
6 охарактеризуйте механизмы взаимодействия вируса иммунодефицита с клеткой кратко
Обновлено: 05.07.2024
Проникнув в организм, вирус в первую очередь атакует клетки, содержащие специфический для него рецептор CD4. Этот рецептор имеют в большом количестве Т-хелперы, в меньшем – макрофаги и моноциты, особенно к вирусу чувствительны Т-хелперы.
Вирус распознает CD4-рецепторы с помощью своего белка gр120. Процесс взаимодействия ВИЧ с клеткой протекает по следующей схеме: рецепторопосредованная адсорбция ? окаймленная ямка ? окаймленный пузырек ? лизосома. В неймембрана вириона сливается с мембраной лизосомы, и нуклеокапсид, освобожденный от суперкапсида, выходит в цитоплазму; на пути к ядру он разрушается, и высвобождаются геномная РНК и ассоциированные с ней компоненты сердцевины. Далее обратная транскриптаза синтезирует на вирионной РНК минус-цепь ДНК, затем РНК-аза Н разрушает вирионную РНК, а вирусная ДНК-полимераза синтезирует плюс-цепь ДНК. На концах ДНК-провируса образуются 5'-LTR и 3'-LTR. ДНК-провирус может находиться в ядре некоторое время в неактивной форме, но рано или поздно он с помощью своей интегразы встраивается в хромосому клетки-мишени. В ней провирус находится в неактивном состоянии до тех пор, пока данный Т-лимфоцит не будет активирован микробными антигенами или другими иммунокомпетентными клетками. Активация транскрипции клеточной ДНК регулируется особым ядерным фактором (NF-kB). Он является ДНК-связывающим белком и вырабатывается в большом количестве при активации и пролиферации Т-лимфоцитов и моноцитов. Этот белок связывается с определенными последовательностями клеточной ДНК и сходными последовательностями LTR ДНК-провируса и индуцирует транскрипцию как клеточной ДНК, так и ДНК-провируса. Индуцируя транскрипцию ДНК-провируса, он и осуществляет переход вируса из неактивного состояния в активное и соответственно персистентной инфекции – в продуктивную. Пребывание провируса в неактивном состоянии может продолжаться очень долго. Активация вируса является критическим моментом в его взаимодействии с клеткой.
С момента проникновения вируса в клетку начинается период ВИЧ-инфекции – вирусоносительства, которое может продолжаться 10 и более лет; а с момента активации вируса начинается болезнь – СПИД. С помощью своих регуляторных генов и их продуктов вирус начинает активно размножаться. ТАТ-белок может повысить скорость репродукции вируса в 1000 раз. Транскрипция вируса имеет сложный характер. Она включает образование как полноразмерных, так и субгеномных мРНК, сплайсинг мРНК, а далее происходит синтез структурных и регуляторных белков.
Другой белок-предшественник Prl60Gag-Pol (белок с м. м. 160 кД) синтезируется в результате сдвига рамки считывания рибосомой при трансляции 3'-конца гена gag в области, расположенной непосредственно перед участком, кодирующим белок р6. Этот полипротеин Gag-Pol содержит неполную последовательность Gag-белка (1 – 423 аминокислоты) и последовательности Pol, которые включают домены PR, RT и IN. Молекулы полипротеина Gag-Pol также синтезируются на свободных полири-
босомах и транспортируются к плазматической мембране. Полипротеин Prl60GagPol содержит все присущие полипротеину Gаg сайты межмолекулярных взаимодействий и сайты связывания с мембраной. Поэтому молекулы полипротеина Gag-Pol сливаются с мембраной и наряду с Gag-молекулами включаются в формирующиеся вирионы, в результате чего появляется активная протеаза и начинается процесс созревания вириона. Протеаза ВИЧ-1 высокоактивна только в виде димера, поэтому для ее самовырезания из Prl60Gаg-Pol требуется димеризация этих молекул. Созревание вириона заключается в том, что освободившаяся активная протеаза разрезает Prl60Gag-Pol и Gаg55 в узнаваемых ею сайтах; образуются белки р17, р24, р7, р6, ревертаза, интеграза и происходит их ассоциация в вирусную структуру.
Белок Env синтезируется на рибосомах, связанных с мембранами эндоплазматического ретикулума, затем он гликозилируется, разрезается клеточной протеазой на gр120 и gр41 и транспортируется на клеточную поверхность. При этом gр41 пронизывает мембрану и связывается матриксными доменами молекулы Gаg-белка, ассоциированными с внутренней поверхностью мембраны. Эта связь сохраняется и в зрелом вирионе.
Таким образом, сборка вирусных частиц заключается в агрегации белков-предшественников и связанных с ними молекул РНК на плазматической мембране клетки-хозяина, образовании незрелых вирионов и их высвобождении путем почкования с клеточной поверхности. При почковании вирион окружает себя клеточной мембраной, в которую встроены молекулы gр41 и gр120. Во время почкования или, возможно, после высвобождения вирионов происходит их созревание, которое осуществляется при помощи вирусной протеазы и заключается в протеолитическом нарезании белков-предшественников Рr55Gаg и Рrl60Gаg-Pol на белки зрелого вируса и их ассоциации в определенные структурные комплексы. Ведущую роль в процессах морфогенеза вируса играет полипротеин-предшественник Рr55Gag, который организует и осуществляет сборку незрелого вириона; процесс его созревания завершает специфическая вирусная протеаза.
Антигенные варианты. Вирус иммунодефицита очень изменчив. Даже из организма одного больного могут быть выделены штаммы вируса, существенно различающиеся по антигенным свойствам. Такой изменчивости способствуют интенсивное разрушение клеток СD4 + и мощный антительный ответ на ВИЧ-инфекцию. У больных из Западной Африки выделена новая форма ВИЧ, биологически близкая к ВИЧ-1, но иммунологически отличающаяся от него, – ВИЧ-2. Гомология первичной структуры геномов этих вирусов составляет ~42 %. ДНК-провирус ВИЧ-2 содержит 9671 п. н., а его LTR – 854 п. н. ВИЧ-2 впоследствии выделен и в других регионах мира. Перекрестного иммунитета между ВИЧ-1 и ВИЧ-2 нет. Известны две крупные формы ВИЧ-1: О (Outlier) и М (Маjor), последнюю подразделяют на 10 субтипов (А – J). В России циркулируют 8 субтипов (А – Н).
Методы культивирования. ВИЧ-1 и ВИЧ-2 удается культивировать в клетках только одного клона ТCD4-лимфоцитов – Н9, полученного из лейкозных ТСВ4лимфоцитов. Для этих же целей могут быть использованы и монослойные культуры клеток астроцитов, в которых ВИЧ-1 хорошо размножается. Из животных к ВИЧ-1 восприимчивы шимпанзе.
Резистентность вируса во внешней среде невелика. Он погибает под влиянием солнечных лучей и УФ-облучения, разрушается при 80 °C в течение 30 мин, при обработке обычно применяемыми дезинфицирующими веществами – в течение 20 – 30 мин. Для обеззараживания вируссодержащего материала необходимо пользоваться микобактерицидными дезинфицирующими веществами, поскольку они эффективны против микроорганизмов, обладающих самой высокой резистентностью.
Начало заболевания обычно проявляется на фоне интенсивной репродукции вирусов в клетках РЭС, эпителии дыхательной или пищеварительной систем, крови и др. Возникающие в результате иммунного ответа антитела в определенной степени препятствуют распространению вируса в организме, но существенно не влияют на процессы его репродукции. Создается впечатление, что антитела не только не определяют исход первичной вирусной инфекции, а более того, могут маскировать циркулирующий вирус в крови и других жидкостях организма. Более выражены в начальном периоде инфицирования факторы неспецифической защиты - повышенная температура, повышение уровня различных ингибиторов, интерферона и др.
![]()
В патогенезе СПИД, по мнению большинства исследователей, решающее значение имеет избирательное цитопатическое воздействие вируса СПИД на Т-лимфоциты-хелперы-индукторы, что в первую очередь проявляется в снижении противовирусного, противомикробного и противоопухолевого иммунитета. Следствием указанного является накопление вируса в крови и интенсивное его распространение в организме, усугубление течения и необратимое развитие оппортунистических инфекций, ускорение злокачественного роста вследствие преморбидного действия персистирующего в клетках опухоли частиц вируса и поражения натуральных киллеров. Первичное селективное цитодеструктивное действие вируса на Т-хелперы-индукторы проявляется в развитии лимфопении, снижении числа циркулирующих в крови Т4-клеток, нарушении соотношения Т4/Т8 и угнетение пролиферации Т-лимфоцитов в ответ на действие митогенов (фитогемагглютинин, конкавалин А, специфические антигены), снижении или полном отсутствии реакции кожной гиперчувствительности к кожным аллергенам, снижении пролиферации аутологичных лимфоцитов в смешанной реакции, уменьшение продукции гамма-глобулина лейкоцитами больных оппортунистическими инфекциями, ослабление экспрессиТ-клеточных рецепторов к интерлейкину-2, дефектности продукции лимфокинов, поликлональной активации В-лимфоцитов с продукцией большого числа фракций гамма-глобулина.
Иммунологические нарушения при СПИД проявляются также в повышении уровня циркулирующих иммунных комплексов, антилимфоцитарных антител невыясненной специфичности, в увеличении количества и лабильности альфа-интерферона. Однако описанные выше иммунологические аномалии не являются строго специфичными именно для СПИД , они встречаются и при иммунодефицитах другой этиологии; дифференциация специфичности поражения особенно затруднительна в начальный период заболевания.
Возбудитель СПИД имеет выраженный тропизм к Т-лимфоцитам и другим клеткам с антигенной формулой — ОКТ-4 (Т-хелперы, В-клетки, моноциты, макрофаги). Указанное сродство обусловлено чрезвычайно высоким аффинитетом гликопротеида наружной оболочки вириона к рецептору Т4 (CD4), располагающемуся на поверхности Т-хелперов. Было доказано, что мембранный белок лимфоцитов CD4 служит рецептором для вируса СПИД. Что касается вирусных белков, то лишь один из них с молекулярной массой 11000 Д (белок gp120) способен связываться с CD4 белком.
![]()
Для ряда вирусов человека установлена связь клеточной чувствительности к ним с определенными хромосомами человеческих клеток. Например, в хромосоме19 локализованы гены чувствительности к вирусам полиомиелита, в хромосоме 3 — к вирусу герпеса, в хромосоме 21 — к вирусам Коксаки В. В какой хромосоме существуют гены чувствительности к вирусу СПИД, пока не установлено.
Генетические исследования показали связь развития СПИД с определенным типом антигенов главного комплекса гистосовместимости (МНС). Установлено, что среди больных СПИД весьма значительно количество лиц, имеющих HLA-DR5 антигены. Подтверждено, что лица с таким фенотипом лимфоцитов составляют группу с повышенным риском заболевания саркомой Капоши, встречающейся в 6 раз чаще, чем в других группах.
Известно, что иммунореактивность зависит как от структуры антигена, так и от генетических особенностей организма и связана с одним из типов клеток или их субпопуляций, участвующих в иммунных реакциях. У слабо реагирующего организма нет Ir-гена (или он по какой-либо причине не реализует свои функции), позволяющего Т-хелперам распознать антиген и оказать помощь В-лимфоцитам. Недостаточность иммунного ответа на уровне В-клеток нередко обусловлена отсутствием другого гена, контролирующего синтез веществ клеточной поверхности (акцепторов), которые воспринимают дополнительный сигнал от Т-хелперов (Leu 3a+). Цепь событий в иммунной системе развертывается следующим образом. Антиген, захваченный макрофагом, разрушается до отдельных фрагментов гидролитическими ферментами. Затем в работу вступают Ir-гены, которые контролируют образование семейства так называемыхIa-белков. У разных индивдуумов или линий животных эти белки отличаются друг от друга по структуре, отражая различие в генах, которые обеспечивают их синтез. Если белки макрофагов способны вступать во взаимодействие с данным антигеном, то образовавшийся комплекс антиген-белок мигрирует к мембране макрофага и оказывается на ее поверхности. В результате создаются условия для работы Т-хелперов. Т-хелперы могут наиболее четко распознавать антиген только в ассоциации его с Ia-белком. Идентифицировав антиген, измененный Ia-белком,Т-хелперы оказывают необходимую помощь В-клеткам, которые без этого не приступают к синтезу иммуноглобулинов. Помощниками Т-В-кооперации служат также различные белки регуляторы, лимфокины, секретируемые Т-клетками.
Обнаружены антитела, нейтрализующие вирус СПИД, у больных со СПИД и СПИД-комплексом..
![]()
В последнее время установлено, что ВИЧ в иммунной системе инфицирует прежде всего макрофаги и моноциты, с помощью которых и распространяется в организме. Местом сохранения вполне могут быть также тромбоциты, В-лимфоциты, эпителиальные клетки, глиальные клетки нервной системы.
Антиген встречается с клетками Лангерганса, которые представляют его (в комплексе с антигеном гистосовместимости) Т-клеткам, запрограммированным отвечать на данный антиген ( подробнее о клетках Лангерганса ниже). Активированные таким образом клетки экспрессируют рецепторы к интерлейкину-1 (ИЛ-1). ИЛ-1 кроме лимфоидных клеток имеет и ряд иных клеток-мишеней, что обуславливает множественную направленность биологической активности этого вещества и его участие в целом ряде процессов, не относящихся собственно к иммунным. Выделяемый кератиноцитами (и, возможно, клетками Лангерганса) фактор заставляет активированные Т-клетки секретировать интерлейкин-2 (ИЛ-2) — важнейший тип лимфокина. Рецептором для ИЛ-2 на клетке является структура носящая название Tac-антигена. ИЛ-2 вступает в связь с рецепторами на поверхности других Т-клеток, отвечающих на данный антиген и находящихся в дерме, лимфатических узлах, селезенке, и инициирует их пролиферацию. Результатом указанного является новая популяция Т-клеток, вырабатывающих антитела к данному антигену.

Известно, что клетками-мишенями для вируса иммунодефицита человека (ВИЧ) являются Т-лимфоциты, дендритные клетки и клетки Лангерганса (их незрелые предшественники), моноциты/макрофаги, эозинофилы, мегакариоциты, тимоциты, некоторые клоны В-лимфоцитов, клетки нервной системы (нейроны, микроглиальные клетки/макрофаги). Эти клетки имеют на мембране молекулы CD4, к которым вирусный эпимембранный гликопротеин gp120(ВИЧ-1) или gp105(ВИЧ-2) проявляет большое сродство. Вместе с тем следует заметить, что существует ряд клеток, которые, не имея CD4, селективно сорбируют, транспортируют на мембране или проводят через себя ВИЧ. Такими клетками являются М-клетки слизистой прямой кишки, граничащие с лимфоидной тканью стенки кишки, и сперматозоиды. По клеточному тропизму изоляты ВИЧ делят на моноцитотропные и лимфоцитотропные. Первые преобладают на начальных стадиях, вторые - в период разгара болезни.
Цель исследования - изучить особенности взаимодействия ВИЧ с Т-клетками иммунной системы. Изучение иммунопатогенеза проведено на основе данных литературы.
Весь процесс взаимодействия ВИЧ с клеткой-мишенью можно разделить на ряд последовательных стадий:
1) связывание вириона с поверхностью клетки и рецепция вируса;
2) слияние мембран вируса и клетки, проникновение вируса внутрь клетки;
3) высвобождение нуклеоида и геномной РНК вируса;
4) синтез провирусной ДНК по матрице геномной РНК вируса;
5) интеграция генома провируса в геном клетки;
6) активация процесса транскрипции с ДНК провируса, трансляция белков вируса;
7) активная репликация вируса, т.е. продукция всех компонентов вируса и формирование из них зрелых дочерних вирионов;
8) высвобождение вирионов и отдельных белков ВИЧ из клетки-хозяина во внешнюю среду и беспрепятственное заражение других клеток, проявление цитопатогенных эффектов ВИЧ.
Ведущим звеном в патогенезе ВИЧ-инфекции является поражение Т-хелперов, которое обусловлено: преждевременным старением и гибелью инфицированных клеток; уничтожением заражённых клеток лимфоцитами-эффекторами антителозависимой клеточной цитотоксичности; блокадой рецепторов CD4 вирусным гликопротеином gp120; аутоиммунными процессами. На Т-хелперы ВИЧ оказывает прямой цитопатогенный эффект. Истощение пула Т-хелперов приводит к тому, что они не могут полноценно обеспечивать свои функции и взаимодействие других иммунокомпетентных клеток. Однако и на ранних этапах течения ВИЧ-инфекции, когда еще нет выраженного снижения содержания CD4+-клеток, а доля инфицированных CD4 + -лимфоцитов не превышает 0,01 % их количества, основное значение в развитии дисбаланса иммунного ответа и формировании иммунодефицита принадлежит нарушениям регуляторных функций Т-хелперов/индукторов. Причиной указанных нарушений является блокада рецептора CD4. G. Furlini с соавт. (1989) установили, что спустя три часа после воздействия ВИЧ-1 (или очищенного рекомбинантного белка gp120) на CD4 + -клетки in vitro наблюдается пик увеличения синтеза и ядерной транслокации белков теплового шока семейства БТШ-70. Эти данные свидетельствуют о способности gp120 запускать каскад процессов, используя сигнальную активность мембран. Одним из таких внутриклеточных процессов является активация системы белков теплового шока, что в свою очередь указывает на нахождение клетки в неблагоприятных условиях и формирование клеточной стресс-реакции. С развитием выраженной виремии количество инфицированных клеток в крови и интенсивность их гибели возрастает. От момента инфицирования до терминальной стадии СПИД содержание CD4 + -клеток уменьшается более чем в 20 раз.
Таким образом, клинические проявления заболевания обусловлены непосредственным патогенным эффектом вируса и его белков на клетки-мишени, истощением пула CD4 + -клеток крови, а также нарушением кооперативных связей и функций иммунокомпетентных клеток, что приводит к формированию иммунодефицита.

Обзор
Распространенность СПИДа в мире на 2009 год.
Автор
Редакторы

Спонсор конкурса — дальновидная компания Thermo Fisher Scientific. Спонсор приза зрительских симпатий — фирма Helicon.
Иммунная система

Рисунок 1. Упрощенная схема иммунного ответа. Молниями указано, как один тип клеток активирует другие. Мф — макрофаг, Б — бактерия, ТХ — Т-хелпер, ВКл — B-клетка, ПлКл — плазматическая клетка, Ат — антитела, Б+Ат — бактерия, покрытая антителами, привлекающими макрофагов, В — вирус, ЗМф — зараженный макрофаг, ТК — Т-киллер, ЗК — клетка, зараженная вирусом, УК — умирающая (апоптотирующая) клетка, В+Ат — вирус, покрытый антителами.
рисунок автора статьи
Жизненный цикл ВИЧ
Видео. Жизненный цикл ВИЧ.
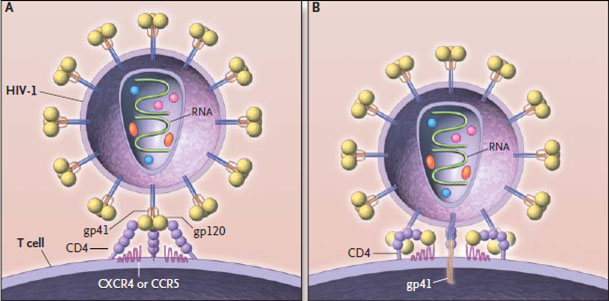
Таким образом, после того как копия вируса в геноме клетки начинает действовать, на её поверхности появляются знакомые нам белки gp41 и gp120, в цитоплазме — остальные вирусные белки и вирусная РНК. И через некоторое время от зараженной клетки начинают отпочковываться всё новые и новые копии ВИЧ.
Способы уничтожения иммунной системы
При отпочковывании вирус использует клеточную мембрану клетки, и со временем это должно вызвать ее разрушение. К тому же, вирусный белок Vpu вызывает увеличение проницаемости мембраны клетки [6]. При активации вируса в клетке появляется неинтегрированная в геном двуцепочечная ДНК [19], наличие которой может рассматриваться клеткой как повреждение ее генетического материала и индуцировать ее смерть через апоптоз (С точки зрения организма такая клетка потенциально может стать раковой). Помимо этого, вирусные белки напрямую нарушают баланс про- и противоапоптотических белков в клетке. К примеру, вирусная протеаза p10 может разрезать противоапоптотический фактор Bcl-2. Белки Nef [26], Env [22] и Tat [24] вовлечены в возрастание количества клеточных белков CD95 и FasL, служащих индукторами апоптоза по т.н. Fas-опосредованному пути. Белок Tat положительно регулирует каспазу 8 [2] — ключевой фактор индукции апоптоза.
Если же клетки не погибли сами, они активно уничтожаются Т-киллерами. (Это стандартный ответ иммунитета на заражение любыми вирусами.)
Помимо этого, при производстве gp120 и Tat часть их выбрасывается зараженной клеткой в кровоток, а это оказывает токсическое действие на организм [4], [28]. gp120 оседает на все СD4 + -клетки (в том числе и здоровые), что имеет три последствия:
- Белок СD4 важен для взаимодействия Т-хелпера с фагоцитом, при слипании же СD4 с gp120 T-хелпер перестает выполнять эту функцию.
- На gp120 как на чужеродный белок образуются антитела; после оседания gp120 на здоровые клетки, последние маркируются антителами, и иммунная система их уничтожает [9].
- После обильного связывания Т-хелпером gp120 клетка умирает сама. Дело в том, что CD4 — это корецептор. Он усиливает сигнал от TCR (T-cell receptor) — главного белка, позволяющего выполнять Т-клеткам функцию иммунитета. В норме эти рецепторы активируются вместе. Если же активируется один тип рецепторов, но не активируется другой, это является сигналом, что что-то пошло не так, и клетка умирает посредством апоптоза.
Антитела, призванные препятствовать инфекции, в случае с ВИЧ часто, наоборот, ее усиливают [28]. Почему так происходит — не совсем понятно. Одну причину мы рассмотрели выше. Возможно, дело в том, что антитела сшивают несколько вирусных частиц вместе, и в клетку попадает не один, а сразу много вирусов. Помимо этого, антитела активируют клетки иммунитета, а в активированной клетке вирусные белки синтезируются быстрее.
Вдобавок, gp120 и gp41 имеют участки, похожие на участки некоторых белков, участвующих в иммунитете (например, MHC-II [25], IgG [23], компонент системы комплемента Clq-A [14]). В результате на эти участки образуются антитела, способные помимо вируса маркировать совершенно здоровые клетки (если на них будут эти белки), с соответствующими для них последствиями.
Так как на поверхности зараженной клетки появляется gp120, то она, как и вирусная частица, будет сливаться с другими СD4-содержащими клетками, образуя огромную многоядерную клетку (синцитий), неспособную выполнять какие-либо функции и обреченную на смерть.
На самом деле, помимо Т-хелперов, СD4 содержатся во множестве других клеток — предшественниках Т-киллеров/Т-хелперов, дендритных клетках, макрофагах/моноцитах, эозинофилах, микроглии (последние четыре — разновидности фагоцитов), нейронах (!), мегакариоцитах, астроцитах, олигодендроцитах (последние две — клетки, питающие нейроны), клетках поперечно-полосатых мышц и хорионаллантоиса (присутствует в плаценте) [29]. Все эти клетки способны заражаться ВИЧ, но сильно токсичен он лишь для Т-хелперов. Это и определяет основное клиническое проявление СПИДа — сильное снижение количества Т-хелперов в крови. Однако для других клеток заражение тоже не проходит бесследно — у больных СПИДом часто снижена свертываемость крови, имеются неврологические и психиатрические отклонения [16], наблюдается общая слабость.
Пути заражения ВИЧ
Внимание! Ввиду того, что СПИД является венерическим заболеванием, информация, приведенная в этом разделе, может вас оскорбить либо вызвать неприятные эмоции при прочтении. Будьте осторожны.
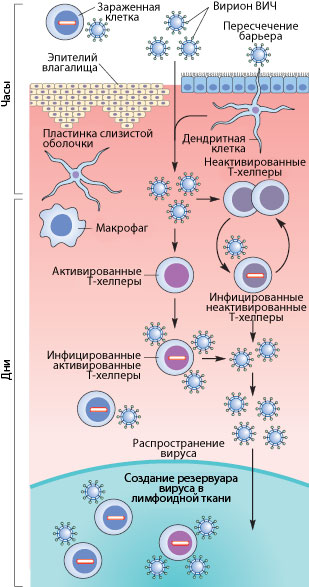
Рисунок 3. Пути проникновения ВИЧ в организм при половом контакте. Слева указано примерные временные рамки каждого этапа.
Как же происходит первичное заражение человека ВИЧ? Думаю, что с переливанием крови или с использованием общего шприца при употреблении наркотиков читателю все понятно — вирус доставляется напрямую из зараженной крови в здоровую. При вертикальном способе передачи инфекции (от матери к ребенку) заражение может произойти во время внутриутробного развития, в момент родов или в период кормления ребенка грудью (т.к. вирус и зараженные вирусом клетки содержатся в молоке матери) [17], [20]. Тем не менее, даже у ребенка, рожденного от ВИЧ-инфицированной матери, есть шанс остаться здоровым, особенно если мать использует ВААРТ (высокоактивную антиретровирусную терапию) [5]. А почему же люди заражаются при половом контакте?
Механизмы передачи вируса при половом акте не до конца ясны. Тем не менее, они понятны в общих чертах. Итак, рассмотрим барьеры, которые необходимо преодолеть вирусу, чтобы добраться до макрофагов и Т-хелперов. К сожалению, рассматривать особо нечего: барьер только один — это слой эпителиальных клеток и вырабатываемая ими слизь, находящиеся во влагалище (а также пенисе, прямой кишке, глотке). Барьер этот тонок — он может быть всего одну клетку в толщину, — но для вируса труднопреодолим, доказательством чего может служить сравнительно небольшой процент заражений — примерно 1–2 случая на 100 половых контактов. Итак, и какими же путями преодолевает вирус это препятствие?
Наиболее очевидный механизм — это микротравмы. При обычном половом контакте, а особенно при анальном сексе, почти неизбежно появляются микротрещины; при наличии же венерических заболеваний часто возникают изъязвления слизистой. Через эти микротравмы вирус, содержащийся в сперме и влагалищном секрете, может проникнуть к иммунокомпетентным клеткам [17], [20].
Помимо дендритных клеток, в кишечнике существует другой тип клеток, предназначенный для ознакомления организма с антигенами внешней среды. Это так называемые М-клетки, пропускающие через себя довольно крупные объекты неповрежденными из просвета кишечника. В т.ч. через них может проходить и вирион ВИЧ.
Свой вклад в заражение вносит и провоспалительный процесс, в норме присутствующий в женских половых путях из-за наличия в них спермы. Благодаря ему Т-клетки подвержены активации, что способствует их восприимчивости к инфекции.
Итак, риск заразиться ВИЧ возрастает в ряду (Классический секс → Анальный секс → переливание крови / использование общих шприцов при принятии наркотиков). Что же касается орального секса, то весьма небольшой шанс заразиться есть только у принимающей стороны — слюна и тем более желудочный сок инактивирует ВИЧ. Теоретически, этим способом может заразиться и мужчина, но для этого необходимы ранки на пенисе.
При поцелуях и обычных бытовых контактах вирус не передается — он очень неустойчив в окружающей среде .
Заключение
. Все эти факты позволяют надеяться, что вскоре ВИЧ, как и оспа, останется в прошлом.
Читайте также: