
Возникновение возбуждения и его проведение кратко
Обновлено: 04.07.2024
Общие представления о физиологии возбудимых тканей
Общие представления о физиологии возбудимых тканей
- Биопотенциал покоя – это разность потенциалов между наружной и внутренней поверхностью мембраны клетки в покое. Наружная поверхность мембраны клетки имеет положительный заряд, а внутренняя – отрицательный.
Биопотенциал покоя регистрируется внутриклеточным методом – с помощью микроэлектродов, один из которых вводится внутрь клетки (рис.1).
Рисунок 1. Схематическое представление метода регистрации биопотенциалов.
В эксперименте биопотенциал покоя можно зарегистрировать между повреждённым и неповреждённым участком ткани. Повреждённый участок является моделью внутренней поверхности мембраны клетки.
При внутриклеточном отведении перезарядка мембраны регистрируется под одним электродом (однофазный ПД), при внеклеточном отведении потенциал действия проходит через два электрода (регистрируется двухфазный ПД).
- Биопотенциал действия – это кратковременные высокоамплитудные изменения МПП, которые возникают при возбуждении. ПД регистрируется в раздражаемых тканях, в которых возникает волна возбуждения (рис.2). Измеряется ПД с помощью внутриклеточного отведения и внеклеточного отведения.

Рисунок 2. Потенциал действия, основные его фазы.
Современная, экспериментально доказанная, мембранно-ионная теория возникновения биопотенциалов (Ходжкин, Хаксли, Катц).
Основные положения:
- Электрические процессы возникают на плазматической мембране клетки, которая состоит из бимолекулярного слоя липидов (остов мембраны) и белков, которые выполняют различные функции в мембране: рецепторную, ферментативную, образуют в ней каналы и насосы (рис.3).

Рисунок 3. Строение биологической мембраны.

Рисунок 4. Строение ионного канала.
Каналы для натрия имеют два типа ворот: быстрые активационные и медленные инактивационные. В покое открыты медленные инактивационные и закрыты быстрые активационные. При возбуждении происходит открытие быстрых активационных и медленное закрытие медленных инактивационных, т.е. на короткий промежуток времени оба типа ворот открыты (рис.5).

Рисунок 5. Работа активационные и инактивационный воротных механизмов натриевого ионного канала.
Калиевые каналы имеют только медленные ворота.
Насосы выполняют функцию транспорта через мембрану ионов против градиента концентрации, для их работы используется энергия АТФ.
- По обе стороны мембраны существует концентрационный градиент.
Внутри клетки в 40 раз > К+; t;/p>
Вне клетки: в 20-30 раз > Na+,
в 50 раз > Cl-.
- Мембрана пропускает молекулы жирорастворимых веществ, а анионы органических кислот не проходят. Мембрана проницаема для воды, для ионов проницаемость мембраны различна: для калия в состоянии покоя проницаемость почти в 25 раз больше, чем для натрия. При возбуждении увеличивается проницаемость и для калия (постепенно), и для натрия (быстро, но на очень короткий промежуток времени).
МПП - один из основных показателей состояния физиологического покоя клетки. При увеличении внеклеточной концентрации калия уменьшается МПП, т.к. уменьшается диффузия калия из клетки в связи со снижением его концентрационного градиента. При действии веществ, блокирующих ресинтез АТФ, т.к. прекращается работа натрий-калиевого насоса, также снижается МПП. Ионы натрия и хлора входят в клетку, но ввиду низкой проницаемости значительного влияния на МП не оказывают.
Рисунок 6. Механизм работы натрий-калиевого насоса.
ПД состоит из пикового потенциала, который образуется фазой деполяризации, реверсии и реполяризации, и следовых потенциалов (рис.2).
Следовые потенциалы:
Отрицательный (следовая деполяризация);
Положительный (следовая гиперполяризация).
Причиной следовых потенциалов являются дальнейшие изменения соотношения между входом натрия в клетку и выходом калия из нее. При следовой деполяризации отмечается остаточный ток натрия в клетку при одновременном снижении калиевого тока. При следовой гиперполяризации – остаточное усиление тока калия из клетки при одновременной активации натрий-калиевого насоса.
Условия, необходимые для возникновения возбуждения (законы раздражения).
Возбудимость тканей различна. Чтобы вызвать возбуждение, раздражитель должен обладать:
1. Достаточной силой – закон порога.
2. Крутизной (градиентом) нарастания этой силы – закон аккомодации.
3. Временем действия – закон силы-времени.
Изменение возбудимости тканей при возбуждении.
При возбуждении возбудимость тканей претерпевает определенные изменения в зависимости от фаз ПД (рис.7):
I – супернормальная возбудимость (первичная) соответствует локальному ответу, при этом два подпороговых стимула, нанесенных с интервалом времени, короче длительности локального ответа могут суммироваться и вызывать ПД;
II – абсолютная рефрактерность – соответствует регенеративной деполяризации и реверсии, при этом ткань становится абсолютно невозбудимой и не отвечает на самые сильные раздражители;
III – относительная рефрактерная фаза, соответствует реполяризации, при этом возбудимость ткани постепенно восстанавливается и сверхпороговый стимул, нанесенный в этот период может генерировать ПД;
IV – супернормальная возбудимость (вторичная или экзальфационная фаза) - следовой деполяризации, ткань становится более возбудимой, чем в исходном состоянии и даже подпороговый стимул способен вызвать ПД;
V – субнормальная возбудимость – следовой гиперполяризации, возбудимость ткани несколько снижена.

Рисунок 7. Изменение возбудимости мембраны при развитии потенциала действия.
2. Закон градиента раздражения (Дюбуа Реймон). Чем больше градиент раздражения, тем больше (до известных пределов) реакция живого образования.
За время действия медленно нарастающего стимула наступает приспособление ткани – аккомодация. Она связана с тем, что при возбуждении проницаемость для ионов натрия увеличивается на короткий промежуток времени, если в течение его раздражитель не достигает пороговой величины, то увеличивающаяся проницаемость для ионов калия инактивирует натриевую проницаемость и возбуждение не наступает. При этом происходит также сдвиг КУД с увеличением порогового потенциала.
3. Закон силы-времени (Лапик). Пороговая величина любого раздражителя находится в обратной зависимости от времени его действия, которая характеризуется математической кривой – гиперболой. Характер кривой свидетельствует о том, что подпороговые стимулы (меньше 1 реобазы) не вызовут возбуждение как долго бы они не действовали, в то же время очень сильный кратковременный стимул, длительность которого меньше полезного времени, также не вызовет возбуждение.
Сила постоянного тока, которая, действуя неопределенное время, вызывает возбуждение, называется реобазой.
Время, в течение которого ток в 1 реобазу вызывает возбуждение – полезное время.
Минимальное время, в течение которого ток силой в 2 реобазы вызовет возбуждение, называется хронаксией. Исследование этого показателя используется в неврологической и травматологической практике для изучения динамики восстановления в нервной или мышечной ткани после травмы.
Раздражители и раздражимость. На живой организм постоянно действуют различные раздражители (свет, звук, различные запахи и др.). Воздействие раздражителя на организм называется раздражением. Организм воспринимает раздражение благодаря особой способности – раздражимости. Раздражимость – это способность клеток, тканей усиливать или уменьшать активность в ответ на воздействие раздражителей. Условно раздражители можно подразделить на три группы: физические, химические и физико-химические. К физическим раздражителям относятся механические, электрические, температурные, световые звуковые. К химическим относятся гормоны, лекарственные вещества и др. К физико-химическим раздражителям относятся изменения осмотического давления и рН крови.
К действию одних раздражителей орган специально приспособлен. Такие раздражители называют адекватными. Неадекватными будут такие раздражители, к воздействию которых данная клетка или ткань не приспособлена. Так для глаза адекватным раздражителем будут световые лучи, а неадекватным звуковые волны.
По силе раздражители подразделяются на подпороговые, пороговые и надпороговые. Пороговый раздражитель характеризуется минимальной силой, достаточной для того чтобы вызвать минимальный специфический эффект в раздражаемой ткани. Подпороговый раздражитель вызывает лишь местную реакцию. Его силы недостаточно для вызывания специфического эффекта. Напротив, надпороговые раздражители обладают большой силой и вызывают самую большую реакцию.
Возбудимость и возбуждение. Некоторые ткани организма (нервная, мышечная) относятся к возбудимым т. е. они обладают способностью отвечать на раздражение - возбуждением. Возбуждение - это специфическая форма реагирования возбудимой клетки или ткани на действие раздражителя. Возбуждение характеризуется как специфическими, так и неспецифическими признаками. К специфическим признакам относят сокращение мышцы, выделение железой секрета. Неспецифические признаки возбуждения – это повышение обмена веществ, усиление теплопродукции изменение электрического состояния клеточной мембраны.
Биоэлектрические явления в возбудимых тканях. Биоэлектрические явления (животное электричество) было открыто в 1791 году итальянским ученым Л. Гальвани. Современные данные происхождения биоэлектрических явлений были получены в 1952 году А. Ходжикиным, А. Хаксли и Б. Катцем в исследованиях, проведенных с гигантским нервным волокном кальмара (диаметром 1 мм).
В покое проницаемость мембраны значительно выше для К, чем для Na. В силу своей высокой концентрации ионы Kстремятся выйти из клетки наружу. Сквозь мембрану они проникают на наружную поверхность клетки, но дальше уйти не могут. Крупные анионы клетки, для которых мембрана не проницаема, не могут следовать за калием, и скапливаются на внутренней поверхности мембраны, создавая здесь отрицательный заряд, который удерживает электростатической связью проскочившие через мембрану положительно заряженные ионы калия. Таким образом, возникает поляризация мембраны, потенциал покоя. По обе стороны образуется двойной электрический слой: снаружи из положительно заряженных ионов K, а внутри из отрицательно заряженных крупных анионов.

Потенциал действия (ПД). Потенциал покоя сохраняется до тех пор, пока не возникло возбуждение. Под действием раздражителя проницаемость мембраны для Naповышается. Поэтому Na сначала медленно, а затем лавинообразно устремляется внутрь клетки. Ионы натрия заряжены положительно, поэтому происходит перезарядка мембраны и ее внутренняя поверхность приобретает положительный заряд, а наружная - отрицательный. Таким образом происходит реверсия потенциала, изменение его на обратный знак (деполяризация). Он становится отрицательным снаружи и положительным внутри клетки. Однако повышение проницаемости мембраны для Na длится не долго. Она быстро снижается и повышается для K. Это вызывает усиление потока положительных ионов калия из клетки во внешний раствор. В итоге происходит реполяризация мембраны, ее наружная поверхность приобретает снова положительный заряд, а внутренняя – отрицательный.
Волна возбуждения. Волной возбужденияназывают всю совокупность последовательных изменений электрического состояния мембраны. К компонентам волны возбуждения относятся пороговый потенциал, потенциал действия и следовые потенциалы.
Законы раздражения. В 1907 году Л. Лапик для характеристики скорости возникновения возбуждения предложил регистрировать два параметра – силу раздражения и время его воздействия. Между силой раздражения и длительностью его действия существует обратно пропорциональная зависимость: чем больше сила раздражения, тем меньше длительность его действия, необходимая для возникновения возбуждения, и наоборот. О возбудимости ткани судят по величине реобазы. Реобаза – это наименьшая сила тока (или напряжения), способная при неограниченном времени вызвать возбуждение ткани. Она измеряется в единицах силы или напряжения тока. Чем меньше реобаза, тем более возбудима ткань.
О скорости возникновения возбуждения судят по величине хронаксии. Хронаксия – это наименьшее время, в течении которого необходимо воздействовать на ткань электрическим током, равным удвоенной реобазе, чтобы вызвать ее возбуждение. Она измеряется в единицах времени. Чем меньше хронаксия тем быстрее возникает возбуждение.
Лабильность. Парабиоз. Лабильность (или функциональная подвижность ткани) – это способность возбудимой ткани к воспроизведению потенциалов действия в соответствии с ритмом раздражения. Она была открыта Н.Е. Введенским в 1892 г. Мерой лабильности является наибольшее число потенциалов действия, которое ткань способна воспроизвести в 1 с в соответствии с частотой действующих раздражителей. Лабильность является величиной непостоянной. Она может понижаться или повышаться. Понижение лабильности может наступить вследствие утомления. Повысить ее можно путем физических упражнений.
При действии на участок нерва различных факторов (солевые растворы, электрический ток, механические раздражения и т.д.) Н.Е. Введенский установил, что лабильность измененного участка понижается. Именно это состояние стойкого нераспространяющегося возбуждения Н.Е. Введенский назвал парабиозом. Парабиоз имеет три стадии. Первая стадия – уравнительная, когда и сильные и слабые раздражения, нанесенные нормальному участку нерва, вызывают одинаковое сокращение мышцы. Вторая стадия – пародоксальная, когда сильные раздражения вызывают слабое сокращения, а слабые раздражения – более сильные сокращения, чем обычно. Третья стадия – тормозящая, когда ни сильные, ни слабые раздражения не вызывают сокращения. Установленные открытия Н.Е. Введенского сыграли большую роль в дальнейшем развитии физиологии.
1. Раздражители и раздражимость.
2. Возбудимость и возбуждение.
4. Потенциал действия.
Раздражители и раздражимость. На живой организм постоянно действуют различные раздражители (свет, звук, различные запахи и др.). Воздействие раздражителя на организм называется раздражением. Организм воспринимает раздражение благодаря особой способности – раздражимости. Раздражимость – это способность клеток, тканей усиливать или уменьшать активность в ответ на воздействие раздражителей. Условно раздражители можно подразделить на три группы: физические, химические и физико-химические. К физическим раздражителям относятся механические, электрические, температурные, световые звуковые. К химическим относятся гормоны, лекарственные вещества и др. К физико-химическим раздражителям относятся изменения осмотического давления и рН крови.
К действию одних раздражителей орган специально приспособлен. Такие раздражители называют адекватными. Неадекватными будут такие раздражители, к воздействию которых данная клетка или ткань не приспособлена. Так для глаза адекватным раздражителем будут световые лучи, а неадекватным звуковые волны.
По силе раздражители подразделяются на подпороговые, пороговые и надпороговые. Пороговый раздражитель характеризуется минимальной силой, достаточной для того чтобы вызвать минимальный специфический эффект в раздражаемой ткани. Подпороговый раздражитель вызывает лишь местную реакцию. Его силы недостаточно для вызывания специфического эффекта. Напротив, надпороговые раздражители обладают большой силой и вызывают самую большую реакцию.
Возбудимость и возбуждение. Некоторые ткани организма (нервная, мышечная) относятся к возбудимым т. е. они обладают способностью отвечать на раздражение - возбуждением. Возбуждение - это специфическая форма реагирования возбудимой клетки или ткани на действие раздражителя. Возбуждение характеризуется как специфическими, так и неспецифическими признаками. К специфическим признакам относят сокращение мышцы, выделение железой секрета. Неспецифические признаки возбуждения – это повышение обмена веществ, усиление теплопродукции изменение электрического состояния клеточной мембраны.
Биоэлектрические явления в возбудимых тканях. Биоэлектрические явления (животное электричество) было открыто в 1791 году итальянским ученым Л. Гальвани. Современные данные происхождения биоэлектрических явлений были получены в 1952 году А. Ходжикиным, А. Хаксли и Б. Катцем в исследованиях, проведенных с гигантским нервным волокном кальмара (диаметром 1 мм).
В покое проницаемость мембраны значительно выше для К, чем для Na. В силу своей высокой концентрации ионы Kстремятся выйти из клетки наружу. Сквозь мембрану они проникают на наружную поверхность клетки, но дальше уйти не могут. Крупные анионы клетки, для которых мембрана не проницаема, не могут следовать за калием, и скапливаются на внутренней поверхности мембраны, создавая здесь отрицательный заряд, который удерживает электростатической связью проскочившие через мембрану положительно заряженные ионы калия. Таким образом, возникает поляризация мембраны, потенциал покоя. По обе стороны образуется двойной электрический слой: снаружи из положительно заряженных ионов K, а внутри из отрицательно заряженных крупных анионов.
Потенциал действия (ПД). Потенциал покоя сохраняется до тех пор, пока не возникло возбуждение. Под действием раздражителя проницаемость мембраны для Naповышается. Поэтому Na сначала медленно, а затем лавинообразно устремляется внутрь клетки. Ионы натрия заряжены положительно, поэтому происходит перезарядка мембраны и ее внутренняя поверхность приобретает положительный заряд, а наружная - отрицательный. Таким образом происходит реверсия потенциала, изменение его на обратный знак (деполяризация). Он становится отрицательным снаружи и положительным внутри клетки. Однако повышение проницаемости мембраны для Na длится не долго. Она быстро снижается и повышается для K. Это вызывает усиление потока положительных ионов калия из клетки во внешний раствор. В итоге происходит реполяризация мембраны, ее наружная поверхность приобретает снова положительный заряд, а внутренняя – отрицательный.
Волна возбуждения. Волной возбужденияназывают всю совокупность последовательных изменений электрического состояния мембраны. К компонентам волны возбуждения относятся пороговый потенциал, потенциал действия и следовые потенциалы.
Законы раздражения. В 1907 году Л. Лапик для характеристики скорости возникновения возбуждения предложил регистрировать два параметра – силу раздражения и время его воздействия. Между силой раздражения и длительностью его действия существует обратно пропорциональная зависимость: чем больше сила раздражения, тем меньше длительность его действия, необходимая для возникновения возбуждения, и наоборот. О возбудимости ткани судят по величине реобазы. Реобаза – это наименьшая сила тока (или напряжения), способная при неограниченном времени вызвать возбуждение ткани. Она измеряется в единицах силы или напряжения тока. Чем меньше реобаза, тем более возбудима ткань.
О скорости возникновения возбуждения судят по величине хронаксии. Хронаксия – это наименьшее время, в течении которого необходимо воздействовать на ткань электрическим током, равным удвоенной реобазе, чтобы вызвать ее возбуждение. Она измеряется в единицах времени. Чем меньше хронаксия тем быстрее возникает возбуждение.
Лабильность. Парабиоз. Лабильность (или функциональная подвижность ткани) – это способность возбудимой ткани к воспроизведению потенциалов действия в соответствии с ритмом раздражения. Она была открыта Н.Е. Введенским в 1892 г. Мерой лабильности является наибольшее число потенциалов действия, которое ткань способна воспроизвести в 1 с в соответствии с частотой действующих раздражителей. Лабильность является величиной непостоянной. Она может понижаться или повышаться. Понижение лабильности может наступить вследствие утомления. Повысить ее можно путем физических упражнений.
При действии на участок нерва различных факторов (солевые растворы, электрический ток, механические раздражения и т.д.) Н.Е. Введенский установил, что лабильность измененного участка понижается. Именно это состояние стойкого нераспространяющегося возбуждения Н.Е. Введенский назвал парабиозом. Парабиоз имеет три стадии. Первая стадия – уравнительная, когда и сильные и слабые раздражения, нанесенные нормальному участку нерва, вызывают одинаковое сокращение мышцы. Вторая стадия – пародоксальная, когда сильные раздражения вызывают слабое сокращения, а слабые раздражения – более сильные сокращения, чем обычно. Третья стадия – тормозящая, когда ни сильные, ни слабые раздражения не вызывают сокращения. Установленные открытия Н.Е. Введенского сыграли большую роль в дальнейшем развитии физиологии.
2.5.1. Мембранные потенциалы
В возникновении и поддержании мембранного потенциала покоя основную роль играют два специальных белка. Один из них выполняет роль особого натрий-калиевого насоса, который за счет энергии АТФ активно перекачивает натрий из клетки наружу, а калий внутрь клетки. В результате концентрация ионов калия становится внутри клетки выше, чем в омывающей клетку жидкости, а ионов натрия – выше снаружи.
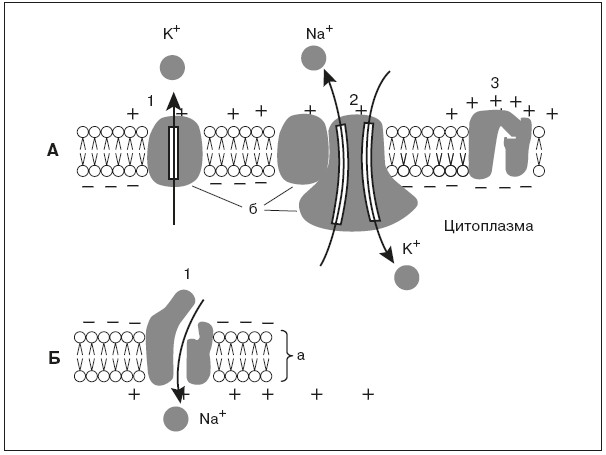
Рис. 1. Мембрана возбудимых клеток в покое (А) и при возбуждении (Б):
а – двойной слой липидов, б – белки мембраны;
Второй белок служит каналом утечки калия, через который ионы калия в силу диффузии стремятся выйти из клетки, где они содержатся в избытке. Ионы калия, выходя из клетки, создают положительный заряд на наружной поверхности мембраны. В результате внутренняя поверхность мембраны оказывается заряженной отрицательно по отношению к наружной. Таким образом, мембрана в состоянии покоя поляризована, т. е. имеется определенная разность потенциалов по обе стороны мембраны, называемая потенциалом покоя. Она равна для нейрона примерно минус 70 мВ, для мышечного волокна – минус 90 мВ. Измеряют мембранный потенциал покоя, вводя тонкий кончик микроэлектрода внутрь клетки, а второй электрод помещая в окружающую жидкость. В момент прокола мембраны и вхождения микроэлектрода внутрь клетки на экране осциллографа наблюдают смещение луча, пропорциональное величине потенциала покоя.
В основе возбуждения нервных и мышечных клеток лежит повышение проницаемости мембраны для ионов натрия – открывание натриевых каналов. Внешнее раздражение вызывает перемещение заряженных частиц внутри мембраны и уменьшение исходной разности потенциалов по обе стороны, или деполяризацию мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению натрия внутрь клетки. Эти реакции являются подпороговыми и вызывают лишь местные (локальные) изменения.
При увеличении раздражения изменения мембранного потенциала достигают порога возбудимости, или критического уровня деполяризации, – около 20 мВ, при этом величина потенциала покоя снижается примерно до минус 50 мВ. В результате открывается значительная часть натриевых каналов. Происходит лавинообразное вхождение ионов натрия внутрь клетки, вызывающее резкое изменение мембранного потенциала, которое регистрируется в виде потенциала действия. Внутренняя сторона мембраны в месте возбуждения оказывается заряженной положительно, а внешняя – отрицательно (рис. 1,Б).
Весь этот процесс чрезвычайно кратковременный. Он занимает всего около 1–2 мс, после чего ворота натриевых каналов закрываются. К этому моменту достигает большой величины медленно нараставшая при возбуждении проницаемость для ионов калия. Выходящие из клетки ионы калия вызывают быстрое снижение потенциала действия. Однако окончательное восстановление исходного заряда продолжается еще некоторое время. В связи с этим в потенциале действия различают кратковременную высоковольтную часть – пик (или спайк) и длительные малые колебания – следовые потенциалы. Потенциалы действия мотонейронов имеют амплитуду пика около 100 мВ и длительность около 1,5 мс, в скелетных мышцах – амплитуда потенциала действия 120–130 мВ, а длительность 2–3 мс.
Возникновение возбуждения (потенциала действия) возможно лишь при сохранении достаточного количества ионов натрия в окружающей клетку среде. Большие потери натрия организмом (например, с потом при длительной мышечной работе в условиях высокой температуры воздуха) могут нарушить нормальную деятельность нервных и мышечных клеток, снизив работоспособность человека. В условиях кислородного голодания тканей (например, при наличии большого кислородного долга во время мышечной работы) процесс возбуждения также нарушается из-за поражения (инактивации) механизма вхождения в клетку ионов натрия, и клетка становится невозбудимой. На процесс инактивации натриевого механизма влияет концентрация ионов Са 2+ в крови. При повышении содержания Са 2+ снижается клеточная возбудимость, а при дефиците Са 2+ возбудимость повышается и появляются непроизвольные мышечные судороги.
Мышца сердца (миокард), подобно нервной ткани и скелетным мышцам, принадлежит к возбудимым тканям. Это означает, что волокна миокарда обладают потенциалом покоя, отвечают на надпороговые стимулы генерацией потенциалов действия и способны проводить эти потенциалы без затухания (бездекрементно). Межклеточные соединения в сердце (к которым относятся, в частности, так называемые вставочные диски, выявляемые при микроскопии) не препятствуют проведению возбуждения. Мышечная ткань предсердий и желудочков ведет себя как функциональный синцитий: возбуждение, возникающее в каком-либо из этих отделов, охватывает все без исключения невозбужденные волокна. Благодаря этой особенности сердце подчиняется закону “все или ничего”:на раздражение оно либо отвечает возбуждением всех волокон, либо (если раздражитель подпороговый) не реагирует вовсе. Этим оно отличается от нервов и скелетных мышц, где каждая клетка возбуждается изолированно, и поэтому только в тех клетках, на которые наносят надпороговые раздражения, возникают потенциалы действия.
Автоматизм.Ритмические сокращения сердца возникают под действием импульсов, зарождающихся в нем самом. Если изолированное сердце поместить в соответствующие условия, оно будет продолжать биться с постоянной частотой. Это свойство называется автоматизмом. В норме ритмические импульсы генерируются только специализированными клетками водителя ритма (пейсмекера) и проводящей системы сердца.
Клетки, способные к автоматической генерации потенциала действия, образуют узлы автоматии (водители ритма, или пейсмекеры).
Выделяют три узла автоматии: 1) синоатриальный узел, расположенный в районе венозного входа в правом предсердии (узел Кис-Фляка). Именно этот узел является реальным водителем ритма в норме.
2) Атриовентрикулярный узел (Ашоффа-Тавара), который расположен на границе правого и левого предсердий и между правым предсердием и правым желудочком. Этот узел состоит из трех частей: верхней, средней и нижней.
3) Волокна Пуркинье — это конечная часть пучка Гиса, миоциты которой расположены в толще миокарда желудочков. Они являются водителями 3-го порядка, их спонтанный ритм — самый низкий, поэтому в норме являются лишь ведомыми, участвуют в процессе проведения возбуждения по миокарду.
Синоатриальный узел представляет собой соединительнотканный остов, в котором расположены специализированные мышечные клетки — в основном так называемые Р-клетки, собранные в агрегаты. Каждая из клеток этого узла способна к автоматии — благодаря высокой проницаемости для ионов натрия, и способна генерировать медленную диастолическую деполяризацию. До сих пор остается неясной причина, порождающая высокую проницаемость для ионов натрия и те особенности электрической активности, которые и приводят к генерации спонтанного потенциала действия.
Единый пейсмекерный ритм строится на основе интегративного взаимодействия всех моментов гетерогенного пейсмекера.
Роль водителя ритма первого порядка — синоатриального узла — огромна. Все регулирующие воздействия, меняющие ритм сердечной деятельности, оказывают свое влияние на сердце посредством воздействия на водитель ритма первого порядка. Если этот водитель выходит из работы, то ни симпатическая, ни парасимпатическая системы сами по себе не могут запустить деятельность сердца. В случае, когда синоатриальный узел повреждается ипри этом человеку успевают оказать квалифицированную медицинскую помощь, больному вживляют стимулятор, задающий самостоятельно ритм для работы сердца. Благодаря такому способу удалось сохранить жизнь многих пациентов.
Возбудимость клеток проводящей системы и рабочего миокарда имеет ту же биоэлектрическую природу, что и в поперечно-полосатых мышцах. Наличие заряда на мембране здесь также обеспечивается разностью концентраций ионов калия и натрия возле ее внешней и внутренней поверхности и избирательной проницаемостью мембраны для этих ионов. В покое мембрана кардиомиоцитов проницаема для ионов калия и почти непроницаема для ионов натрия. В результате диффузии ионы калия выходят из клетки и создают положительный заряд на ее поверхности. Внутренняя сторона мембраны становится электроотрицательной по отношению к наружной.
В клетках атипического миокарда, обладающих автоматией, мембранный потенциал способен спонтанно уменьшаться до критического уровня, что приводит к генерации потенциала действия. В норме ритм сердечных сокращений задается всего несколькими наиболее возбудимыми клетками синоатриального узла, которые называются истинными водителями ритма, или пейсмекерными клетками. В этих клетках во время диастолы мембранный потенциал, достигнув максимального значения, соответствующего величине потенциала покоя (60—70 мВ), начинает постепенно снижаться. Этот процесс называют медленной спонтанной диастолической деполяризацией. Она продолжается до того момента, когда мембранный потенциал достигает критического уровня (40—50 мВ), после чего возникает потенциал действия.
Частота возбуждения пейсмекерных клеток у человека составляет в покое 70—80 в минуту при амплитуде потенциала действия 70—80 мВ. Во всех остальных клетках проводящей системы потенциал действия в норме возникает под влиянием возбуждения, приходящего из синоатриального узла. Такие клетки называют латентными водителями ритма. Потенциал действия в них возникает раньше, чем их собственная медленная спонтанная диастолическая деполяризация достигает критического уровня. Латентные водители ритма принимают на себя ведущую функцию только при условии разобщения с синоатриальным узлом. Частота спонтанной деполяризации таких клеток у человека составляет 30—40 в минуту.
Спонтанная медленная диастолическая деполяризация обусловлена совокупностью ионных процессов, связанных с функциями плазматических мембран. Среди них ведущую роль играют медленное уменьшение калиевой и повышение натриевой и кальциевой проводимости мембраны во время диастолы, параллельно чему происходит падение активности электрогенного натриевого насоса. К началу диастолы проницаемость мембраны для калия на короткое время повышается, и мембранный потенциал покоя приближается к равновесному калиевому потенциалу, достигая максимального диастолического значения. Затем проницаемость мембраны для калия уменьшается, что и приводит к медленному снижению мембранного потенциала до критического уровня. Одновременное увеличение проницаемости мембраны для натрия и кальция приводит к поступлению этих ионов в клетку, что также способствует возникновению потенциала действия. Снижение активности электрогенного насоса дополнительно уменьшает выход натрия из клетки и, тем самым, облегчает деполяризацию мембраны и возникновение возбуждения.
Ионные механизмы возникновения потенциала действия кардиомиоцитов. Соотношения возбуждения, возбудимости и сократимости в различные фазы кардиоцикла. Экстрасистолы, механизм формирования компенсаторной паузы.
Клетки миокарда обладают возбудимостью, но им не присуща автоматия. В период диастолы мембранный потенциал покоя этих клеток стабилен, и его величина выше (80—90 мВ), чем в клетках водителей ритма. Потенциал действия в этих клетках возникает под влиянием возбуждения клеток водителей ритма, которое достигает кардиомиоцитов, вызывая деполяризацию их мембран.
Конечная реполяризация в клетках миокарда обусловлена постепенным уменьшением проницаемости мембраны для кальция и повышением проницаемости для калия. В результате входящий ток кальция уменьшается, а выходящий ток калия возрастает, что обеспечивает быстрое восстановление мембранного потенциала покоя. Длительность потенциала действия кардиомиоцитов составляет 300—400 мс, что соответствует длительности сокращения миокарда.
Экстрасистолы.Внеочередные раздражения, действующие по окончании рефракторной фазы и ведущие к внеочередной систоле, т. е. к экстрасистоле, могут появляться в различных участках проводящей системы сердца. Если внеочередное раздражение возникает в синусном узле, то оно приводит к преждевременному возникновению внеочередного цикла сердечной деятельности, протекающему с обычной последовательностью сокращений предсердий и желудочков. Характерно для такой синусной экстрасистолы, что пауза после этого внеочередного сокращения сердца короче, чем пауза между обычными сокращениями.
После экстрасистолы желудочков наступает удлиненная пауза между нею и следующей (очередной) систолой желудочков. Эта удлиненная пауза, называемая компенсаторной паузой, зависит от того, что экстрасистола, так же как нормальная систола, оставляет за собой рефракторную фазу. Очередной импульс, в обычном порядке зарождающийся в синусном узле, приходит в желудочки тогда, когда они оказываются в рефракторной фазе после экстрасистолы. Этот импульс не может вызвать систолы желудочков, и она наступит лишь в ответ на следующий импульс.
Возникновение распространяющегося возбуждения (ПД) возможно при условии, когда действующий на клетку раздражитель имеет некоторую минимальную (пороговую силу), иначе говоря, когда сила раздражителя соответствует порогу раздражения.
Порог раздражения – это та наименьшая величина раздражителя, которая действуя на клетку какое-то определенное время, способна вызвать максимальное возбуждение.
– наименьшая величина раздражителя, при действии которой потенциал покоя может сместиться до уровня критической деполяризации;
– критическая величина деполяризации клеточной мембраны, при которой активируется перенос ионов натрия внутрь клетки.

Рисунок 5. Возникновение местного потенциала при прохождении электрического тока, через участок нерва. Ток идет от анода к катоду (оба электрода – вне нерва) частично через пленку жидкости на поверхности нерва, а частично через оболочку нерва и в продольном направлении внутри волокна. Кривая внизу показывает вызываемое током изменения мембранного потенциала нервного волокна (по В. Катц)
Зависимость пороговой силы стимула от его длительности
Пороговая сила любого стимула в определенных пределах находится в обратной зависимости от его длительности. Эта зависимость, открытая Гоорвегом, Вейсом, Лапиком получила название кривой “сила – длительность” или “сила – время”. Кривая “силы времени” имеет форму, близкую к равносторонней гиперболе и в первом приближении может быть описана эмпирической формулой:
где I – сила тока, Т – длительность его действия, а и b – постоянные, определяемые свойствами ткани.
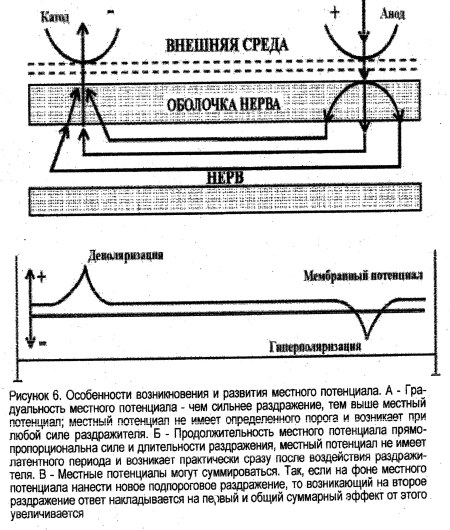
Рисунок 6. Особенности возникновения и развития местного потенциала. А – Градуальность местного потенциала – чем сильнее раздражение, тем выше местный потенциал; местный потенциал не имеет определенного порога и возникает при любой силе раздражителя. Б – Продолжительность местного потенциала прямопропорциональна силе и длительности раздражения, местный потенциал не имеет латентного периода и возникает практически сразу после воздействия раздражителя. В – Местные потенциалы могут суммироваться. Так, если на фоне местного потенциала нанести новое подпороговое раздражение, то возникающий на второе раздражение ответ накладывается на первый и общий суммарный эффект от этого увеличивается
Таким образом, го этой кривой вытекают два следствия:
1. Ток величиной ниже порога не вызывает возбуждения, как бы длительно он не действовал.
2. Какой бы сильный не был раздражитель, но если он действует очень короткое время, то возбуждение не возникает.
Реобаза – минимальная сила тока (или напряжение), способная вызвать возбуждение. Наименьшее время, в течение которого должен действовать стимул в одну реобазу, чтобы вызвать возбуждение – полезное время. Дальнейшее его увеличение не имеет значения для возникновения возбуждения.
Порог (реобаза) – величины непостоянные, зависят от функционального состояния клеток в покое, поэтому Лапик предложил определять более точный показатель – хронаксию.
Хронаксия – наименьшее время, в течение которого ток в две реобазы должен действовать на ткань, чтобы вызывать возбуждение.
Метод определения хронаксии – хронаксиметрия используется в клинике для диагностики повреждения нервных стволов и мышц.
Зависимость порога от крутизны нарастания раздражителя (аккомодация)
Порог раздражения имеет наименьшую величину при толчках электрического тока прямоугольной формы, когда сила нарастает очень быстро.
При уменьшении крутизны нарастания стимула ускоряются процессы инактивации натриевой проницаемости, приводящие к повышению порога и снижению амплитуды потенциалов действия. Чем круче должен нарастать ток, чтобы вызвать возбуждение, тем выше скорость аккомодации. Очень мала скорость аккомодации тех образований, которые склонны к автоматической деятельности (миокард, гладкие мышцы).
“Все” — на пороговые и сверхпороговые стимулы ответ максимальный и развивается потенциал действия; “ничего” – надо – пороговый стимул потенциал действия не развивается. Закон “все или ничего” установлен Боудичем в 1871 г. на мышце сердца: при подпороговой силе раздражения мышца сердца не сокращается, а при пороговой силе раздражения – сокращение максимально. При дальнейшем увеличении силы раздражения амплитуда сокращений не увеличивается.
Со временем была установлена и относительность этого закона. Оказалось, что “все” зависит от функционального состояния ткани (охлаждение, исходное растяжение мышцы и т. д.). С появлением микроэлектродной техники было установлено еще одно несоответствие: подпороговое раздражение вызывает местное, не распространяющееся возбуждение, следовательно, нельзя говорить, что допороговое раздражение не дает ничего. Процесс развития возбуждения подчиняется этому закону с уровня критической деполяризации, когда запускается лавинообразное поступление ионов калия в клетку.
Изменение возбудимости при возбуждении
Мера возбудимости – это порог раздражения. При местном, локальном, возбуждений возбудимость увеличивается. Потенциал действия сопровождается многофазными изменениями возбудимости.
1. Период повышенной возбудимости соответствует локальному ответу, когда мембранный потенциал достигает УКД, возбудимость повышена.
2. Период абсолютной рефрактерности соответствует фазе деполяризации потенциала действия, пику и началу фазы реполяризации, возбудимость снижена вплоть до полного отсутствия во время пика.
3. Период относительной рефрактерности соответствует оставшейся части фазы реполяризации, возбудимость постепенно восстанавливается к исходному уровню.
4. Супернормальный период соответствует фазе следовой деполяризации потенциала действия (отрицательный следовый потенциал), возбудимость повышена.
5. Субнормальный период соответствует фазе следовой гиперполяризации потенциала действия (положительный следовый потенциал), возбудимость снижена.

Рисунок 7. Изменения возбудимости нервного волокна в различные фазы потенциала действия и следовых изменений потенциала действия (по Б.И. Ходорову). Для наглядности длительность первых двух фаз на каждой кривой несколько увеличена. Пунктирной линией на рисунке А обозначает потенциал покоя, а на рисунке Б исходный уровень возбудимости
Закон лабильности или функциональной подвижности
Лабильность — скорость протекания физиологических процессов в возбудимой ткани. Например, можно творить о максимальной частоте раздражения, которую возбудимая ткань способна воспроизводить без трансформации ритма.
Мерой лабильности могут служить:
– длительность отдельного потенциала
– величина абсолютной рефрактерной фазы
– скорость восходящей и нисходящей фаз ПД.
Уровень лабильности характеризует скорость возникновения и компенсации возбуждения в любых клетках и уровень их функционального состояния. Можно измерять лабильность мембран, клеток, органов, причем, в системе из нескольких элементов (тканей, органов, образований) лабильность определяется по участку с наименьшей лабильностью:
Полярный закон раздражения (закон Пфлюгера)
Закон изменения мембранного потенциала при действии на возбудимые ткани постоянного электрического тока открыл Пфлюгер в 1859 г.
1. Постоянный ток проявляет свое раздражающее действие только в момент замыкания и размыкания цепи.
2. При замыкании цепи постоянного тока возбуждение возникает под катодом; при размыкании по анодом.
Изменение возбудимости под катодом.
При замыкании цепи постоянного тока под катодом (действуют допороговым, но продолжительным раздражителем) на мембране возникает стойкая длительная деполяризация, которая не связана с изменением ионной проницаемости мембраны, а обусловлена перераспределением ионов снаружи (они привносятся на электроде) и внутри – катион перемещается к катоду.
Вместе со смещением мембранного потенциала уровень критической деполяризации (УКД) смещается к нулю. При размыкании цепи постоянного тока под катодом мембранный потенциал быстро возвращается к исходному уровню, а УКД медленно, следовательно, порог увеличивается, возбудимость снижается и отмечается католическая депрессия Вериго. Таким образом, возбуждение возникает только при замыкании цепи постоянного тока под катодом.
Изменение возбудимости под анодом.
При замыкании цепи постоянного тока под анодом (действуют допороговым, но продолжительным раздражителем) на мембране развивается гиперполяризация за счет перераспределения ионов по обе стороны мембраны (без изменения ионной проницаемости мембраны) и возникающее за ней смещение уровня критической деполяризации в сторону мембранного потенциала. Следовательно, порог уменьшается, возбудимость повышается – анодическая экзальтация.
При размыкании цепи мембранный потенциал быстро восстанавливается к исходному уровню и достигает сниженного уровня критической деполяризации, генерируется потенциал действия. Таким образом, возбуждение возникает только при размыкании цепи постоянного тока под анодом. Сдвиги мембранного потенциала вблизи полюсов постоянного тока получили название электротонических. Сдвиги мембранного потенциала не связанные с изменением ионной проницаемости мембраны клетки называют пассивными.
Проведение возбуждения.
Потенциал действия – это волна возбуждения, распространяющаяся по мембранам нервных и мышечных клеток.
ПД обеспечивает передачу информации от рецепторов к нервным центрам и от них к исполнительным органам. Синоним ПД – нервный импульс или спайк. Сложная информация о действующих на организм раздражениях кодируется в виде отдельных групп потенциалов действия – рядов.
Согласно закону “все или ничего” амплитуда и длительность отдельных потенциалов действия постоянны, а частота и количество в ряду зависит от интенсивности раздражения. Такой способ кодирования информации и ее передачи является наиболее психоустойчивым.
В живых организмах информация может передаваться и гуморальным путем.
Преимущества ПД:
1. Информация более целенаправлена;
2. Передается быстро;
3. Адресат точно известен;
4. Информация может быть точнее закодирована.
ПД распространяется за счет местных токов, возникающих между возбужденным и невозбужденным участками. Из-за перезарядки мембраны во время генерации потенциала действия последний обладает способностью к самораспространению. Возникнув на одном участке, является стимулом для соседних. Наступающая после возбуждения в данном участке мембраны рефрактерностъ, обусловливает поступательное движение ПД.
Конкретные особенности распространения возбуждения связаны со строением мембраны клетки, нервных волокон. По мембранам мышечных клеток и в безмякотных нервных волокнах возбуждение распространяется непрерывно вдоль всей мембраны.
В волокнах, покрытых миелиновой оболочкой, потенциал действия может распространяться только скачкообразно (сальтаторно), перепрыгивая через участки волокна, покрытые шванновскими клетками, с одного перехвата Ранвье на другой.
Перехваты Ранвье представляют собой своеобразные станции ретрансляции, постоянно усиливающие сигнал, не давая ему угаснуть.
Причины сальтаторного проведения:
1. В перехватах Ранвье, свободных от миелина, сопротивление электрическому току минимально;
2. Порог раздражения в перехватах Ранвье минимальный;
3. Амплитуда ПД в каждом перехвате в 5 – 6 раз превышает порог в соседнем перехвате;
4. Велика плотность натриевых каналов на мембране перехвата.
Следовательно, возбуждение, возникающее в одном перехвате Ранвье, вызывает смещение электронов во внешней среде данного волокна и этого смещения достаточно, чтобы вызвать возбуждение в соседнем участке. Таким образом, скорость проведения возбуждения по нервному волокну зависит от диаметра волокон и наличия перехватов Ранвье.
Различают декрементное и бездекрементное распространение волны возбуждения.
ДЕКРЕМЕНТНОЕ проведение:
1. Наблюдается в безмиелиновых волокнах;
2. Скорость проведения невелика;
3. По мере удаления от места возникновения раздражающее действие местных токов постепенно уменьшается вплоть до полного угасания;
4. Свойственно волокнам, которые иннервируют внутренние органы, обладающие низкой функциональной активностью.
БЕЗДЕКРЕМЕНТНОЕ проведение:
1. ПД проходит весь путь от места раздражения до места реализации без затухания.
2. Характерно для миелиновых и тех безмиелиновых волокон, которые передают сигналы к органам, обладающими высокой реактивностью (сердце).
Распространение одиночного потенциала действия само по себе не требует энергетических затрат. Однако, восстановление исходного состояния мембраны и поддержание ее готовности к проведению нового импульса связано с затратой энергии.
Законы проведения возбуждения в нервах
Закон анатомической и физиологической непрерывности волокна.
Любая травма волокна нарушает проводимость. При действии новокаина (дикаина, кокаина) блокируются натриевые и калиевые каналы мембраны. Возникновение возбуждения и его проведение в этом случае становится невозможным.
Закон двустороннего проведения возбуждения
В целом организме по рефлекторной дуге возбуждение всегда распространяется в одном направлении: от рецептора к эффектору.
Причины:
1. Возбуждение всегда возникает при раздражении специфических рецепторов;
2. Рефрактерность во время возбуждения обусловливает поступательное движение;
3. В рефлекторной дуге возбуждение с одной нервной клетки на другую передастся в синапсах с помощью медиатора, который может выделяться только в одном направлении.
Закон изолированного проведения возбуждения в нервных стволах.
Передача возбуждения на большие расстояния невозможна из-за значительной потери тока во внеклеточной среде.
Физиология нейронов, глиальных клеток, рецепторов и синапсов
Классическая рефлекторная дуга состоит из:
– рецептора;
– афферентного пути (афферентного нейрона, который располагается в спинальном ганглии);
– нервного центра, где возбуждение с афферентного нейрона переходит на вставочную нервную клетку.
Затем возбуждение переходит на эффекторный орган (эффектор), в роли которого может выступать мышца. Многие нервные волокна покрыты глиальными клетками (миелиновая оболочка). Между этими Шванновскими клетками есть промежутки – перехваты Ранвье. Возбуждение с одного нейрона на другой и с мотонейрона на мышцу передается в синапсах с помощью медиатора.
Нервная клетка – структурная и функциональная единица ДНС, которая окружена клетками нейроглии.
Нейроглия (глиоциты) – совокупность всех клеточных элементов нервной ткани кроме нейронов.
В мозге взрослого человека 1150 – 200 млрд. глиальных клеток, что в 10 раз больше нервных.
Нейроглия:
1. макроглия:
– астроциты;
– олигодендроциты;
– эиендимоциты.
2. микроглия: глиальные макрофаги.
Астроциты составляют 45 – 60% серого вещества мозга. Покрывают 85% поверхности капилляров мозга (сосудистые ножки астроцитов), крупные отростки астроцитов контактируют с телами нейронов. Основная функция – трофическая.
Олигодендроциты образуют миелин в нервной системе к поддерживают его целостность.
Эпендимоциты – клетки, выстилающие стенки спинномозгового канала и всех желудочков головного мозга. Это граница между спинномозговой жидкостью (ликвор) и тканью мозга.
Функции нейроглии:
1. Опорная – вместе с сосудами и мозговыми оболочками образуют строму ткани мозга.
2. Трофическая – обеспечивают метаболизм нервных клеток (связь с кровеносными сосудами). В глиоцитах сосредоточен весь гликоген ЦНС.
3. Участие в интегративной деятельности мозга:
– формирование следов воздействия (память), а значит и условного рефлекса;
– без глиоцитов (блокада антиглиальным гамма-глобулином) меняется электрическая активность нейронов.
Особенности глиальных клеток:
1. Более чувствительны к ионным изменениям среды;
2. Высокая активность калий – натриевой АТФ-азы;
3. Высокая проницаемость для ионов калия;
4. Мембранный потенциал равен 90 мВ, у нейронов 60 – 80 мВ;
5. На раздражение отвечает только медленной деполяризацией не более 10 мВ;
6. Потенциал действия в глиальных клетках не генерируется.
Читайте также: