
Рецепторная функция плазмалеммы краткое содержание
Обновлено: 02.07.2024
Механическая устойчивость плазматической мембраны определяется не только свойствами самой мембраны, но и свойствами прилежащих к ней гликокаликса и кортикального слоя цитоплазмы.
Внешняя поверхность плазматической мембраны покрыта рыхлым волокнистым слоем вещества толщиной 3-4 нм — гликокаликсом. Он состоит из ветвящихся полисахаридных цепей мембранных интегральных белков, между которыми могут располагаться выделенные клеткой гликолипиды и протеогликаны. Тут же обнаруживаются некоторые клеточные гидролитические ферменты, участвующие во внеклеточном расщеплении веществ (внеклеточное пищеварение, например, в эпителии кишечника). Кортикальный слой цитоплазмы, толщиной 0,1-0,5 мкм, не содержит рибосом и мембранных структур, но богат актиновыми микрофиламентами.
Плазматическая мембрана, как и другие липопротеидные мембраны клетки, является полупроницаемой. Максимальной проникающей способностью обладает вода и растворенные в ней газы. Транспорт ионов может проходить по градиенту концентраций, т. е. пассивно, без затрат энергии. В этом случае некоторые мембранные транспортные белки образуют молекулярные комплексы, каналы, через которые ионы проходят сквозь мембрану за счет простой диффузии. В других случаях специальные мембранные белки-переносчики избирательно связываются с тем или иным ионом и переносят его через мембрану. Такой тип переноса называется активным транспортом и осуществляется с помощью белковых ионных насосов. Например, затрачивая 1 молекулу АТФ, система К-Nа насоса откачивает за один цикл из клетки 3 иона Nа и закачивает 2 иона К против градиента концентрации. В сочетании с активным транспортом ионов через плазмалемму проникают различные сахара, нуклеотиды и аминокислоты. Макромолекулы, такие как, например, белки, через мембрану не проходят. Они, а также более крупные частицы вещества транспортируются внутрь клетки посредством эндоцитоза. При эндоцитозе определенный участок плазмалеммы захватывает, обволакивает внеклеточный материал, заключает его в мембранную вакуоль. Эта вакуоль — эндосома — сливается в цитоплазме с первичной лизосомой и происходит переваривание захваченного материала. Эндоцитоз формально разделяют на фагоцитоз (поглощение клеткой крупных частиц) и пиноцитоз (поглощение растворов). Плазматическая мембрана принимает участие и в выведении веществ из клетки с помощью экзоцитоза — процесса, обратного эндоцитозу.
Рецептурная роль плазмолеммы. Белки-переносчики внешней мембраны клетки являются также рецепторами, узнающими определенные ионы и взаимодействующими с ними. В качестве рецепторов на поверхности клетки могут выступать белки мембраны или элементы гликокаликса. Такие чувствительные к отдельным веществам участки разбросаны по поверхности клетки или собраны в небольшие зоны. Роль многих клеточных рецепторов заключается не только в связывании специфических веществ, но и в передаче сигналов с поверхности внутрь клетки. Например, при действии гормона на клетку цепь событий развертывается следующим образом: молекула гормона специфически взаимодействует с рецепторным белком плазмалеммы и, не проникая в клетку, активирует фермент, синтезирующий ЦАМФ. Последний активирует или ингибирует внутриклеточный фермент или группу ферментов.
Обязательным компонентом поверхностного аппарата любой клетки является цитоплазматическая мембрана (плазмалемма). Она отделяет и защищает внутреннее содержимое клетки от внешней среды. Важнейшим свойством цитоплазматической мембраны является избирательная проницаемость. Различные вещества проходят через плазмалемму с разной скоростью, а для некоторых она практически непроницаема. Таким образом, цитоплазматическая мембрана обеспечивает обмен веществ между клеткой и внеклеточной средой и постоянство химического состава клетки.
Химический состав и строение плазмалеммы. Цитоплазматическая мембрана состоит преимущественно из липидов и белков. В состав плазмалеммы также входят углеводы, которые являются компонентами сложных липидов и белков — гликолипидов и гликопротеинов. Толщина цитоплазматической мембраны составляет около 7,5 нм.
Около половины массы плазмалеммы составляют липиды, представленные в основном фосфолипидами. Они формируют основу строения мембраны — липидный бислой (двойной слой), в котором гидрофобные хвосты молекул погружены внутрь, а гидрофильные головки располагаются снаружи (см. рис. 6.3). Кроме фосфолипидов, в состав двойного слоя входят и другие липиды например стероиды.
*С липидным бислоем связаны белки. Молекулы интегральных белков пронизывают двойной слой липидов насквозь. Полуинтегральными называют белки, погруженные лишь в один из двух липидных слоев (внешний либо внутренний). Периферические белки примыкают к поверхности бислоя с наружной или внутренней стороны (рис. 11.1).*
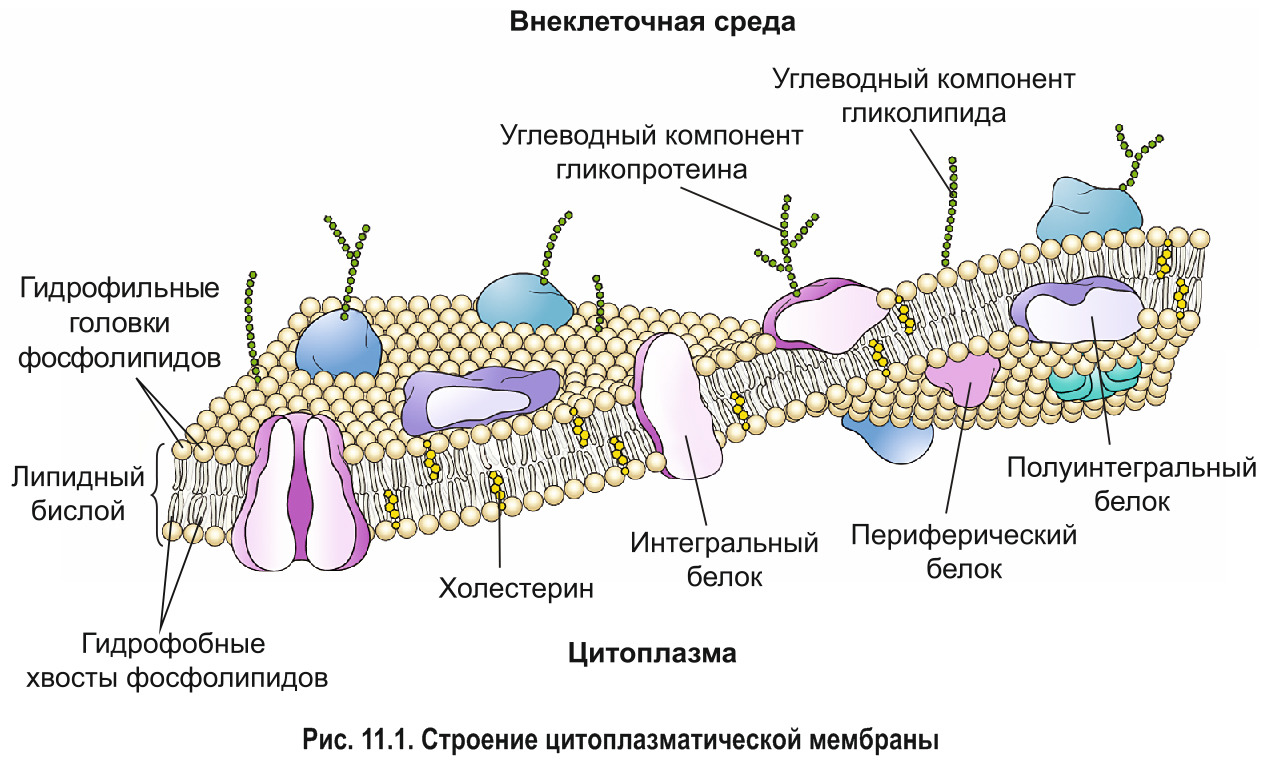
Мембранные липиды и белки связаны между собой не ковалентными связями, а за счет гидрофобных и электростатических взаимодействий. В связи с этим молекулы данных веществ могут двигаться вдоль плоскости мембраны. Таким образом, плазмалемма является динамичной структурой — молекулы белков перемещаются в подвижном, текучем липидном бислое, как в жидкости. *Данная модель организации мембраны получила название жидкостно-мозаичной. Она была предложена американскими биологами Дж. Сингером и Г. Николсоном в 1972 г.* Свойство текучести обусловливает пластичность плазмалеммы, благодаря чему она способна, например, быстро восстанавливать свою целостность после незначительных повреждений.
*Движение в плоскости плазмалеммы происходит достаточно легко. Однако переход белков с одной стороны мембраны на другую (так называемый флип-флоп переход) практически невозможен. Подобное перемещение липидов происходит, но очень редко, при участии специальных белков флиппаз. Поэтому состав наружного и внутреннего слоев плазмалеммы различается.*
*Текучесть мембраны зависит от ее липидного состава и температуры. С увеличением содержания остатков ненасыщенных жирных кислот или температуры подвижность молекул возрастает. Стероиды , например холестерин в организме животных, регулируют текучесть плазмалеммы и придают ей жесткость. Стероиды располагаются в свободном пространстве между гидрофобными хвостами фосфолипидов, ограничивая подвижность их молекул друг относительно друга (см. рис. 11.1). Таким образом, стероиды обеспечивают стабильность мембраны и поддержание ее структуры. Кроме того, они регулируют проницаемость плазмалеммы.*
В клетках эукариот, кроме плазмалеммы, имеются внутренние мембраны, ограничивающие ядро и мембранные органоиды. Для всех биологических мембран характерен общий план строения — их основу составляет липидный бислой, с которым связаны молекулы белков.
Плазмолемма (plasmalemma), или внешняя клеточная мембрана, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку.
Химический состав плазмолеммы.
Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран.
Снаружи от плазмолеммы располагается надмембранный слой — гликокаликс (glycocalyx). Толщина этого слоя около 3-4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гликопротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы (см. рис. 5). При использовании специальных методов выявления полисахаридов (краситель рутениевый красный) видно, что они образуют как бы чехол поверх плазматической мамбраны.
В гликокаликсе могут располагаться белки, не связанные непосредственно с билипидным слоем. Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др.
Функции плазмолеммы. Эта мембрана выполняет ряд важнейших клеточных функций, ведущими из которых являются барьерная функция (разграничения цитоплазмы с внешней средой), функции рецепции и транспорта различных веществ как внутрь клетки, так и из нее.
С плазмолеммой связана локализация специфических рецепторов, отвечающих за такие важные процессы, как взаимное распознавание клеток, развитие иммунитета, рецепторов, реагирующих на физические факторы. Так, в плазмолемме светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что в свою очередь приводит к генерации электрического импульса.
Плазмолемма (plasmalemma), или внешняя клеточная мембрана, среди различных клеточных мембран занимает особое место. Это поверхностная периферическая структура, не только ограничивающая клетку снаружи, но и обеспечивающая ее непосредственную связь с внеклеточной средой, а следовательно, и со всеми веществами и стимулами, воздействующими на клетку.
Химический состав плазмолеммы.
Основу плазмолеммы составляет липопротеиновый комплекс. Она имеет толщину около 10 нм и, таким образом, является самой толстой из клеточных мембран.
Снаружи от плазмолеммы располагается надмембранный слой — гликокаликс (glycocalyx). Толщина этого слоя около 3-4 нм, он обнаружен практически у всех животных клеток, но степень его выраженности различна. Гликокаликс представляет собой ассоциированный с плазмолеммой гликопротеиновый комплекс, в состав которого входят различные углеводы. Углеводы образуют длинные, ветвящиеся цепочки полисахаридов, связанные с белками и липидами, входящими в состав плазмолеммы (см. рис. 5). При использовании специальных методов выявления полисахаридов (краситель рутениевый красный) видно, что они образуют как бы чехол поверх плазматической мамбраны.
В гликокаликсе могут располагаться белки, не связанные непосредственно с билипидным слоем. Как правило, это белки-ферменты, участвующие во внеклеточном расщеплении различных веществ, таких как углеводы, белки, жиры и др.

Функции плазмолеммы. Эта мембрана выполняет ряд важнейших клеточных функций, ведущими из которых являются барьерная функция (разграничения цитоплазмы с внешней средой), функции рецепции и транспорта различных веществ как внутрь клетки, так и из нее.
С плазмолеммой связана локализация специфических рецепторов, отвечающих за такие важные процессы, как взаимное распознавание клеток, развитие иммунитета, рецепторов, реагирующих на физические факторы. Так, в плазмолемме светочувствительных клеток животных расположена специальная система фоторецепторных белков (родопсин), с помощью которых световой сигнал превращается в химический, что в свою очередь приводит к генерации электрического импульса.
• Различные молекулы проходят через цитоплазматическую мембрану за счет пассивной диффузии или активного перемещения
• Большинство растворенных веществ перемещается через мембрану с участием специфических транспортных белков
• Цитоплазматическая мембрана поддерживает протонный градиент между цитоплазмой и внеклеточной средой
Клетки всех организмов обладают цитоплазматической (или плазматической) мембраной, которая предотвращает выход и поступление в клетку растворимых соединений. У прокариот цитоплазматическая мембрана, толщиной примерно 8 нм, служит барьером между содержимым клетки и окружающей средой. Цитоплазматические мембраны содержат липиды и белки. Так же как и для других биологических мембран, в основе их структуры лежит фосфолипидный бислой. Фосфолипиды содержат фосфатную группу, присоединенную к основной трехуглеродной структуре глицерина.
Гидрофобные цепи жирных кислот связаны со свободными атомами углерода этой структуры, причем они ориентированы по направлению друг к другу и в противоположную сторону от внешней среды, содержащей воду. Напротив, гидрофильные фосфатные группы ориентированы в сторону водного окружения. Цитоплазматическая мембрана не дает возможность большинству биологических молекул и ионов пассивно диффундировать через нее в обоих направлениях. Исключение составляет вода, которая способна к медленной диффузии через цитоплазматическую мембрану; небольшой размер молекул и отсутствие заряда позволяют ей свободно проходить через фосфолипидный бислой.
С цитоплазматической мембраной связано много типов белков. Часто мембранные белки содержат участки гидрофобных аминокислот, которые взаимодействуют с гидрофобными цепями жирных кислот, находящихся в мембране. Белки, гидрофобные участки которых пронизывают мембрану, называются интегральными мембранными белками. Многие белки, находящиеся в цитоплазматической мембране, играют определенную роль в транспорте молекул в клетку и в выходе из нее. Такой транспорт через плазматическую мембрану может представлять собой активный или пассивный процесс. При пассивном транспорте молекулы движутся по концентрационному градиенту, т. е. из области их высокой концентрации в область с низкой концентрацией.
Таким образом, пассивный транспорт не требует затрат энергии. В противоположность пассивной диффузии, в результате активного транспорта, внутри клетки и вне ее устанавливаются различные концентрации растворенных метаболитов. Транспортные системы состоят из белков, которые связаны или с мембраной в виде ее стабильных интегральных компонентов, или с расположенными на периферии липидами через модифицированные концевые аминогруппы, за счет которых белок прикрепляется к поверхности мембраны. Такие большие молекулы, как белки, которые не могут свободно диффундировать через мембраны, транспортируются в процессе активного транспорта. Часто транспортные системы проявляют высокую специфичность, транспортируя только молекулы одного вещества или группы веществ.
Комплекс транспортных белков, связывающий АТФ (ABC), представляет собой самое обширное семейство транспортных белков прокариот. Только у Е. coli оно насчитывает свыше 200 белков. ABC-переносчики могут перемещать субстраты в клетку и из нее. Они проявляют активность по отношению к разнообразным субстратам, от ионов до молекул белков. ABC-переносчики, транспортирующие субстраты в клетки грамотрицательных бактерий, обычно состоят из трех компонентов: транспортный белок, пронизывающий мембрану, периплазматический белок, связывающий субстрат, и белок, гидролизующий АТФ, локализованный в цитоплазме. Периплазматические связывающие белки проявляют крайне высокое сродство к специфическим субстратам.
Это позволяет им осуществлять транспорт субстратов, даже если они присутствуют в крайне низких концентрациях. Белки связывающие АТФ, расположенные на цитоплазматической стороне мембраны, обеспечивают энергией процесс транспорта. Эти белки стабильно связаны с мембраной через липидную часть со стороны N-концевого участка.
Цитоплазматическая мембрана играет важную роль в энергетических процессах у прокариот. Электроны, которые генерируются в процессе дыхания, сопрягаются со своими рецепторами в мембране. Протоны транспортируются к клеточной поверхности посредством мембранных транспортных белков; при этом наружная мембрана приобретает слабый положительный заряд.
Внутренняя часть мембраны заряжена отрицательно. Таким образом, создается трансмембранный протонный градиент. Энергетически выгодное движение протонов по градиенту, от наружной поверхности вглубь клетки, обеспечивает протекание многих внутриклеточных реакций. Иными словами, энергия, запасенная в цитоплазматической мембране может использоваться клеткой для разных целей. Например, ферменты используют протонный градиент для образования АТФ из АДФ. Некоторые мембраносвязанные ферментные комплексы способствуют генерации протонного градиента в ходе окислительного фосфорилирования.
При окислительном фосфорилировании терминальным акцептором электронов служит кислород. Однако в анаэробных условиях многие прокариоты могут использовать другие акцепторы электронов, например серу, азот, железо и марганец. Энергия, которая запасается в цитоплазматической мембране за счет функционирования ферментов, обеспечивает большую часть потребностей растущей клетки, т. е. такие процессы, как синтез молекул, транспорт белков и субстратов, а также ее подвижность. Изучение молекулярных механизмов генерации энергии в бактериальных клетках представляет собой активно развивающуюся область современных исследований.
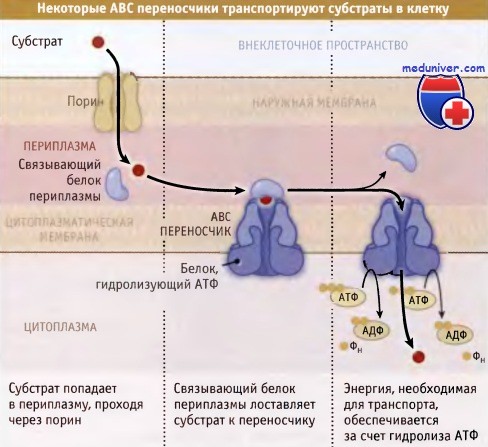
У грамотрицательных бакерий растворимые субстраты поступают в периплазму при участии поринов,
представляющих собой белки, которые образуют в наружной мембране определенных размеров поры.
Субстраты связываются специфическими белками, присутствующими в периплазме, которые, в свою очередь,
в цитоплазматической мембране связываются с ABC переносчиками.
Для транспортировки субстратов в цитоплазму, эти переносчики используют энергию гидролиза АТФ.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: