
Ведущая роль ядра в наследственности кратко
Обновлено: 05.07.2024
Свидетельство и скидка на обучение каждому участнику
Зарегистрироваться 15–17 марта 2022 г.
Дата: Класс: 9 Урок: 9 Учитель: Юсупова А.Т.
Тема: Клеточное ядро. Ведущая роль ядра в наследственности . Особенности клеток прокариот и эукариот.
Цель: организация деятельности учащихся по восприятию, осмыслению и первичному запоминанию новых знаний и способов деятельности.
- образовательная: Создать условия для формирования знаний учащихся о строении ядра;
- развивающая: Продолжить развитие умений работы с учебником, выявление особенностей органоидов связанных с выполняемой функцией.
- воспитательная: Формирование положительного отношения к занятиям.
Тип урока: изучение и первичное запоминание новых знаний и способов деятельности.
Форма организации урока: беседа, рассказ.
Оборудование: электронный учебник, презентация, таблица.
Ожидаемый результат: знать особенность строения ядра.
I . Организационный этап.
Обеспечить нормальную внешнюю обстановку для работы на уроке.
Психологически подготовить учащихся к общению.
II . Этап проверки домашнего задания.
Установить правильность, полноту и осознанность выполнения домашнего задания всеми учащимися.
Выявить пробелы в знаниях и способах деятельности учащихся.
Устранить в ходе проверки обнаруженные пробелы.
Индивидуальный опрос :
- Какие органоиды клетки вам известны?
- Как устроена мембрана клеток живых организмов?
- В чем сходство и различия в строении оболочек клеток разных организмов?
- Какими способами могут поступать вещества в клетку?
III . Этап актуализации субъективного опыта учащихся.
Обеспечить мотивацию учения школьников.
Обеспечить включение школьников в совместную деятельность.
Актуализировать субъективный опыт учащихся.
На слайде покажите и назовите органоиды клетки.
- Назовите основные части, характерные для любой клетки эукариот.
- Чем клетки эукариот отличаются от клеток прокариот?
IV . Этап изучения нового материала.
Обеспечить восприятие, осмысление и первичное запоминание учащимися изучаемого материала.
Обеспечить усвоение учащимися методики воспроизведения изучаемого материала.
Ядро – важнейшая структура клетки ( слайд 3 ).
*Объяснение учителя с использованием слада:
Итак, все клетки эукариот имеют хорошо сформированное ядро.
1) Ядро эукариот состоит:
А) Кариолема – Двойная ядерная мембрана, отделяет кариоплазму от цитоплазмы.
Б) Поры – отверстия в кариолеме, обеспечивают связь ядра со всей клеткой.
В) Кариоплазма – ядерный сок, содержащий органические и неорганические вещества.
Г) Хроматин – деспирализованные хромосомы.
Д) Ядрышки – органоиды ядра, образованные хроматином и белками.
Обычно клетки имеют по одному ядру, но бывают и исключения.
*Организация самостоятельной работы с текстом учебника ( слайд 4 ).
На странице 46 учебника, 3 абзац. Найдите примеры организмов имеющих 2 и более ядер.
- Клетки каких организмов, относящихся к эукариотам, не имеют ядер и почему?
- Каковы размеры клеточных ядер, от чего это может зависеть?
2) Особенности кариолемы ( слайд 5 ).
Объяснение учителя:
Содержимое ядра отделено от цитоплазмы ядерной оболочкой – кариолемой, состоящей из двух близко расположенных друг к другу мембран. Каждая мембрана толщиной 8 нм, расстояние между ними около 30 нм. Через определенные интервалы обе мембраны сливаются друг с другом, образуя отверстия, диаметром 70 нм – ядерные поры. Число пор непостоянно, оно зависит от размеров ядер и их функциональной активности.
3) Особенности кариоплазмы ( слайд 6 ).
* Объяснение учителя с элементами самостоятельной работы:
Внутренняя полость ядра заполнена кариоплазмой, состоящей из органических и неорганических веществ, а также хроматина и ядрышек.
Обратитесь к учебнику на странице 47 первый абзац, ответьте на вопросы:
- Что такое спирализация хромосом, когда она происходит?
- Какую роль в этом процессе играют белки?
*Объяснение учителя с элементами беседы:
Хромосомы – органоиды ядра, образующиеся в период деления клетки. Каждая хромосома состоит из одной нити ДНК и белков. Функция – носитель генетической информации.
Подумайте : Каково значение спирализации и деспирализации хромосом?
* Организация самостоятельной работы с учебником .
На страницах 32-33 найдите ответ на вопрос: Что представляет собой ядрышко, какова его функция в клетке?
Хромосомный набор клетки – кариотип .
*Объяснение учителя, сопровождающееся демонстрацией слайда:
Кариотип – уникален. И если количество хромосом у разных организмов может совпадать (например: картофель и шимпанзе имеют по 48 хромосом), то форма и размеры, а также структура хромосом будет отличаться.Диплоидный и гаплоидный наборы хромосом .
Клетки организма
Соматические (двойной набор хромосом – диплоидный).
Половые (одинарный – гаплоидный набор).
В диплоидном наборе, исходно половина хромосом досталась каждой клетке от материнской яйцеклетки и столько же хромосом – от сперматозоида отца.Парные, т.е. одинаковые хромосомы, получили название – гомологичных хромосом .
V . Этап закрепления новых знаний и способов деятельности.
Обеспечить закрепление учащимися знаний и способов действий, которые им необходимы для самостоятельной работы.
Задание 1. Фронтальный опрос:
- Почему ядро, считается носителем наследственной информации клетки? - Какие клетки не имеют ядер?- Каковы функции ядрышек?
Ядро — главная часть клетки. Роль ядра: участие в делении клетки, хранение и передача наследственных признаков организма, регуляция процессов жизнедеятельности в клетке, благодаря генетической информации, записанной в молекуле ДНК. В ядре каждой клетки содержится основная наследственная информация, необходимая для развития целого организма со всем разнообразием его свойств и признаков. Именно ядро играет центральную роль в явлениях наследственности.
Андрогенез наблюдается у отдельных видов животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть женское и мужское ядра не сливаются (Псевдогамия) и в дроблении участвует только мужское ядро.
Хромосомная теория наследственности Т.Г. Моргана. Основные положения. Ограниченность 3 закона Менделя. Кроссинговер и его значение для доказательства линейного расположения хромосом.
Хромосомная теория наследственности - теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности.
Детальная разработка хромосомной теории была произведена Т.Х. Морганом и его учениками (начиная с 1910 г.). Изучая наследование окраски глаз у плодовой мушки дрозофилы, Морган показал, что цвет глаз – признак, сцепленный с полом, и что по характеру его наследования ген, определяющий этот признак, должен находиться в половой хромосоме (Х-хромосоме). Так экспериментально была доказана связь конкретного гена с конкретной хромосомой. В дальнейшем было установлено, что многие признаки наследуются совместно – как один комплекс. Это означало, что контролирующие их гены образуют группы сцепления. Число таких групп сцепления оказалось равным гаплоидному числу хромосом, постоянному для каждого вида организмов.
Затем Морган обнаружил, что сцепленное наследование признаков может нарушаться в результате кроссинговера во время мейоза. На основании детального исследования сцепления генов и кроссинговера (на материале различных мутаций у дрозофилы) Морган и его сотрудники разработали методы определения взаимного положения различных генов на хромосомах и построения генетических карт хромосом. Хромосомная теория нашла подтверждение и дальнейшее развитие в открытии химической природы гена, выяснении строения хромосом и в других достижениях молекулярной генетики.
основные положения хромосомной теории наследственности(1911, Т.Морган)
1. основным материальным носителем наследственности являются хромосомы с локализованными в них генами
2. гены наследственно дискретны, относительно стабильны, но при этом могут мутировать
3. гены расположены в хромосомах линейно в особых участках-локусах на определенном расстоянии друг от друга
4. гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются, как правило, вместе
5. число групп сцепления равно гаплоидному набору хромосом и является постоянным для каждого вида
6. сцепление генов может нарушаться в результате кроссинговера-обмена участками гомологичных хромосом, (это и есть доказательство линейного расположения генов. Кстати, расстояние между соседними генами можно подсчитать. Например, между геном А и В расстояние 3М-морганиды, а между В и С-5 М. Значит, между А и С расстояние будет 8 М)
7.частота кроссинговера прямо пропорциональна расстоянию между генами
Значение теории в том, что она дала объяснение законам Меделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.

Закон независимого наследования (третий закон Менделя) — при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум (и более)парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). Когда скрещивались растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные горошины, наследование каждого из признаков следовало первым двум законам и в потомстве они комбинировались таким образом, как будто их наследование происходило независимо друг от друга. Первое поколение после скрещивания обладало доминантным фенотипом по всем признакам. Во втором поколении наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть 9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами и зелёными горошинами, 1:16 с белыми цветами и зелёными горошинами.
Если в скрещивании будет учитываться большее число генов, чем имеющееся у данного организма число пар гомологичных хромосом, то третий закон Менделя не осуществится. В этом случае вступят в действие другие закономерности, которые будут изложены позднее.
На первый взгляд это ограничение закона независимого сочетания признаков может создать впечатление ограниченности наследственной изменчивости в силу небольшого числа возможных комбинаций гамет. Однако это не так. Рассмотрим для примера возможный размах комбинативной изменчивости, возникающей в силу свободной комбинации гамет у человека. Допустим, что в каждой из 23 пар хромосом человека имеется только по одной паре аллелей. При этом число различных типов гамет выразится величиной 8 388 608, а их возможных комбинаций — 70 368 744 177 664.
Закономерное расщепление при полигибридном скрещивании осуществляется при соблюдении всех условий, о которых шла речь при анализе моногибридного скрещивания. К ним нужно добавить 2 следующих условия: 1) нахождение учитываемых генов в негомологичных хромосомах; 2) равновероятное образование всех сортов гамет на основе случайного расхождения негомологичных хромосом в мейозе.
Кроссинговер (другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между уи w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус.
Особенности молекулярного строения ДНК и РНК. Модель структуры ДНК Уотсона-Крика. Правило Э. Чаргаффа. Комплиментарность структуры ДНК. Жесткость молекулярной структуры и ее бесконечная вариабельность. Реакция на выявление ДНК и РНК. Проблема избыточности ДНК.
РНК – это полинуклеотиды, но состоят только из одной цепи, их мол.масса меньше, чем у ДНК. Кроме этого, они отличаются следующим: 1) количество РНК в клетке зависит от возраста, физиологического состояния, органной принадлежности клетки; 2) в мононуклеотидах РНК содержатся рибоза, вместо тимина урацил; 3) для РНК не характерны правила Чаргаффа; 4) в РНК больше минорных оснований, чем в ДНК, при этом в т-РНК количество минорных оснований приближается к 50. Все РНК синтезируются на ДНК, этот процесс называется транскрипцией.
В зависимости от локализации в клетке, функции различают 3 вида РНК: м-РНК (матричная, или информационная), транспортная – т-РНК, рибосомальная – р-РНК.
Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.).
Принцип комплементарности используется в синтезе ДНК. Это строгое соответствие соединения азотистых оснований, соединёнными водородными связями, в котором: А-Т и Г-Ц.
Ядро — главная часть клетки. Роль ядра: участие в делении клетки, хранение и передача наследственных признаков организма, регуляция процессов жизнедеятельности в клетке, благодаря генетической информации, записанной в молекуле ДНК. В ядре каждой клетки содержится основная наследственная информация, необходимая для развития целого организма со всем разнообразием его свойств и признаков. Именно ядро играет центральную роль в явлениях наследственности.
Андрогенез наблюдается у отдельных видов животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть женское и мужское ядра не сливаются (Псевдогамия) и в дроблении участвует только мужское ядро.
Хромосомная теория наследственности Т.Г. Моргана. Основные положения. Ограниченность 3 закона Менделя. Кроссинговер и его значение для доказательства линейного расположения хромосом.
Хромосомная теория наследственности - теория, согласно которой хромосомы, заключённые в ядре клетки, являются носителями генов и представляют собой материальную основу наследственности.
Детальная разработка хромосомной теории была произведена Т.Х. Морганом и его учениками (начиная с 1910 г.). Изучая наследование окраски глаз у плодовой мушки дрозофилы, Морган показал, что цвет глаз – признак, сцепленный с полом, и что по характеру его наследования ген, определяющий этот признак, должен находиться в половой хромосоме (Х-хромосоме). Так экспериментально была доказана связь конкретного гена с конкретной хромосомой. В дальнейшем было установлено, что многие признаки наследуются совместно – как один комплекс. Это означало, что контролирующие их гены образуют группы сцепления. Число таких групп сцепления оказалось равным гаплоидному числу хромосом, постоянному для каждого вида организмов.
Затем Морган обнаружил, что сцепленное наследование признаков может нарушаться в результате кроссинговера во время мейоза. На основании детального исследования сцепления генов и кроссинговера (на материале различных мутаций у дрозофилы) Морган и его сотрудники разработали методы определения взаимного положения различных генов на хромосомах и построения генетических карт хромосом. Хромосомная теория нашла подтверждение и дальнейшее развитие в открытии химической природы гена, выяснении строения хромосом и в других достижениях молекулярной генетики.
основные положения хромосомной теории наследственности(1911, Т.Морган)
1. основным материальным носителем наследственности являются хромосомы с локализованными в них генами
2. гены наследственно дискретны, относительно стабильны, но при этом могут мутировать
3. гены расположены в хромосомах линейно в особых участках-локусах на определенном расстоянии друг от друга
4. гены, расположенные в одной хромосоме, образуют группу сцепления и наследуются, как правило, вместе
5. число групп сцепления равно гаплоидному набору хромосом и является постоянным для каждого вида
6. сцепление генов может нарушаться в результате кроссинговера-обмена участками гомологичных хромосом, (это и есть доказательство линейного расположения генов. Кстати, расстояние между соседними генами можно подсчитать. Например, между геном А и В расстояние 3М-морганиды, а между В и С-5 М. Значит, между А и С расстояние будет 8 М)
7.частота кроссинговера прямо пропорциональна расстоянию между генами
Значение теории в том, что она дала объяснение законам Меделя, вскрыла цитологические основы наследования признаков и генетические основы теории естественного отбора.
Закон независимого наследования (третий закон Менделя) — при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум (и более)парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). Когда скрещивались растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные горошины, наследование каждого из признаков следовало первым двум законам и в потомстве они комбинировались таким образом, как будто их наследование происходило независимо друг от друга. Первое поколение после скрещивания обладало доминантным фенотипом по всем признакам. Во втором поколении наблюдалось расщепление фенотипов по формуле 9:3:3:1, то есть 9:16 были с пурпурными цветами и желтыми горошинами, 3:16 с белыми цветами и желтыми горошинами, 3:16 с пурпурными цветами и зелёными горошинами, 1:16 с белыми цветами и зелёными горошинами.
Если в скрещивании будет учитываться большее число генов, чем имеющееся у данного организма число пар гомологичных хромосом, то третий закон Менделя не осуществится. В этом случае вступят в действие другие закономерности, которые будут изложены позднее.
На первый взгляд это ограничение закона независимого сочетания признаков может создать впечатление ограниченности наследственной изменчивости в силу небольшого числа возможных комбинаций гамет. Однако это не так. Рассмотрим для примера возможный размах комбинативной изменчивости, возникающей в силу свободной комбинации гамет у человека. Допустим, что в каждой из 23 пар хромосом человека имеется только по одной паре аллелей. При этом число различных типов гамет выразится величиной 8 388 608, а их возможных комбинаций — 70 368 744 177 664.
Закономерное расщепление при полигибридном скрещивании осуществляется при соблюдении всех условий, о которых шла речь при анализе моногибридного скрещивания. К ним нужно добавить 2 следующих условия: 1) нахождение учитываемых генов в негомологичных хромосомах; 2) равновероятное образование всех сортов гамет на основе случайного расхождения негомологичных хромосом в мейозе.
Кроссинговер (другое название в биологии перекрёст) — явление обмена участками гомологичных хромосом во время конъюгации при мейозе.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды. Человек специально проводит гибридизацию с целью получения необходимых вариантов комбинаций для использования в селекционной работе.
Т. Морган предположил, что гены расположены в хромосомах линейно, а частота кроссинговера отражает относительное расстояние между ними: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме; чем реже кроссинговер, тем они ближе друг к другу.
Процент перекреста является функцией расстояния между генами. Поскольку расстояние между крайними генами у и bi равно сумме двух расстояний между уи w, w и bi, следует предположить, что гены расположены в хромосоме последовательно, т.е. линейно.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов в хромосоме строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место — локус.
Особенности молекулярного строения ДНК и РНК. Модель структуры ДНК Уотсона-Крика. Правило Э. Чаргаффа. Комплиментарность структуры ДНК. Жесткость молекулярной структуры и ее бесконечная вариабельность. Реакция на выявление ДНК и РНК. Проблема избыточности ДНК.
РНК – это полинуклеотиды, но состоят только из одной цепи, их мол.масса меньше, чем у ДНК. Кроме этого, они отличаются следующим: 1) количество РНК в клетке зависит от возраста, физиологического состояния, органной принадлежности клетки; 2) в мононуклеотидах РНК содержатся рибоза, вместо тимина урацил; 3) для РНК не характерны правила Чаргаффа; 4) в РНК больше минорных оснований, чем в ДНК, при этом в т-РНК количество минорных оснований приближается к 50. Все РНК синтезируются на ДНК, этот процесс называется транскрипцией.
В зависимости от локализации в клетке, функции различают 3 вида РНК: м-РНК (матричная, или информационная), транспортная – т-РНК, рибосомальная – р-РНК.
Комплемента́рность (в химии, молекулярной биологии и генетике) — взаимное соответствие молекул биополимеров или их фрагментов, обеспечивающее образование связей между пространственно взаимодополняющими (комплементарными) фрагментами молекул или их структурных фрагментов вследствие супрамолекулярных взаимодействий (образование водородных связей, гидрофобных взаимодействий, электростатических взаимодействий заряженных функциональных групп и т. п.).
Принцип комплементарности используется в синтезе ДНК. Это строгое соответствие соединения азотистых оснований, соединёнными водородными связями, в котором: А-Т и Г-Ц.
Итак, в ядре клеток заключены хромосомы, которые содержат ДНК - хранилище наследственной информации. Этим определяется ведущая роль клеточного ядра в наследственности. Данное важнейшее положение современной биологии не просто вытекает из логических рассуждений, оно доказано рядом точных опытов. Приведем один из них. В Средиземном море обитает несколько видов одноклеточных зеленых водорослей - ацетабулярий. Они состоят из тонких стебельков, на верхних концах которых располагаются шляпки. По форме шляпок различают виды ацетабулярий.
В нижнем конце стебелька ацетабулярии находится ядро.
У ацетабулярии одного вида искусственно удалили шляпку и ядро, а к стебельку подсадили ядро, извлеченное у ацетабулярии другого вида. Что же оказалось? Через некоторое время на водоросли с подсаженным ядром образовалась шляпка, характерная для того вида, которому принадлежало пересаженное ядро (рис. 17).
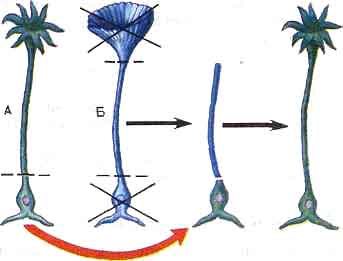
Рис. 17. Схема опыта с ацетабулярией. А и Б - разные виды ацетабулярий.
Хотя ядру принадлежит ведущая роль в явлениях наследственности, из этого, однако, не следует, что только ядро ответственно за передачу всех свойств из поколения в поколение. В цитоплазме также существуют органоиды (хлоропласты и митохондрии), содержащие ДНК и способные передавать наследственную информацию.
Таким образом, именно в ядре каждой клетки содержится основная наследственная информация, необходимая для развития целого организма со всем разнообразием его свойств и признаков. Именно ядро играет центральную роль в явлениях наследственности .
Наследственность является свойством клетки как системы в процессе ее работы и деления.
Нельзя считать, что для осуществления наследственности важно ядро и не важна цитоплазма, так как во всякой работающей системе все детали важны. Задача исследований состоит в выяснении функции и структуры отдельных компонентов клетки, которые в процессе эволюции приобрели свое специальное назначение.
Ядро можно назвать органом хранения наследственности, цитоплазму — органом осуществления наследственности.
Согласно принятому нами определению наследственности, все самовоспроизводящиеся структуры клетки должны обладать преемственностью. В этом аспекте и необходимо рассмотреть принципиальные отличия в структуре и функции ядра и цитоплазмы.
В общей форме можно сказать, что отличительные черты в структуре и функции ядра и цитоплазмы обусловлены их специализацией и различным назначением в деятельности клетки как системы. Поскольку органы цитоплазмы, как и ядро, содержат нуклеиновые кислоты и установлена цитоплазматическая дискретная наследственность, теперь принято различать ядерные, или хромосомные, гены и внеядерные, или экстрахромосомные, гены.
Для изучения роли ядра и цитоплазмы в наследственности применяются различные методы. К ним относятся:
- метод замещения ядра;
- метод реципрокных и возвратных скрещиваний;
- метод получения цитоплазматических мутаций и изучения поведения их в поколениях.
Одним из наиболее надежных в настоящее время вариантов метода замещения ядра является разрушение ядра яйцеклетки одного вида с последующим оплодотворением ее сперматозоидом другого вида. При развитии таких зигот должны получаться гаплоидные андрогенное особи, поскольку они несут одно только отцовское ядро и материнскую цитоплазму. Но гаплоидные зиготы погибают на ранней эмбриональной или личиночной стадиях, и поэтому долгое время таким путем не удавалось получить взрослых форм андрогенного происхождения. Лишь недавно Б. Л. Астаурову и В. П. Остряковой-Варшавер удалось получить диплоидных андрогенных гибридов при скрещивании двух видов шелкопряда: Bombyx mori и B. mandarina.
У шелкопряда, как и у других бабочек, гетерогаметным полом являются самки, а гомогаметным самцы. Кроме того, у шелкопряда, как и у многих насекомых, наблюдается полиспермия. Хромосомы тутового шелкопряда легко метятся определенными генами, с которым можно различать в потомстве признаки обоих видов. В скрещивании В. mandarina X В. mori самец был маркирован тремя рецессивными генами, находящимися в разных хромосомах: ch — шоколадный, определяющий желто-коричневую окраску личинок, р — ген белой окраски бабочек и ml — ген молочно-белой окраски гиподермы гусениц. Самка была отмечена соответственно тремя доминантными аллелями этих генов, определяющими черный цвет личинок, темную окраску бабочек и серый цвет гусениц. Следовательно, при указанном скрещивании у всех нормальных гибридных потомков должны быть черные личинки, серые гусеницы и тёмные бабочки, а у андрогенных особей должны проявиться рецессивные гены: личинки желто-коричневого цвета, гусеницы и бабочки светлой окраски. В этом исследовании разрушение ядра яйцеклетки после проникновения в нее сперматозоидов производилось в момент второго мейотического деления тепловым шоком, а именно воздействием температуры 40° в течение 120—135 мин. каждой кладки одну часть яиц (грены) подвергали такому воздействию, другую — оставляли в качестве контроля. Так как в опыт-группе материнское ядро убивалось, то развитие могло идти только при условии, если сливались два мужских пронуклеуса образуя одно диплоидное ядро (диспермический андрогенез). В результате все развивающиеся особи должны быть мужского пола, так как самцы имеют две Х-хромосомы; кроме того, все потомки должны иметь рецессивные признаки, поскольку хромосомы сами были мечены рецессивными генами.
Схема получения диплоидных андрогенных особей у Bombyx методом теплового воздействия
В опыте было получено большое число андрогенных личинок и несколько половозрелых самцов, имевших сперматозоиды, способные к оплодотворению. Результаты опыта можно рассматривать как прямое доказательство ведущей роли ядра в наследственности и отсутствия заметного влияния материнской цитоплазмы.
Г. Гемерлинг провел опыт с замещением ядра у зеленой водоросли рода Acetabularia. Он взял два вида этого рода mediterranea и A. wetfsteinii, различающихся формой шляпки. Эти водоросли на определенном этапе жизненного цикла имеют по одному ядру, находящемуся в одном из ризоидов.
Влияние ядра на форму шляпки у Acetobularia
Отрезая ризоиды, содержащие ядро, и затем сращивая их с отрезками стебельков так, чтобы ядро одного вида соединялось с плазмой другого, можно было наблюдать влияние ядра в чужой плазме на развитие новой шляпки. Оказалось, что форма шляпки развивается соответственно тому виду, которому принадлежит пересаженное ядро.
Микрохирургическим путем удавалось замещать ядра у разных видов амеб. И в этом случае признаки вида-реципиента, сохранявшие свою цитоплазму, проявлялись лишь в первые дни после операции, а затем обнаруживались признаки вида, которому принадлежало пересаженное ядро.
Читайте также: