
Развитие рецепторов и синапсов кратко
Обновлено: 08.07.2024
Синапс - это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую. Термин "синапс" введен Ч. Шеррингтоном и означает "сведение", "соединение", "застежка".
Классификация синапсов. Синапсы можно классифицировать по:
1) их местоположению и принадлежности соответствующим структурам:
• периферические (нервно-мышечные, нейро-секреторные, рецеп-торнонейрональные);
• центральные (аксо-соматические, аксо-дендритные, аксо-аксо-нальные, сомато-девдритные, сомато-соматические);
2) знаку их действия - возбуждающие и тормозящие;
3) способу передачи сигналов - химические, электрические, смешанные.
4) медиатору, с помощью которого осуществляется передача - холинергические, адренергические, серотонинергические, глицинергические и т. д.
Строение синапса. Все синапсы имеют много общего, поэтому строение синапса и механизм передачи возбуждения в нем можно рассмотреть на примере нервно-мышечного синапса (рис. 7).
Синапс состоит из трех основных элементов:
• пресинаптической мембраны (в нервно-мышечном синапсе - это утолщенная концевая пластинка);
Пресинаптическая мембрана - это часть мембраны нервного окончания в области контакта его с мышечным волокном. Постси-наптическая мембрана - часть мембраны мышечного волокна. Часть постсинаптической мембраны, которая расположена напротив пресинаптической, называется субсинаптической мембраной. Осо-бенностью субсинаптической мембраны является наличие в ней специальных рецепторов, чувствительных к определенному медиатору, и наличие хемозависимых каналов. В постсинаптической мем-брне, за пределами субсинаптической, имеются потенциалозависимые каналы.
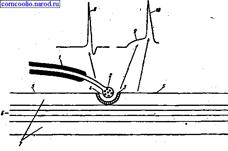
Рис. 7. Строение синапса (схема). 1 - миелинизированное нервное волокно; 2 - нервное окончание с пузырьками медиатора; 3 - субсинаптическая мембрана мышечного волокна; 4 - синаптическая щель; 5 - постсинаптическая мембрана мышечного волокна; 6 - миофибриллы; 7 - саркоплазма; 8 - потенциал действия нервного волокна; 9 - потенциал концевой пластинки (ВПСП); 10 - потенциал действия мышечного волокна.
Механизм передачи возбуждения в химических возбуждающих синапсах. В синапсах с химической передачей возбуждение передатся с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах. Медиаторы в зависимости от их природы делятся на несколько групп:
• моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.);
• Аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.);
• нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.). Медиатор в молекулярном виде находится в пузырьках пресинаптического утолщения (синаптической бляшке), куда он поступает:
• из околоядерной области нейрона с помощью быстрого аксо-нального транспорта (аксотока);
• за счет синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления;
• за счет обратного захвата медиатора из синаптической щели в неизменном виде.
Когда по аксону к его терминалям приходит возбуждение, пресинаптическая мембрана деполяризуется, что сопровождается поступлением ионов кальция из внеклеточной жидкости внутрь нервного окончания. Поступившие ионы кальция активируют перемещение синаптических пузырьков к пресинаптической мембране, их соприкосновение и разрушение (лизис) их мембран с выходом медиатора в синаптическую щель. В ней медиатор диффундирует к суб-синаптической мембране, на которой находятся его рецепторы. Взаимодействие медиатора с рецепторами приводит к открытию преимущественно каналов для ионов натрия. Это приводит к деполяризации субсинаптической мембраны и возникновению так называемого возбуждающего постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе ВПСП называется потенциалом концевой пластинки (ПКП). Между деполяризованной субсинаптической мембраной и соседними с ней участками постсинаптической мембраны возникают местные токи, которые деполяризуют мембрану. Когда они деполяризуют мембрану до критического уровня, в постсинаптической мембране мышечного волокна возникает потенциал действия, который распространяется по мембранам мышечного волокна и вызывает его сокращение.

Химические тормозные синапсы. Эти синапсы по механизму передачи возбуждения сходны с синапсами возбуждающего действия. тормозных синапсах медиатор (например, глицин) взаимодействует с рецепторами субсинаптической мембраны и открывает в ней хлорные каналы, это приводит к движению ионов хлора по концентрационному градиенту внутрь клетки и развитию гиперполяризации на субсинаптической: мембране. Возникает так называемый тормозной постсинаптический потенциал (ТПСП).
Ранее полагали, что каждому медиатору соответствует специфическая реакция постсинаптической клетки - возбуждение или торможение в той или иной форме. В настоящее время установлено, что одному медиатору чаще всего соответствует не один, а несколько различных рецепторов. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц действует на Н-холинорецепторы (чувствительные к никотину), которые открывают широкие каналы для натрия (и калия), что порождает ВПСП (ПКП) В ваго-сердечных синапсах тот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие селективные каналы для ионов калия, поэтому здесь генерируется тормозной постсинаптический потенциал (ТПСП). Следовательно, возбуждающий или тормозной характер действия медиатора определяется свойствами субсинаптической мембраны (точнее, видом рецептора), а не самого медиатора.
Физиологические свойства химических синапсов. Синапсы с химической передачей возбуждения обладают рядом общих свойств:
• Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами субсинаптической мембраны;
• передача возбуждения через синапсы осуществляется медленнее, чем по нервному волокну - синаптическая задержка;
• передача возбуждения осуществляется с помощью специальных химических посредников - медиаторов;
• в синапсах происходит трансформация ритма возбуждения;
• синапсы обладают низкой лабильностью;
• синапсы обладают высокой утомляемостью;
• синапсы обладают высокой чувствительностью к химическим (в том числе и к фармакологическим) веществам.
Электрические синапсы возбуждающего действия. Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока.
Схема передачи возбуждения в электрическом синапсе: ток, вызванный пресинаптическим потенциалом действия, раздражает по-стсинаптическую мембрану, где возникает ВПСП и потенциал действия.
Поперечные каналы объединяют клетки не только электрически, но и химически, так как они проходимы для многих низкомолекулярных соединений. Поэтому возбуждающие электрические синапсы с поперечными каналами формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы).
Общими свойствами возбуждающих электрических синапсов являются:
• быстродействие (значительно превосходит таковое в химических синапсах);
• слабость следовых эффектов при передаче возбуждения (в результате этого в них практически невозможна суммация последовательных сигналов);
• высокая надежность передачи возбуждения.
Возбуждающие электрические синапсы могут возникать при благоприятных условиях и исчезать при неблагоприятных. Например, при повреждении одной из контактирующих клеток ее электрические синапсы с другими клетками ликвидируются. Это свойство называется пластичностью.
Электрические синапсы могут быть с односторонней и двусторонней передачей возбуждения.
Электрический тормозный синапс. Наряду с электрическими синапсами возбуждающего действия могут встречаться электрические тормозные синапсы. Примером такого синапса может служить синапс, который образует нервное окончание на выходном сегменте маутнеровского нейрона у рыб. Тормозящее влияние возникает за счет действия тока, вызванного потенциалом действия пресинапти-ческой мембраны. Пресинаптический потенциал вызывает значительную гиперполяризацию сегмента и гиперполяризующий ток мгновенно тормозит генерацию потенциала действия в начальном сегменте аксона.
В смешанных синапсах Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Синапс - это специализированная структура, которая обеспечивает передачу возбуждения с одной возбудимой структуры на другую. Термин "синапс" введен Ч. Шеррингтоном и означает "сведение", "соединение", "застежка".
Классификация синапсов. Синапсы можно классифицировать по:
1) их местоположению и принадлежности соответствующим структурам:
• периферические (нервно-мышечные, нейро-секреторные, рецеп-торнонейрональные);
• центральные (аксо-соматические, аксо-дендритные, аксо-аксо-нальные, сомато-девдритные, сомато-соматические);
2) знаку их действия - возбуждающие и тормозящие;
3) способу передачи сигналов - химические, электрические, смешанные.
4) медиатору, с помощью которого осуществляется передача - холинергические, адренергические, серотонинергические, глицинергические и т. д.
Строение синапса. Все синапсы имеют много общего, поэтому строение синапса и механизм передачи возбуждения в нем можно рассмотреть на примере нервно-мышечного синапса (рис. 7).
Синапс состоит из трех основных элементов:
• пресинаптической мембраны (в нервно-мышечном синапсе - это утолщенная концевая пластинка);
Пресинаптическая мембрана - это часть мембраны нервного окончания в области контакта его с мышечным волокном. Постси-наптическая мембрана - часть мембраны мышечного волокна. Часть постсинаптической мембраны, которая расположена напротив пресинаптической, называется субсинаптической мембраной. Осо-бенностью субсинаптической мембраны является наличие в ней специальных рецепторов, чувствительных к определенному медиатору, и наличие хемозависимых каналов. В постсинаптической мем-брне, за пределами субсинаптической, имеются потенциалозависимые каналы.
Рис. 7. Строение синапса (схема). 1 - миелинизированное нервное волокно; 2 - нервное окончание с пузырьками медиатора; 3 - субсинаптическая мембрана мышечного волокна; 4 - синаптическая щель; 5 - постсинаптическая мембрана мышечного волокна; 6 - миофибриллы; 7 - саркоплазма; 8 - потенциал действия нервного волокна; 9 - потенциал концевой пластинки (ВПСП); 10 - потенциал действия мышечного волокна.
Механизм передачи возбуждения в химических возбуждающих синапсах. В синапсах с химической передачей возбуждение передатся с помощью медиаторов (посредников). Медиаторы - это химические вещества, которые обеспечивают передачу возбуждения в синапсах. Медиаторы в зависимости от их природы делятся на несколько групп:
• моноамины (ацетилхолин, дофамин, норадреналин, серотонин и др.);
• Аминокислоты (гамма-аминомасляная кислота - ГАМК, глутаминовая кислота, глицин и др.);
• нейропептиды (вещество Р, эндорфины, нейротензин, АКТГ, ангиотензин, вазопрессин, соматостатин и др.). Медиатор в молекулярном виде находится в пузырьках пресинаптического утолщения (синаптической бляшке), куда он поступает:
• из околоядерной области нейрона с помощью быстрого аксо-нального транспорта (аксотока);
• за счет синтеза медиатора, протекающего в синаптических терминалях из продуктов его расщепления;
• за счет обратного захвата медиатора из синаптической щели в неизменном виде.
Когда по аксону к его терминалям приходит возбуждение, пресинаптическая мембрана деполяризуется, что сопровождается поступлением ионов кальция из внеклеточной жидкости внутрь нервного окончания. Поступившие ионы кальция активируют перемещение синаптических пузырьков к пресинаптической мембране, их соприкосновение и разрушение (лизис) их мембран с выходом медиатора в синаптическую щель. В ней медиатор диффундирует к суб-синаптической мембране, на которой находятся его рецепторы. Взаимодействие медиатора с рецепторами приводит к открытию преимущественно каналов для ионов натрия. Это приводит к деполяризации субсинаптической мембраны и возникновению так называемого возбуждающего постсинаптического потенциала (ВПСП). В нервно-мышечном синапсе ВПСП называется потенциалом концевой пластинки (ПКП). Между деполяризованной субсинаптической мембраной и соседними с ней участками постсинаптической мембраны возникают местные токи, которые деполяризуют мембрану. Когда они деполяризуют мембрану до критического уровня, в постсинаптической мембране мышечного волокна возникает потенциал действия, который распространяется по мембранам мышечного волокна и вызывает его сокращение.
Химические тормозные синапсы. Эти синапсы по механизму передачи возбуждения сходны с синапсами возбуждающего действия. тормозных синапсах медиатор (например, глицин) взаимодействует с рецепторами субсинаптической мембраны и открывает в ней хлорные каналы, это приводит к движению ионов хлора по концентрационному градиенту внутрь клетки и развитию гиперполяризации на субсинаптической: мембране. Возникает так называемый тормозной постсинаптический потенциал (ТПСП).
Ранее полагали, что каждому медиатору соответствует специфическая реакция постсинаптической клетки - возбуждение или торможение в той или иной форме. В настоящее время установлено, что одному медиатору чаще всего соответствует не один, а несколько различных рецепторов. Например, ацетилхолин в нервно-мышечных синапсах скелетных мышц действует на Н-холинорецепторы (чувствительные к никотину), которые открывают широкие каналы для натрия (и калия), что порождает ВПСП (ПКП) В ваго-сердечных синапсах тот же ацетилхолин действует на М-холинорецепторы (чувствительные к мускарину), открывающие селективные каналы для ионов калия, поэтому здесь генерируется тормозной постсинаптический потенциал (ТПСП). Следовательно, возбуждающий или тормозной характер действия медиатора определяется свойствами субсинаптической мембраны (точнее, видом рецептора), а не самого медиатора.
Физиологические свойства химических синапсов. Синапсы с химической передачей возбуждения обладают рядом общих свойств:
• Возбуждение через синапсы проводится только в одном направлении (односторонне). Это обусловлено строением синапса: медиатор выделяется только из пресинаптического утолщения и взаимодействует с рецепторами субсинаптической мембраны;
• передача возбуждения через синапсы осуществляется медленнее, чем по нервному волокну - синаптическая задержка;
• передача возбуждения осуществляется с помощью специальных химических посредников - медиаторов;
• в синапсах происходит трансформация ритма возбуждения;
• синапсы обладают низкой лабильностью;
• синапсы обладают высокой утомляемостью;
• синапсы обладают высокой чувствительностью к химическим (в том числе и к фармакологическим) веществам.
Электрические синапсы возбуждающего действия. Кроме синапсов с химической передачей возбуждения преимущественно в центральной нервной системе (ЦНС) встречаются синапсы с электрической передачей. Возбуждающим электрическим синапсам свойственны очень узкая синаптическая щель и очень низкое удельное сопротивление сближенных пре- и постсинаптических мембран, что обеспечивает эффективное прохождение локальных электрических токов. Низкое сопротивление, как правило, связано с наличием поперечных каналов, пересекающих обе мембраны, т. е. идущих из клетки в клетку (щелевой контакт). Каналы образуются белковыми молекулами (полумолекулами) каждой из контактирующих мембран, которые соединяются комплементарно. Эта структура легко проходима для электрического тока.
Схема передачи возбуждения в электрическом синапсе: ток, вызванный пресинаптическим потенциалом действия, раздражает по-стсинаптическую мембрану, где возникает ВПСП и потенциал действия.
Поперечные каналы объединяют клетки не только электрически, но и химически, так как они проходимы для многих низкомолекулярных соединений. Поэтому возбуждающие электрические синапсы с поперечными каналами формируются, как правило, между клетками одного типа (например, между клетками сердечной мышцы).
Общими свойствами возбуждающих электрических синапсов являются:
• быстродействие (значительно превосходит таковое в химических синапсах);
• слабость следовых эффектов при передаче возбуждения (в результате этого в них практически невозможна суммация последовательных сигналов);
• высокая надежность передачи возбуждения.
Возбуждающие электрические синапсы могут возникать при благоприятных условиях и исчезать при неблагоприятных. Например, при повреждении одной из контактирующих клеток ее электрические синапсы с другими клетками ликвидируются. Это свойство называется пластичностью.
Электрические синапсы могут быть с односторонней и двусторонней передачей возбуждения.
Электрический тормозный синапс. Наряду с электрическими синапсами возбуждающего действия могут встречаться электрические тормозные синапсы. Примером такого синапса может служить синапс, который образует нервное окончание на выходном сегменте маутнеровского нейрона у рыб. Тормозящее влияние возникает за счет действия тока, вызванного потенциалом действия пресинапти-ческой мембраны. Пресинаптический потенциал вызывает значительную гиперполяризацию сегмента и гиперполяризующий ток мгновенно тормозит генерацию потенциала действия в начальном сегменте аксона.
В смешанных синапсах Пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Область контакта между двумя нейронами называют синапсом.
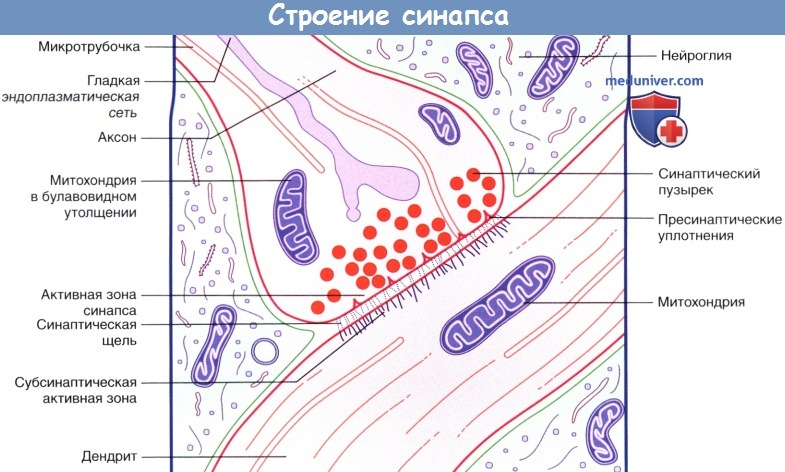
Внутреннее строение аксодендритического синапса.
а) Электрические синапсы. Электрические синапсы в нервной системе млекопитающих встречаются редко. Они образованы щелевидными контактами (нексусами) между дендритами или сомами соприкасающихся нейронов, которые соединяются с помощью цитоплазматических каналов диаметром 1,5 нм. Процесс передачи сигнала происходит без синаптической задержки и без участия медиаторов.
Посредством электрических синапсов возможно распространение электротонических потенциалов от одного нейрона к другому. Вследствие тесного синаптического контакта модуляция проведения сигнала невозможна. Задача этих синапсов — осуществление одновременного возбуждения нейронов, выполняющих одинаковую функцию. Примером служат нейроны дыхательного центра продолговатого мозга, которые во время вдоха синхронно генерируют импульсы. Кроме того, примером могут служить нейронные цепи, управляющие саккадами, при которых точка фиксации взора перемещается от одного объекта внимания к другому.
б) Химические синапсы. Большинство синапсов нервной системы — химические. Функционирование таких синапсов зависит от высвобождения медиаторов. Классический химический синапс представлен пресинаптической мембраной, синаптической щелью и постсинаптической мембраной. Пресинаптическая мембрана — часть булавовидного расширения нервного окончания клетки, передающей сигнал, а постсинаптическая мембрана — часть клетки, получающей сигнал.
Медиатор высвобождается из булавовидного расширения посредством экзоцитоза, проходит через синаптическую щель и связывается с рецепторами на постсинаптической мембране. Под постсинаптической мембраной расположена субсинаптическая активная зона, в которой после активации рецепторов постсинаптической мембраны происходят разнообразные биохимические процессы.
В булавовидном расширении расположены содержащие медиаторы синаптические пузырьки, а также большое количество митохондрий и цистерны гладкой эндоплазматической сети. Применение традиционных методик фиксации при исследовании клеток позволяет различить на пресинаптической мембране пресинаптические уплотнения, ограничивающие активные зоны синапса, к которым при помощи микротрубочек направляются синаптические пузырьки.
Аксодендритический синапс.
Срез препарата спинного мозга: синапс между концевым участком дендрита и, предположительно, двигательным нейроном.
Наличие округлых синаптических пузырьков и постсинаптического уплотнения характерно для возбуждающих синапсов.
Срез дендрита проведен в поперечном направлении, о чем свидетельствует наличие множества микротрубочек.
Кроме того, видны некоторые нейрофиламенты. Участок синапса окружен протоплазматическим астроцитом. Процессы, происходящие в нервных окончаниях двух типов.
(А) Синаптическая передача небольших молекул (например, глутамата).
(1) Транспортные пузырьки, содержащие мембранные белки синаптических пузырьков, направляются вдоль микротрубочек к плазматической мембране булавовидного утолщения.
В это же время происходит перенос молекул ферментов и глутамата путем медленного транспорта.
(2) Мембранные белки пузырьков выходят из плазматической мембраны и формируют синаптические пузырьки.
(3) Глутамат погружается в синаптические пузырьки; происходит накопление медиатора.
(4) Пузырьки, содержащие глутамат, подходят к пресинаптической мембране.
(5) В результате деполяризации происходит экзоцитоз медиатора из частично разрушенных пузырьков.
(6) Высвобождающийся медиатор распространяется диффузно в области синаптической щели и активирует специфические рецепторы на постсинаптической мембране.
(7) Мембраны синаптических пузырьков транспортируются обратно в клетку путем эндоцитоза.
(8) Происходит частичный обратный захват глутамата в клетку для повторного использования.
(Б) Передача нейропептидов (например, субстанции Р), осуществляющаяся одновременно с синаптической передачей (например, глутамата).
Совместная передача этих веществ происходит в центральных нервных окончаниях униполярных нейронов, обеспечивающих болевую чувствительность.
(1) Синтезированные в комплексе Гольджи (в области перикариона) пузырьки и предшественники пептидов (пропептиды) транспортируются к булавовидному расширению путем быстрого транспорта.
(2) При их попадании в область булавовидного утолщения завершается процесс формирования молекулы пептида, и пузырьки транспортируются к плазматической мембране.
(3) Деполяризация мембраны и перенос содержимого пузырьков в межклеточное пространство путем экзоцитоза.
(4) Одновременно с этим происходит высвобождение глутамата.
1. Активация рецепторов. Молекулы медиаторов проходят через синаптическую щель и активируют рецепторные белки, расположенные парами на постсинаптической мембране. Активация рецепторов запускает ионные процессы, которые приводят к деполяризации постсинаптической мембраны (возбуждающее постсинаптическое действие) или гиперполяризации постсинаптической мембраны (тормозящее постсинаптическое действие). Изменение электротонуса передается в сому в виде затухающего по мере распространения электротонического потенциала, за счет которого происходит изменение потенциала покоя в начальном сегменте аксона.
Ионные процессы подробно описаны в отдельной статье на сайте. При преобладании возбуждающих постсинаптических потенциалов начальный сегмент аксона деполяризуется до порогового уровня и генерирует потенциал действия.
Наиболее распространенный возбуждающий медиатор ЦНС — глутамат, а тормозной — гамма-аминомасляная кислота (ГАМК). В периферической нервной системе медиатором для двигательных нейронов поперечно-полосатой мускулатуры служит ацетилхолин, а для чувствительных нейронов — глутамат.
Последовательность процессов, происходящих в глутаматергических синапсах, показана на рисунке ниже. При передаче глутамата совместно с другими пептидами высвобождение пептидов осуществляется внесинаптическим путем.
Большинство чувствительных нейронов помимо глутамата выделяет и другие пептиды (один или несколько), высвобождающиеся в различных участках нейрона; однако основная функция этих пептидов — модуляция (повышение или снижение) эффективности синаптической передачи глутамата.
Кроме того, нейротрансмиссия может происходить путем диффузной внесинаптической передачи сигнала, характерной для моноаминергических нейронов (нейронов, использующих биогенные амины для обеспечения нейротрансмиссии). Выделяют две разновидности моноаминергических нейронов. В одних нейронах осуществляется синтез катехоламинов (норадреналина или дофамина) из аминокислоты тирозина, а в других — серотонина из аминокислоты триптофана. Например, дофамин высвобождается как в синаптической области, так и из варикозных утолщений аксона, в которых также происходит синтез этого нейромедиатора.
Дофамин проникает в межклеточную жидкость ЦНС и до момента деградации способен активировать специфические рецепторы на расстоянии до 100 мкм. Моноаминергические нейроны присутствуют во многих структурах ЦНС; нарушение передачи импульса этими нейронами приводит к различным заболеваниям, среди которых выделяют болезнь Паркинсона, шизофрению и глубокую депрессию.
Оксид азота (газообразная молекула) также участвует в диффузной нейропередаче в глутаматергической системе нейронов. Избыточное влияние оксида азота оказывает цитотоксическое действие, особенно в тех участках, кровоснабжение которых нарушено за счет тромбоза артерий. Глутамат также является потенциально цитотоксическим нейромедиатором.
Существует три вида химических межнейрональных взаимодействий: синаптическое (например, глутаматергическое), внесинаптическое (пептидергическое) и диффузное (например, моноаминергическое, серотонинергическое).
Химические синапсы классифицируют по анатомическому строению на аксодендритические, аксосоматические, аксоаксональные и дендро-дендритические. Синапс представлен пре- и постсинаптическими мембранами, синаптической щелью и субсинаптической активной зоной.
Электрические синапсы обеспечивают одновременную активацию целых групп нейронов, образуя между ними электрические связи за счет щелевидных контактов (нексусов).
Диффузная нейротрансмиссия в головном мозге.
Аксоны глутаматергического (1) и дофаминергического (2) нейронов образуют плотные синаптические контакты с отростком звездчатого нейрона (3) полосатого тела.
Дофамин высвобождается не только из пресинаптической области, но и из варикозного утолщения аксона, откуда диффузно распространяется в межклеточное пространство и активирует дофаминовые рецепторы дендритного ствола и стенки перицита капилляра. Растормаживание.
(А) Возбуждающий нейрон 1 активирует тормозной нейрон 2, который в свою очередь затормаживает нейрон 3.
(Б) Появление второго тормозного нейрона (2б) оказывает противоположное влияние на нейрон 3, поскольку происходит торможение нейрона 2б.
Спонтанно-активный нейрон 3 генерирует сигналы в условиях отсутствия тормозных влияний.
3. Торможение и растормаживание. Функционирование спонтанно-активных нейронов сдерживается под влиянием тормозных нейронов (обычно, ГАМКергических). Деятельность тормозных нейронов, в свою очередь, может быть ингибирована воздействующими на них другими тормозными нейронами, в результате чего происходит растормаживание клетки-мишени. Процесс растормаживания — важная особенность нейрональной активности в базальных ганглиях.
4. Редкие виды химических синапсов. Выделяют два типа аксоаксональных синапсов. В обоих случаях булавовидное утолщение образует тормозной нейрон. Синапсы первого типа образуются в области начального сегмента аксона и передают мощное ингибирующее влияние тормозного нейрона. Синапсы второго типа образуются между булавовидным утолщением тормозного нейрона и булавовидными утолщениями возбуждающих нейронов, что приводит к угнетению высвобождения медиаторов. Этот процесс получил название пресинаптического торможения. В этом плане традиционный синапс обеспечивает постсинаптичсекое торможение.
Дендро-дендритические (Д-Д) синапсы образуются между дендритными шипиками дендритов смежных шипиковых нейронов. Их задача — не генерирование нервного импульса, а изменение электротонуса клетки-мишени. В последовательных Д-Д-синапсах синаптические пузырьки располагаются только в одном дендритном шипике, а в реципрокном Д-Д-синапсе— в обоих. Возбуждающие Д-Д-синапсы изображены на рисунке ниже. Тормозные Д-Д-синапсы широко представлены в переключающих ядрах таламуса.
Кроме того, выделяют немногочисленные сомато-дендритические и сомато-соматические синапсы.
Аксоаксональные синапсы коры головного мозга.
Стрелками указано направление проведения импульсов. (1) Пресинаптическое и (2) постсинаптическое торможение спинномозгового нейрона, направляющегося к головному мозгу.
Стрелками указано направление проведения импульсов (возможно торможение переключательного нейрона под действием тормозных влияний). Возбуждающие дендро-дендритические синапсы. Изображены дендриты трех нейронов.
Реципрокный синапс (справа). Стрелками указано направление распространения электрото-нических волн.
Читайте также: