
Строение древесины хвойных деревьев смолоносная система кратко
Обновлено: 05.07.2024
Древесина хвойных пород отличается довольно простым и правильным строением.
В центре ствола хвойных пород находится так называемая сердцевина, имеющая вид неправильного круга диаметром 1—4 мм. Сердцевина состоит из паренхимных тонкостенных и одревесневших клеток. В молодом деревце или вершинном побеге, где еще нет законченной дифференциации тканей древесины, клетки сердцевины выполняют функции запасающих тканей и содержат крахмал, жиры и другие органические продукты. По мере роста дерева и выделения специализированных тканей, надобность в запасающих клетках сердцевины отпадает, они отмирают и заполняются воздухом.
На поперечном и продольном разрезах видно, что у сосны, лиственницы и кедра центральная часть ствола отличается от периферической более темным цветом, а в свежесрубленном состоянии, кроме того, и меньшим содержанием воды.
Темноокрашенная центральная часть древесины называется ядром, а периферическая светлая — заболонью.
У ели и пихты центральная часть ствола отличается от периферической только меньшим содержанием воды и называется спелой древесиной.
Ядровая и спелая часть древесины у растущих деревьев выполняет механические функции, не участвуя в обмене веществ.
С возрастом деревьев процентное содержание ядра или спелой древесины увеличивается.
В условиях Севера протяжение заболони по радиусу деревьев в возрасте 140 лет (на высоте 1,3 м) неодинаково для различных пород.
Ядро и спелая древесина образуются тогда, когда окаймленные поры трахеид закупориваются и выключаются из водопроводящей системы дерева, а клеточные оболочки пропитываются особыми ядерными веществами.
Водопроводящие и механические функции у хвойных деревьев выполняют трахеиды, они являются основным элементом древесины и занимают 90—95% общего ее объема.
Трахеиды расположены радиальными рядами и на поперечном срезе имеют вид четырехугольных или шестиугольных клеток, а на радиальном и тангентальном разрезах — вид волокон с косо срезанными концами. Радиальные ряды состоят из повторяющихся в каждом годичном кольце двух типов трахеид — ранних и поздних. Ранние трахеиды, выполняя проводящие функции, имеют широкую внутреннюю полость и тонкие стенки с многочисленными порами. Поздние трахеиды, образующие так называемую позднюю древесину, выполняют механические функции, поэтому имеют утолщенные стенки и сравнительно узкие полости.
Характерная особенность трахеид — окаймленные поры, сгруппированные преимущественно на радиальных стенках у концов, которыми каждая трахеида вклинивается между соседними, образуя плотное соединение.
Окаймленные поры — это не простые отверстия, соединяющие полости двух соседних клеток, с поверхности они прикрыты тонким первичным слоем клеточной оболочки. Поры соседних трахеид всегда находятся одна против другой и разделены общей для двух клеток срединной пластинкой (мембраной). Следовательно, окаймленная пора образуется двумя соседними трахеидами. Мембрана окаймленной поры в центре имеет утолщение, называемое торусом. Над первичным слоем и торусом вторичные слои соседних клеток нависают в виде купола, как бы окаймляя их, отсюда и название — окаймленная пора.
При выходе из полости клеток диаметр пор примерно равен 4—5 μ, затем сильно расширяется и в месте примыкания к срединной пластинке достигает 12—25. Диаметр торуса всегда несколько превышает внутренний диаметр поры.
Первичные слои клеток, образующие мембрану окаймленной поры, в своей неутолщенной части имеют тончайшие отверстия, облегчающие проникновение веществ из клетки в клетку. В живых клетках эти мельчайшие отверстия заполнены плазматическими нитями, при помощи которых плазма всех живых клеток организма соединена в одно целое.
При отклонении мембраны в ту или другую сторону торус закрывает выходное отверстие из полости клетки, прекращая этим дальнейшее функционирование поры.
У трахеид, расположенных в зоне ядра или спелой древесины, подавляющее большинство пор закупорено торусом, и они выключаются из водопроводящей системы. В заболони число пор, закрытых торусом, незначительно, и практически они не влияют на передвижение воды в древесине.
Число окаймленных пор в трахеидах довольно велико. Так, количество их у ранних трахеид 80—100, а у поздних — 10—20.
Размеры трахеид у хвойных деревьев изменяются не только в зависимости от породы, но также и от возраста, условий местопроизрастания и других факторов. Основываясь на обширных исследованиях ученых, можно принять для сосны, ели, лиственницы и пихты следующие размеры трахеид: длина 2—4 мм, ширина или наружный диаметр 0,2—0,04 мм, толщина стенок 0,005—0,008 мм. У поздних трахеид длина и толщина стенок больше, а ширина меньше.
Клеточные стенки трахеид состоят из целлюлозы, лигнина, гемицеллюлоз и полиуронидов. Полости этих клеток заполнены водой и воздухом.
Вторым элементом хвойных пород являются сердцевинные лучи, идущие между трахеидами в строго радиальном направлении.
Длина сердцевинных лучей увеличивается с каждым новым вегетационным периодом, по мере удаления камбиального слоя от центра ствола.
Сердцевинные лучи состоят из паренхимных живых клеток и предназначены для хранения питательных веществ. Клетки сердцевинных лучей значительно меньше трахеид, длина их примерно 0,04—0,08 мм, ширина 0,01—0,03 мм. Сердцевинные лучи у хвойных пород по ширине обычно образуются одним рядом клеток, если же в сердцевинном луче залегает горизонтальный смоляной ход, — то и несколькими рядами.
По высоте в луче всегда несколько рядов клеток: у сосны, кедра, лиственницы и ели верхний и нижний ряды образованы горизонтальными трахеидами с мелкими окаймленными порами, внутренние ряды — паренхимными клетками с простыми порами.
Выполняя функции запасающих тканей, сердцевинные лучи должны быть постоянно связаны с лубом, по которому идут питательные вещества от листьев. Эта связь осуществляется через камбий, который образует сердцевинные лучи как в сторону древесины, так и в сторону луба.
Стенки клеточной оболочки сердцевинных лучей сравнительно тонки и состоят в основном из целлюлозы, что делает их непрочными в механическом отношении. Полости клеток заполнены протопластом, содержащим в себе живую часть — протоплазму и ядро, а также неживые включения вроде зерен крахмала, жиров и т. п.
Протоплазма клеток сердцевинных лучей через ультрамикроскопические отверстия в мембране пор соединена в одно целое, а также соединена с протоплазмой клеток камбиального слоя и живых клеток луба. Благодаря этому клетки сердцевинных лучей могут выполнять функцию запасающих тканей и передавать запасы органических веществ камбию и другим живым клеткам.
В ядровой и спелой древесине сердцевинные лучи, как и все другие ткани, отмирают и перестают выполнять запасающие функции.
У хвойных пород, кроме сердцевинных лучей, есть еще одна ткань, которая выполняет запасающие функции, — это древесная паренхима. Клетки древесной паренхимы разбросаны между трахеидами или сгруппированы около смоляных ходов, стенки их сравнительно тонки и состоят из целлюлозы, гемицеллюлоз и лигнина.
В ядровой и спелой древесине клетки паренхимы отмирают и заполняются воздухом и водой. Количество древесной паренхимы в хвойных породах незначительно и обычно не превышает 1 % всей древесины.
Характерной особенностью древесины хвойных пород являются смоляные ходы, за исключением пихты, у которой они наблюдаются только иногда и то единично. Каждый ход состоит из канала, образуемого паренхимными клетками путем их дифференциации.
Смоляные ходы бывают вертикальные, тянущиеся параллельно оси ствола, и горизонтальные, проходящие по сердцевинным лучам.
Диаметр вертикальных смоляных ходов в среднем у сосны равен 0,1 мм, у ели и лиственницы — 0,06 мм. Длина ходов колеблется от 10 до 100 см.
Горизонтальные смоляные ходы имеют диаметр 0,03—0,04 мм и длину не выше сердцевинного луча. На 1 см 2 поперечного среза сосны и ели насчитывается от 200 до 300 вертикальных смоляных ходов.
Горизонтальные смоляные ходы своими каналами всегда соединены с вертикальными, так что вся система образует одно целое. Такое строение дает возможность обильно заливать смолой ранения в любой части ствола.
Кроме того, через горизонтальные смоляные ходы живые клетки соединены с другими живыми тканями дерева.
В ядровой и спелой древесине горизонтальные смоляные ходы зарастают тиллами, часть ходов в ядровой древесине перестает функционировать и отключает от общей системы вертикальные смоляные ходы.
Смоляные ходы в заболони, вследствие смоляного давления, способны выделять терпентин непосредственно после их перерезания. Смоляные ходы ядровой древесины этим свойством не обладают, и терпентин в них находится как бы в хранилище.
При растрескивании древесины или других разрушениях смоляных ходов в ядре терпентин, разливаясь, не кристаллизуется, а пропитывает мертвые клетки. Как показывают исследования, смола и терпены обладают слабой токсичностью по отношению к дереворазрушающим грибам, поэтому большая стойкость смолистой древесины против гниения объясняется не столько антисептическими, сколько физическими их свойствами.
Смоляные ходы располагаются, главным образом, в позднем слое годичного кольца.
Обобщая все вышеизложенное, можно сделать следующее заключение о строении древесины хвойных пород.
Ткани, составляющие древесину, располагаются в стволе от периферии к центру в определенном порядке. Снаружи ствол покрывает пробковый слой коры, состоящий из мертвых пробковых и пробковидных клеток, очень устойчивых против микроорганизмов и насекомых. Клетки пробкового слоя у сосны и лиственницы содержат воды около 20%, у ели — 30%. К пробковым клеткам примыкает очень тонкий слой феллодермы, состоящий из клеток пробкового камбия, а также паренхимных клеток коры. Затем идет лубяной слой, состоящий из ситовидных трубок, сердцевинных лучей, лубяных волокон и лубяной паренхимы.
В феллодерме и лубе много живых клеток, заполненных плазмой и содержащих жиры и крахмал. Ситовидные трубки заполнены органически ми веществами, идущими от листьев. Все это делает феллодерму и луб весьма благоприятным питательным материалом для развития микроорганизмов и насекомых. Клетки луба и феллодермы имеют в среднем влажность у ели 141%, у сосны 152, у лиственницы 200% от абсолютно сухого веса; при высыхании они изменяют объем, что приводит к растрескиванию луба и постепенному отпаду коры.
За лубом идет камбиальный слой, состоящий из живых клеток, при жизни очень стойких против грибов, а в отмершем-состоянии, после рубки дерева, являющихся хорошей питательной средой для микроорганизмов.
За камбием следуют слои собственно древесины. В заболони, площадь которой колеблется в широких пределах в зависимости от диаметра деревьев, 50—60% клеток первого годичного слоя, считая от камбия, — живые, содержат протопласт, а также некоторые другие органические вещества. В последующих годичных кольцах заболони количество живых клеток резко уменьшается, они сохраняются в сердцевинных лучах, в смоляных ходах и в древесной паренхиме около лучей и смоляных каналов.
Следовательно, весь поверхностный слой заболони — это весьма хорошая питательная среда для микроорганизмов, в последующих слоях эти благоприятные условия для развития грибов остаются только в сердцевинных лучах и клетках древесной паренхимы.
Ядровая и спелая древесина состоит целиком из мертвых клеток, полости которых заполнены воздухом, а стенки пропитаны смолистыми и ядерными веществами. Поэтому ядро и спелая древесина могут служить питательной средой только для более высокоорганизованных видов грибов — базидиомицетов.
В круглых лесных сортиментах, непосредственно после валки деревьев, древесина по всей боковой поверхности прикрыта корой и обнажена только с торцов. От поражения ее грибами до некоторой степени механически предохраняет пробковый слой коры, а камбиальный слой до отмирания клеток сохраняет способность противостоять грибам и препятствует их проникновению в древесину. Влажность заболони также неблагоприятна для развития грибов.
Таким образом, в теплое время года, в первые 10—15 дней после рубки, древесина в круглых лесных сортиментах не подвергается серьезным изменениям и при надлежащих защитных мероприятиях может долго сохранять свое качество. Это в известной мере относится к пиломатериалам и другим сортиментам, о чем будет подробно сказано в последующих разделах.
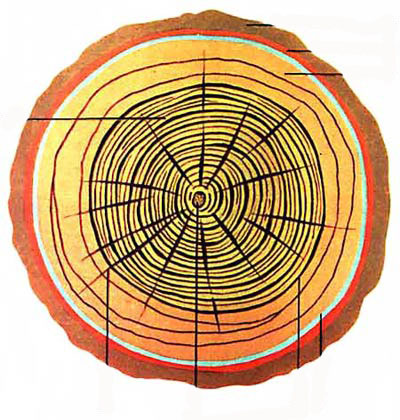
Хвойные породы возникли в эволюции раньше лиственных и имеют наиболее простое однородное анатомическое строение древесины, состоящей практически полностью из клеток одного типа. Главными анатомическими элементами древесины хвойных пород служат прозенхимные клетки — трахеиды, составляющие до 91. 95% ее объема. Это длинноватые клетки со стенками разной толщины. В стволе растущего дерева лишь последний годичный слой содержит живые трахеиды, которые отмирают к зиме. В поперечном сечении трахеиды чаще всего имеют прямоугольную форму, а время от времени пяти- либо шестиугольную. Концы традиционно кососрезанные с заостренными либо закругленными кончиками. В дереве трахеиды размещены главным образом вертикально (вдоль оси ствола). Длина трахеид обычно составляет 1,6. 5,5 мм (с колебаниями для представителей отдельных семейств от 0,7 до 11 мм) при ширине 0,02. 0,09 мм.
Древесину хвойных пород применяют в целлюлозно-бумажном производстве как длинноволокнистое сырье. Ранние трахеиды наиболее широкие, имеют тонкие стенки и широкие полости (соотношение длины к ширине около ста). Они выполняют проводящие функции. Поздние трахеиды наиболее узкие (соотношение длины к ширине примерно двести), имеют толстые стенки и узкие полости. Они выполняют механические функции. Размеры трахеид зависят от древесной породы и условий произрастания дерева.
Вторым анатомическим элементом являются паренхимные клетки, образующие живую ткань — паренхиму, главным образом лучевую. Ее клетки образуют сердцевинные лучи — ряды из паренхимных клеток, идущие горизонтально по радиусам ствола. Лучи могут состоять лишь из паренхимных клеток (гомогенные лучи), или содержать не считая паренхимных клеток горизонтальные лучевые трахеиды (гетерогенные лучи). У хвойных пород сердцевинные лучи узкие, однорядные (кроме лучей, в которых образуются горизонтальные смоляные ходы), а их высота по числу клеток колеблется. Горизонтальные (лучевые) трахеиды по размерам близки к паренхимным клеткам, но малочисленны по сравнению с последними. В отличие от паренхимных клеток лучевые трахеиды имеют окаймленные поры (см ниже). Сердцевинные лучи проводят растворы питательных веществ в горизонтальном направлении. В древесине некоторых хвойных пород в маленьких количествах (например, у сосны 0,6%) содержится вертикальная (осевая, или тяжевая) паренхима, ряды клеток которой проходят вдоль ствола.
Паренхима выполняет наряду с проводящей запасающую функцию В ее клетках хранятся резервные питательные вещества, содержатся экстрактивные вещества и минеральные. У хвойных деревьев часть запасов питательных веществ хранится также в хвое и поэтому доля паренхимных клеток довольно мала (3,5. 6%). Исключение составляет древесина лиственницы, сбрасывающей хвою на зиму, содержащая около 10% паренхимных клеток.
В древесине некоторых хвойных пород (сосна, лиственница, ель) находятся также эпителиальная паренхима, образующая смоляные ходы (смоляные каналы), и сопровождающая их паренхима. Смоляные ходы -это межклеточные каналы, заполненные живицей (смолой). Распознают вертикальные и горизонтальные смоляные каналы, образующие единую смолоносную систему. Вертикальные каналы размещаются чаще в поздней зоне годичного кольца и образуются 3-мя слоями клеток: внутренним выстилающим слоем живых эпителиальных клеток; слоем мертвых клеток, заполненных воздухом; слоем живых клеток сопровождающей паренхимы, с течением времени отмирающих. Диаметр вертикальных смоляных ходов составляет 0,10. 0,14 мм. Их можно заметить и невооруженным глазом.
Горизонтальные смоляные каналы проходят в многорядных сердцевинных лучах. Они образованы живыми эпителиальными клетками и мертвыми паренхимными клетками. Эти ходы наблюдают под микроскопом. Диаметр смоляных каналов зависит от породы дерева. Так, в древесине сосны каналы имеют большие размеры, чем в древесине ели. Смолоносная система служит в дереве защитной системой, позволяющей переносить экстремальные условия обитания, и в том числе обеспечивает защиту от повреждения дерева насекомыми и иными живыми организмами.
Клетки древесины сообщаются между собой через поры. Поры -это неутолщенные участки клеточной стенки. Пора не является свободным отверстием, потому что в ней имеется узкая мембрана (первичная стенка и межклеточное вещество), пронизанная мелкими отверстиями. В живых клетках через эти отверстия проходят тонкие нити цитоплазмы, которые соединяют содержимое живых клеток в одно целое. Поре в оболочке одной клетки соответствует пора соседней клетки, другими словами образуется пара пор.
Различают простые, окаймленные и полуокаймленные поры (пары пор). Простые поры образуются в стенках 2-х смежных паренхимных клеток, а окаймленные поры — в стенках 2-х смежных трахеид, располагаясь преимущественно на радиальных стенках у концов трахеид. Поздние трахеиды по сравнению с ранними имеют наименьшее число пор меньших размеров (щелевидные поры). У окаймленной поры мембрана имеет в центре утолщение — торус, играющий роль клапана, который может перекрывать пору. Структура торуса отличается от структуры мембраны. Окаймление образуется нависающим выступом вторичной стенки. Оно может быть выражено четко либо слабо заметно.
Трахеиды с паренхимными клетками сердцевинных лучей сообщаются через полуокаймленные поры в так называемых полях перекреста. Форма, размер и число пор в поле перекреста служат диагностическими признаками при определении хвойных древесных пород.
X. Характерные собенности коры, корки древесины хвойных древесных растений
10.1. Строение коры хвойных растений
Корой называют совокупность тканей стебля и корня, лежащих наружу от камбия, независимо от их происхождения, выполняемой функции и анатомического строения. У многолетних древесных растений различают внешнюю кору, которая называется коркой (ритидом) и внутреннюю, состоящую из вторичного луба, или вторичной флоэмы. Образование корки связано с возрастными изменениями вторичного луба, обусловленными активной деятельностью паренхимных клеток, в которых увеличивается содержание крахмала, таннидов и кристаллов оксалата кальция. Клетки способны метаморфизироваться в склереиды, приобретая, таким образом, механическую функцию.
Лубяная паренхима представляет собой также меристематический резерв коры, за счет которого возникает феллоген и осуществляется делатация (расширение тканей). Способность к делатации присуща как клеткам тяжевой, так и лучевой паренхимы.
Делатация в молодых побегах хвойных происходит за счет паренхимы первичной коры, а в старой коре делегирует тяжевая паренхима, общая поверхность клеток которой на поперечных срезах коры увеличивается в десятки раз по сравнению с размерами поверхности лубяной паренхимы в проводящей флоэме. Род Pinus отличается от других представителей семейства Сосновые отсутствием механических элементов во вторичном лубе. У сосны обыкновенной, у сосны черной, австрийской механической функции лишена также первичная кора молодых побегов, состоящая из паренхимы.
В однолетних побегах клетки пробкового камбия вычленяются вследствие периклинальных делений субгиподермальных клеток. Образовавшаяся перидерма отделяет от первичной коры к стеблю основания листьев, которые постепенно сбрасываются, и стебель становится округлым в поперечном сечении. Рост в толщину мало отражается на строении первичной коры, клетки претерпевают сравнительно небольшое поверхностное растяжение и редко делятся в радиальном направлении. При разрушении клеток в коре возникают воздушные полости.
На восьмом - десятом году жизни побега в первичной коре, на внутренней стороне от вертикальных смоляных каналов, закладывается феллоген, и после развития перидермы образуется типичная кольцевая корка. Следующие полоски феллогена возникают во вторичном лубе. Протяженность дут пробкового камбия на поперечных срезах коры постепенно уменьшается, корка становится чешуйчатой. Ритмичность камбиальной деятельности, обуславливающая тангенциальную слоистость коры, позволяет различать в ней ранний и поздний луб, аналогичный ранней и поздней древесине. Границу годичного кольца составляют уплотненные ситовидные клетки.
В связи с тем, что поздний луб часто редуцируется, границы годичных приростов не всегда хорошо заметны. Годичная слоистость хорошо выражена у сосны обыкновенной, с. черной, с. крымской, с. веймутовой, с. гималайской. Трудно определить возраст коры у сосны пицундской. В лубе сосны приморской слоистость полностью отсутствует. Сердцевинные лучи в проводящей флоэме обычно однорядные и редко содержат смоляные каналы. В непроводящей флоэме сердцевинные лучи сильно извилисты.
Таким образом, кора многолетних хвойных растений состоит из корки и вторичного луба, в котором можно различить проводящую и непроводящую зоны. Непроводящая флоэма сосны может быть разделена на внутреннюю, граничащую с проводящей флоэмой и характеризующуюся типичным для нее строением; внутреннюю, или промежуточную, в которой наблюдается образование смолоносных вместилищ, увеличивается число кристаллических клеток и начинается делатация, и наружную, граничащую с коркой, состоящую из делатационной паренхимы, в которой закладывается феллоген.
Видоспецифическим является характер пробки. У большинства сосен феллодерма состоит из 4 - 5 рядов клеток.
При диагностике хвойных по коре важное значение имеют особенности отложения щавелекислого кальция (оксалата кальция). У сосновых, в отличие от других семейств, кристаллы оксалата кальция крупные, одиночные, которые находятся в полости клеток тяжевой паренхимы. У кедра и пихты они встречаются и в клетках лучевой паренхимы. Форму кристаллов, варьирующую от длинной до призматической, напоминающей стилоиды (ель, пихта, кедр, лиственница), включают в ранг родового признака. Смолоносная система у хвойных свойственна представителям семейств сосновых, араукариевых и кипарисовых.
Смолоносный аппарат у сосновых состоит из радиальных смоляных ходов и смолоносных полостей в виде шаровидных вместилищ, возникающих в сердцевинных лучах. У кипарисовых мешковидные смолоносные вместилища возникают в клетках тяжевой паренхимы.
К смолоносной системе относятся также слизевые (пихта, кетелеерия) и масляные (кипарисовые) клетки, содержащие масла. Кора кипарисовых отличается от таксодиевых только по содержанию смоловместилищ.
Наибольшее значение при определении хвойных пород по коре имеют особенности строения совокупности одревесневших элементов, которые получили название твердого луба. Сравнение ведется по строению смолоносной системы, формы кристаллизации щавелевокислого кальция и особенности расположения тканей на поперечных срезах коры (см. ключ по Лотовой Л.И.).
Твердый луб у хвойных состоит только из волокон, из волокон и склереид или только из склереид. Волокна развиваются непосредственно из материнских клеток луба и появляются уже близ камбиальной зоны. В поперечном сечении они квадратные, прямоугольные или многоугольные, нередко со слегка закругленными углами. Клеточные полости очень малы, очертания их округлые или угловатые.
Склереиды возникают из клеток тяжевой и лучевой паренхимы. Наиболее часто встречаются каменистые клетки (брахисклереиды) и более или менее разветвленные склереиды (астросклереиды). Стенки всех склереид очень толстые, сильно одревесневшие, тонкослойные с многочисленными поровыми каналами. Основным критерием, позволяющим различать волокнистые трахеиды и волокна, служит приуроченность склереид к непроводящему лубу.
10.2. Смолоносная система древесины и коры хвойных растений
Возможности диагностики хвойных по строению смолоносной системы широко используются при установлении родственных связей между семействами, родами и видами.
Смоловыделения представляют собой защитную реакцию растения на поранение. Активный синтез терпеноидных соединений, входящих в состав смолы, играет важную роль в повышении морозоустойчивости растений и усилении их сопротивляемости болезнетворным агентам.
Вместе с дубильными веществами они относятся к фенольным соединениям и одинаково задерживают развитие патогенных организмов.
Различают первичные смоляные каналы, которые имеются в первичной коре молодых стеблей хвойных, кроме тиса. Они возникают схизогенно (без участия камбия), выстланы тонкостенными эпителиальными клетками и окружены обкладкой из мелких паренхимных клеток, часто заполненных крахмалом.
В большинстве случаев эти каналы связаны с листовыми следами, располагаются по 1 - 3 листовых подушечках или более глубоко в первичной коре. Долговечность первичных смоляных каналов зависит от места и времени заложения феллогена. Очень рано отмирают смоляные каналы, находящиеся в листовых подушках лиственницы, так как уже в первом году жизни побега под подушками формируется перидерма.
При более глубоком расположении смоляных каналов они сохраняются в течение нескольких лет и срастаются вместе с чешуями корки после формирования внутренних перидерм. Наиболее долговечны первичные смоляные каналы у араукариевых и пихты, у которых делатированная первичная кора существует десятилетиями. У этих растений смоляные каналы сильно разрастаются в тангенциальном направлении. В первичной коре молодых побегов пихты, кроме смоляных каналов, встречаются вместилища выделений, которые относят к категории идиобластов. По данным Л.И. Лотовой [9], особенно много идиобластов в первичной коре испанской (Abies pinsapo Boiss) и алжирской
Вторичная смоловыделительная система развита только в лубе араукариевых, кипарисовых и сосновых.
В лубе кипарисовых встречаются разные по степени морфологической специализации секреторные образования: идиобласты, полости схизо-лизогенного происхождения и вертикальные каналы, развитие которых осуществляется схизогенным путем. Образованию отдельных полостей или каналов предшествует деление тангенциальными перегородками одиночных или двух-трех рядом расположенных клеток тяжевой паренхимы. Чаще всего деления начинаются одновременно в клетках двух соседних тангенциальных слоев. В обеих группах клеток, возникших вследствие этого деления, схизогенно формируются парные межклетники - полости будущих смоловместилищ, размеры которых увеличиваются за счет частичного лизиса окружающих его тонкостенных клеток. Паренхимные клетки, выстилающие полость вместилища, становятся эпителиальными. Слой волокон, разделяющих пару образовавшихся смоловместилищ, нередко разрывается, и возникает одна крупная полость. Смоловместилища расположены вертикальными рядами. У можжевельника вместилища обычно короткие, у кипариса вытянуты в длину и имеют вид слепых каналов, у туи западной вертикальные каналы достигают нескольких сантиметров.
Идиобласты, характерные для вторичной коры пихты и кетелеерии, обычно называют слизевыми клетками, так как, кроме смолы, они содержат слизь и фенольные соединения. Появление этих идиобластов обусловлено сильным разрастанием отдельных клеток тяжевой, реже лучевой, паренхимы. У большинства родов сосновых образование смоловыделительного аппарата является функцией лучевых инициалей камбия и паренхимных клеток лучей. У сосны, лиственницы, ели, лжетсуги система, секретирующая терпены, появляется уже в проводящей зоне луба в виде горизонтальных каналов, расположенных в многорядных веретеновидных лучах. Эти каналы представляют собой продолжение горизонтальных каналов древесины. Обычно в каждом луче проходит один канал, очень редко их два-три.
В непроводящей зоне луба в лучах могут формироваться замкнутые смоловместилища в виде цист или головчатых образований. Особенно сильное развитие таких смоловместилищ наблюдается у лиственницы сибирской, лиственницы Сукачева, лиственницы даурской и лиственницы Чекановского, отличающихся высокой зимостойкостью.
В настоящее время принято делить вторичную флоэму на проводящую зону, прилегающую к камбию, и непроводящую зону, занимающую периферическое положение.
Проводящая флоэма у большинства древесных растений представляет собой однолетний прирост, так как деятельность ситовидных элементов ограничена, как правило, одним вегетационным периодом.
В непроводящей зоне наиболее важны паренхимные клетки, осуществляющие запасающую функцию. Из-за периферического расположения в стволе дерева луб больше, чем древесина, должен противостоять механическим и биологическим повреждениям. Это достигается приобретением корой твердости и прочности вследствие дифференциации в непроводящей зоне луба механических элементов, а также образования у многих древесных растений секреторной системы, вырабатывающей вещества, обладающие антисептическими свойствами. Высокая митотическая активность паренхимы в наружных слоях непроводящей флоэмы обусловливает развитие в ней дилатационной зоны, представляющей собой не только мощный резервуар запасных веществ, но служащей также местом заложения феллогена, появление которого предшествует формированию корки, которая обеспечивает возможность удаления вредных для растений продуктов метаболизма. Л.И. Лотова [10] обратила внимание на то, что между лубом и древесиной имеются существенные корреляции по строению и расположению древесинной паренхимы и структурными особенностями ситовидных трубок, так как обе ткани развиваются из производных одной меристемы - камбия. Ситовидные клетки и трахеиды имеют большое сходство в очертаниях ситовидных почек и окаймленных пор, расположенных на их радиальных стенках.
Задание 12. Сравнительная характеристика хвойных растений по древесине и коре
Цель задания - изучение особенностей микроскопического и анатомического строения древесины и коры хвойных пород на поперечном, радиальном и тангенциальном срезах при малом увеличении микроскопа.
Материал и оборудование: 1) микроскоп; 2) образцы древесины и коры сосны обыкновенной, ели европейской, пихты сибирской, пихты кавказской, лиственницы сибирской, псевдотсуги Мензиса, можжевельника обыкновенного, кипариса вечнозеленого, туи западной, кипарисовика Лавсона, секвойи вечнозеленой; 3) постоянные препараты поперечных, радиальных и тангенциальных срезов древесины и коры перечисленных выше видов хвойных пород.
-
1. Зарисуйте участок древесины сосны обыкновенной на поперечном срезе в пределах одного годичного кольца.
2. Обозначьте элементы древесины.
3. Зарисуйте и обозначьте элементы древесины на радиальном срезе.
4. Зарисуйте и обозначьте элементы древесины на тангенциальном срезе.
Предварительно рассмотрите образцы древесины невооруженным глазом, обратите внимание, что древесина сосны ядровая. Заболонь желтовато-бурого цвета, а ядро розоватое, красноватое и буровато-красное. Годичные слои хорошо видны на всех разрезах. Ранняя часть годичного слоя светлого оттенка, поздняя часть более темной окраски. Переход от ранней древесины к поздней резкий. Сердцевинные лучи простым глазом не видны ни на одном разрезе. Смоляные ходы сосредоточены главным образом в поздней части слоя и видны на поперечном разрезе в виде светлых точек, а на радиальном и тангенциальном разрезах в виде беловато-матовых точек.
Работа с микропрепаратами производится при малом увеличении микроскопа. На поперечном срезе древесины сосны обыкновенной видны однообразные четырех-шестиугольные клетки, расположенные правильными рядами. Это трахеиды. Просматривая весь препарат, обратите внимание на то, что трахеиды имеют разное строение. Одни трахеиды тонкостенные с крупными полостями и окаймленными порами на радиальных стенках. Это весенние, или ранние, трахеиды. Они образуются весной и в начале лета и выполняют проводящую функцию.
Другая часть древесины состоит из мелких, толстостенных трахеид. Эта древесина называется поздней; она образуется во второй половине лета и выполняет механическую функцию. Слой древесины, состоящий из ранних и поздних трахеид, называется годичным слоем, или годичным кольцом. Переход от ранней древесины к поздней постепенный, а от поздней к ранней древесине следующего года резкий, поэтому границы годичного слоя хорошо заметны.
В поздней древесине встречаются крупные полости - это смоляные ходы. Они окружены мелкими неодревесневшими клетками древесной паренхимы. В виде радиальных полосок видны сердцевинные лучи; у сосны они очень узкие, состоят из одного слоя живых, вытянутых в длину клеток.
На радиальном срезе древесины сосны видны, длинны почти параллельные полоски - стенки трахеид. Передвигая препарат вверх и вниз, можно увидеть, что в некоторых местах трахеиды на концах заострены или закруглены. Эти концами одна трахеида очень плотно вклинивается в другую. Как и на поперечном срезе, хорошо видна слоистость древесины. Весенние трахеиды более широкие, на их стенках имеется множество отверстий в виде двух концентрических окружностей. Эти отверстия называются окаймленными порами, и служат они для передвижения воды из одной трахеиды в другую. Поздние трахеиды плотные, темноокрашенные, поры в них или отсутствуют или очень мелкие. Слой древесины из ранних и поздних трахеид составляет годичный прирост (годичный слой). Поперек трахеид проходят полоски из 3 - 6 рядов клеток. Это сердцевинные лучи. На радиальном срезе видна их высота. Средние ряды сердцевинных лучей состоят из живых клеток с протоплазмой и клеточным ядром и несут запасы питательных веществ, а наружные клетки мертвые. В центре некоторых лучей проходит полый цилиндр - это поперечный смоляной ход. Среди клеток трахеид встречаются и продольные смоляные ходы. На продольном тангенциальном срезе хорошо видны трахеиды с окаймленными порами. Поры видны на боковых стенках трахеид и имеют вид вилочек, обращенных друг к другу. Сердцевинные лучи, перерезанные поперек, имеют вид веретен. Некоторые лучи сильно расширены в центре из-за наличия в них смоляных ходов. Годичной слоистости на данном срезе не видно.
Строение коры хвойных пород. Рассмотрите особенности микро-и макроскопического строения коры побегов, молодых и старых стволов сосны обыкновенной на образцах коры и препаратах (поперечные, радиальные, тангенциальные срезы).
Обратите внимание на строение проводящего, непроводящего луба и корки. Сравните кору и корку сосны обыкновенной с елью европейской, лиственницей сибирской, пихтой сибирской и псевдотсугой Мензиса.
Обратите внимание на характер смолоносной системы в коре перечисленных видов семейства Сосновые.
В порядке самоконтроля, пользуясь собственными наблюдениями, а также определителями данного "Практикума", составьте ключи по определению пяти родов сосновых по древесине, коре и корке.
Древесина — слоисто-пористый материал растительного происхождения. Состоит из многообразных клеток, связанных между собой порами. Строение древесины сложно и совершенно. Природа создала материал с такими уникальными качествами как: твердость, низкая тепло-, звукопроницаемость, высокая прочность. Дерево с легкостью поддается обработке инструментами, хорошо склеивается. Древесина была и остается ведущим материалом для изготовления домов.

Макроскопическое строение древесины
Если на срезе заметно строение штамба невооруженным взглядом, то говорят о таком понятии, как макростроение древесины. Бывает, что не вся плоскость среза окрашена равномерно: ближе к центру она может быть темнее, а дальше — светлее. Темная часть, самая прочная, созданная из мертвых клеток ткани, является ядром, а светлая — заболонью. Клетки ядра отмирают из-за закупорки проводящих сосудов смолой. Породы древесины с такой окраской называют ядровыми (дуб, сосна, ясень, лиственница). Если срез окрашен равномерно, то такие породы являются безъядровыми (ольха, береза).

Каждый год жизни отмечается на стволе увеличением слоя древесины определенного размера, который зависит от возраста, условий жизни растения, скорости роста. Такие слои называют годичными кольцами. Они особенно ярко видны на спилах хвойных пород.
Годичное кольцо содержит два таких слоя ткани, как:
- Ранний мягкий. Возникает в первой половине годичной вегетации. Имеет светлую окраску. Находится у центра ствола.
- Поздний твердый. Создается во второй половине годичного роста. Отличается темной окраской. Располагается ближе к коре.
По каналам ранней ткани транспортируются питательные элементы к вершине и обратно. Зона позднего слоя защищает растение от механических повреждений. В комле находятся самые узкие кольца. Из-за плохих условий произрастания они могут иметь волнистость, что повышает декоративность древесной массы. Древесный материал с самыми узкими кольцами считается лучшим.
От коры по радиусу к центру растения протягиваются светлые линии, используемые для перемещения питательных элементов. Эти линии называются сердцевинными лучами. Лучи характерны для всех пород. Они отчетливо видны на срезах. Ширина лучей меняется в пределах 0,05 — 1 мм. Их размер непосредственно зависит от условий жизни растения. Сердцевинные лучи ответственны за текстуру древесной массы. Некоторые из них прерываются далеко от сердцевины. Такие лучи называются вторичными в отличие от первичных, которые доходят до нее.

На поперечном разрезе лиственных пород замечаются небольшие отверстия, которые являются сосудами растения. Они поставляют дереву воду и питание. Если в раннем слое лежат крупные сосуды, а в позднем — мелкие, то такой сосудистый рисунок соответствует кольцесосудистой материи (дуб, вяз, ясень). Она отличается прочной древесной массой. Равномерное расположение сосудов по годичному кольцу соответствует мягкой рассеяннососудистой ткани (береза, осина). Весной с некоторых деревьев (береза, сахарный клен) собирают сок, перерезая их сосуды.
Для строения хвойных пород характерны протоки, наполненных смолой. Это смоляные ходы, которые свойственны лишь некоторым хвойным деревьям. Например, пихта и можжевельник их не имеют. Смоляные ходы разных направлений создают одну смолоносную систему.

Самый центр ствола занимает сердцевина, рыхлая масса, пронизывающая растение снизу доверху. Быстро поддается разложению. Она создается в начале жизни дерева. На срезе сердцевина представляется в виде отметины, как правило, круглой формы диаметром 2 — 5 мм. Лиственные породы имеют сердцевину большей площади, чем хвойные. Наибольшую сердцевину имеет бузина.
Тонкий слизистый слой клеток, лежащих в области между корой и древесиной, именуется камбием. Он вырабатывает микроэлементы для роста древесной ткани, принимая от луба питательные элементы. Начинаясь весной, процесс синтезирования новых клеток заканчивается осенью. Этим объясняется слоистое строение древесной массы.
Микроскопическое строение древесины
Только микроскопическое строение древесины отвечает в полной мере на вопрос: что же такое древесина? Множество разнообразных клеток, скрепленных между собой — это есть древесная масса. Каждая клетка наполнена протопластом, а межклеточное пространство — сложными полимерными соединениями. Однозначные по строению и функциям клетки создают соответствующие ткани: механические (опорные), проводящие и запасающие.

Оболочка клетки создана из природных высокомолекулярных полимеров: углеводов (70 — 80%) и лигнина (20 — 30%). Углеводная часть представлена холоцеллюлозой, гемицеллюлозой и целлюлозой. Лигнин — аморфное вещество, связывающее целлюлозные волокна между собой, благодаря чему целлюлоза приобретает прочность и эластичность. Лигнин и целлюлоза пропитывают стенки клеток, вызывая их одревеснение. В результате оболочка становится жесткой, твердой, по своей прочности не уступающая железобетону.
Химический состав древесины и коры
Ткань древесной массы создана из клеток. Поэтому все химические компоненты располагаются в клеточных оболочках. Древесина состоит из минеральных и органических компонентов. К минеральным (неорганическим) веществам относятся элементы, которые остаются после сгорания древесной ткани (зола). Их величина составляет 1% от общей массы. По химическому составу эти элементы представляют собой смесь разных солей, растворимых (натрия, калия) и нерастворимых (магния, кальция, железа) в воде.
Остальную часть занимают органические составляющие, занимающие 99% общей массы. Их элементный состав содержит 49 — 50% углерода, 43 — 44 % кислорода, 6 % водорода и 0,1 — 0,3 % азота.

Органические вещества представлены в виде двух групп:
- Структурные компоненты, образующие структуру клетки (целлюлоза, холоцеллюлоза, гемицеллюлоза, лигнин).
- Экстрактивные вещества — компоненты, которые можно извлечь из древесины растворителями (экстрагировать). Они не входят в состав клеточной стенки. К ним относятся эфирные масла, красители, дубильные вещества, жиры, пектины. Древесина обязана им запахом, цветом, вкусом, сопротивлению гниению и болезням. Экстрактивные элементы составляют 3 — 5% от общей массы органических компонентов.
Химический состав лиственных пород отличается от хвойных большим содержанием структурных компонентов (гемицеллюлозы), но меньшим содержанием лигнина. В зависимости от географического места произрастания, возраста растения химический состав может меняться в пределах одной породы.
Химический состав коры отличается повышенным содержанием экстрактивных веществ, лигнина и пониженным содержанием целлюлозы. Доля неорганических веществ в общем количестве составляет 10-15 %, это в 10 раз больше, чем в древесине. Преобладающими элементами золы являются кальций (82-95 %), калий, магний.

Кора — ценное растительное сырье:
- Дубильные вещества незаменимы при выделке кож.
- Экстрактивные компоненты находят применение в медицине.
- Кору используют в качестве топлива.
- Измельченная кора служит основой корокомпостов в сельском хозяйстве.
Разные химические составы коры и древесины приводят к необходимости перерабатывать их раздельно.
Пороки древесины
Повреждения всей структуры древесины или отдельных участков, которые снижают качество и ограничивают применение, называются пороками древесины. Некоторые пороки возникают в растущем дереве, другие — при хранении или эксплуатации сырья. Качество древесной массы определяется в соответствии с видами и размерами пороков, их расположения, назначения продукции.

ГОСТ 2140-81 устанавливает классификацию пороков по следующим группам:
- Сучки, основания бывших ветвей.
- Трещины, разрывы ткани вдоль волокон.
- Дефекты формы ствола. К ним относятся сужение ствола, кривизна, овальность, наросты, закомелистость.
- Пороки строения древесины: прожилки, полоски, пятнышки.
- Химические окраски. Они возникают в заготовленном сырье в результате окисления дубильных веществ.
- Грибные повреждения. Их вызывают грибы, которые являются растительными организмами, развивающимися из спор.
- Повреждения насекомыми, птицами. Такой порок ухудшает декоративность и физико-механические качества.
- Инородные включения.
- Покоробленность. Порок возникает в результате обработки материала.
ГОСТ 2140-81 Видимые пороки древесины. Классификация, термины и определения, способы измерения
Основные части дерева
Дерево — многолетнее растение, состоящее из таких компонентов как:
Каждый элемент выполняет конкретную задачу и имеет свое предназначение для хозяйственных нужд. Строение деревьев лиственных пород не отличается от строения хвойных.
Ствол – это доля дерева, расположенная выше корней. В густом лесу только с его помощью ветви могут достичь света. По штамбу вверх и вниз перемещаются элементы питания и вода. Сверху ствол заканчивается тонкой вершиной. Нижняя зона, расположенная сразу над корнем, называется комлем. Ствол — основной материал для строительства. Он используется как источник тепла для нужд потребителей, служит сырьем для производства композитных материалов (ДСП, МДФ), скипидара, канифоли.
Снаружи штамб покрыт оболочкой: корой, которая является кожухом, защищающим дерево. Строение и свойства коры имеют свои особенности. Она включает в себя два слоя: наружный корковый или пробковый, состоящий из мертвых клеток луба, и внутренний лубяной, являющийся главной артерией ствола. Корковый слой оберегает древесную массу от повреждений.

Использование кроны для хозяйственных нужд невелико, несмотря на ценность материала. Измельченные листья или хвоя используются как витаминная добавка для корма скота или птицы. После сгорания веток получается зола, являющаяся ценным удобрением. Из раздробленных веток садоводы изготавливают мульчу для защиты корней растений от мороза.
Строение дерева таково, что важны все элементы. Но корни — главная составляющая. Если по каким-то причинам гибнут крона и штамб, то дерево может возродиться благодаря корням. Корневая система отличается сложным строением. Главный корень разветвляется на мелкие втягивающие корешки. Они собирают питательные элементы с большой площади и поставляют их всем зонам. Обширная корневая система поддерживает растение вертикально. Корни не служат топливом, не используются для хозяйственных нужд.
Читайте также: