор, [4] приводит к развитию черт, облегчающих репродуктивный успех, а не индивидуального выживания, таких как раздутый размер королевы термитов. Для самок также важно выбирать потенциальных партнеров, которые могут иметь инфекция, передающаяся половым путем, поскольку болезнь может не только повредить репродуктивную способность самки, но и повредить полученное потомство. [8]
Хотя это не редкость у мужчин, женщины больше связаны с родительская забота. [9] Поскольку у самок более ограниченный репродуктивный график, чем у самцов, самка часто вкладывает больше средств в защиту потомства до достижения половой зрелости, чем самец. Как и при выборе партнера, уровень родительской заботы сильно различается у разных видов и часто зависит от количества потомства, производимого за один половой акт. [9]
У большинства видов, таких как Drosophila melanogaster, самки могут использовать хранение спермы, [10] процесс, с помощью которого самка может накапливать излишки спермы от партнера и оплодотворять свои яйцеклетки спустя долгое время после репродуктивного события, если возможности спаривания уменьшаются или качество партнеров ухудшается. Имея возможность спасти сперму от более желанных партнеров, самка получает больший контроль над своим репродуктивным успехом, что позволяет самке быть более избирательной по отношению к самцам, а также потенциально более частым временем оплодотворения, если самцов мало. [10]
Мужские фенотипы
Для мужчин всех видов производимые ими сперматозоиды оптимизированы для обеспечения оплодотворение женского яйца. Эти сперматозоиды создаются в результате сперматогенеза, формы гаметогенез который фокусируется на развитии максимально возможных гамет за один половой акт. [7] Сперматогенез происходит в яички, специфический мужской орган, который также производит гормоны, вызывающие развитие вторичных половых признаков. Поскольку мужские гаметы энергетически дешевы и обильны при каждой эякуляции, мужчина может значительно увеличить свой сексуальный успех, спариваясь гораздо чаще, чем женщина. [7] Сперматозоиды, в отличие от яйцеклеток, также подвижны, что позволяет сперме плавать к яйцеклетке через половые органы самки. Конкуренция спермы также является важным фактором развития сперматозоидов. Только один сперматозоид может оплодотворить яйцеклетку, и поскольку самки потенциально могут воспроизводиться более чем одним самцом до того, как произойдет оплодотворение, производство сперматозоидов, которые будут более быстрыми, более многочисленными и более жизнеспособными, чем те, которые производятся другими самцами, может дать репродуктивное преимущество самцов. [7]
Поскольку самки часто являются ограничивающим фактором в репродуктивном успехе вида, самки часто ожидают, что самцы будут искать и соревноваться за самку, известную как внутривидовая конкуренция. [5] Это можно увидеть на таких организмах, как бобовые жуки, поскольку самцы, которые чаще ищут самок, часто более успешны в поиске партнеров и воспроизведении. У видов, подвергающихся этой форме отбора, подходящий самец будет быстрым, с более совершенными органами чувств и пространственным восприятием. [5]
Некоторые вторичные половые характеристики предназначены не только для привлечения партнеров, но и для конкуренции с другими самцами за возможность совокупления. Некоторые структуры, такие как рога оленей, могут способствовать репродуктивному успеху самцов, предоставляя оружие, не позволяющее конкурирующим самцам добиться репродуктивного успеха. [8] Однако другие структуры, такие как большие разноцветные рулевые перья, обнаруженные у самцов павлинов, являются результатом Рыбак-беглец а также еще несколько видоспецифичных факторов. Из-за того, что самки выбирают определенные черты у самцов, со временем эти черты преувеличиваются до такой степени, что могут препятствовать выживанию самцов. [8] Однако, поскольку эти черты очень полезны для полового отбора, их полезность в предоставлении большего количества возможностей для спаривания перевешивает возможность того, что эта черта может привести к сокращению продолжительности ее жизни из-за хищничества или голода. Эти желательные черты выходят за рамки физических частей тела и часто включают ухаживания и ухаживания. свадебные подарки также.
Хотя некоторые модели поведения мужчин должны работать в рамках параметров загадочного женского выбора, некоторые мужские черты работают против него. Достаточно сильные самцы, в некоторых случаях, могут навязывать себя самке, вынуждая оплодотворение и отвергая выбор самки. [11] Поскольку это часто может быть опасно для самок, эволюционная гонка вооружений между полами часто является результатом.
Эволюция
Анизогамия - это феномен оплодотворение крупных гамет (яйцеклетки, яйцеклетки) маленькими гаметы (сперма клетки: сперматозоиды или сперматозоиды). Разница в размере гамет - принципиальная разница между самки и самцы. Анизогамия впервые возникла в многоклеточных гаплоидный виды после дифференциации разных типы вязки уже было установлено. [12] Однако в Аскомицеты, анизогамия произошла от изогамии до типов спаривания. [13] [ неудачная проверка ]
Три основных теории эволюции анизогамии - это конкуренция гамет, ограничение гамет и внутриклеточные конфликты, но последняя из этих трех недостаточно хорошо подтверждена имеющимися данными. [14] Как конкуренция гамет, так и ограничение гамет предполагают, что анизогамия возникла в результате разрушительного отбора, действующего на предков изогамной популяции с внешним оплодотворением, из-за компромисса между большим числом гамет и размером гамет (что, в свою очередь, влияет на выживаемость зиготы), потому что общий ресурс один предполагается, что человек может инвестировать в воспроизводство. [15]
Первая формальная математическая теория, предложенная для объяснения эволюции анизогамии, была основана на ограничении гамет: [16] эта модель предполагала, что естественный отбор приведет к размеру гамет, который приведет к наибольшему числу успешных оплодотворений в популяции. [16] [17] [18] Если предположить, что определенное количество ресурсов, обеспечиваемых гаметами, необходимо для выживания полученной зиготы, и что существует компромисс между размером и количеством гамет, то этот оптимум оказался таким, при котором оба образуются мелкие (мужские) и большие (женские) гаметы. Однако эти ранние модели предполагают, что естественный отбор действует в основном на уровне популяции, что сегодня, как известно, является весьма проблематичным предположением. [19]
Таким образом, существует относительно прочная теоретическая база для понимания этого фундаментального перехода от изогамии к анизогамии в эволюции репродукции, который, как предполагается, будет связан с переходом к многоклеточности. Фактически, Hanschen et al. (2018) демонстрируют, что анизогамия произошла от изогамных многоклеточных предков и что впоследствии анизогамия будет приводить к вторичному половому диморфизму. [12] Существуют сравнительные эмпирические доказательства теорий конкуренции гамет, [14] [28] [29] хотя трудно использовать это свидетельство, чтобы полностью разделить теории конкуренции и ограничений, поскольку их проверяемые прогнозы схожи. [15] Также было заявлено, что некоторые организмы, использованные в таких сравнительных исследованиях, не соответствуют теоретическим предположениям. [30]
Ценной модельной системой для изучения эволюции анизогамии являются водоросли вольвоцина, группа которых хлорофиты довольно уникален тем, что его современные виды демонстрируют разнообразие систем спаривания (изогамия и анизогамии) в дополнение к крайностям в обоих одноклеточность и многоклеточность с разнообразием форм у видов промежуточных размеров. [31] Морские водоросли были тщательно изучены, чтобы понять траектории таких разнообразных репродуктивных систем, [32] эволюция пола и типов спаривания, [33] а также адаптивность и устойчивость анизогамии. [34] [32] [12]
Читайте также: