
Правило непрерывности хромосом кратко
Обновлено: 04.07.2024
Вопрос 1.
Деление клетки бывает прямое (митоз) и непрямое (амитоз).
Прямое деление происходит в результате деления ядра и цитоплазмы клетки на две части. Непрямое деление происходит в несколько этапов или фаз. Ему предшествует подготовительный период — интерфаза. В этот период клетка растет, накапливает энергию, хромосомы удваиваются.
Фазы непрямого деления:
I. Ядерная оболочка исчезает; хромосомы спирализуются; клеточный центр образует веретено деления.
II. Хромосомы выстраиваются по экватору клетки; нити веретена деления от каждого полюса соединяются с хромосомами.
III. Хромосомы расходятся к полюсам клетки; нити веретена деления исчезают.
IV. Формируется ядерная оболочка; хромосомы деспирализуются; в цитоплазме появляется перетяжка. Образуются две дочерние клетки с таким же набором хромосом, как и у материнской (исходной) клетки.
Вопрос 2.
Хромосомы (Рис. 1.).
Хромосома это пара хроматид.
Материнская хроматида - это и есть дочерняя хромосома.
В хромосоме имеются эу- и гетерохроматиновые участки. Диффузный или деконденсированный хроматин – эухроматин – он генетически активен, т.к. с него может идти транскрипция. Конденсированные участки хроматина – гетерохроматин – это неактивные участки хромосом. Чередование эу- и гетерохроматиновых участков используют для идентификации хромосом.
Хромосома на стадии метафазы имеет вид нитей или палочек, максимально спирализованных и состоящих из двух хроматид, соединенных первичной перетяжкой или центромерой. В центральной части центромеры находятся кинетохоры, к которым во время митоза прикрепляются микротрубочки веретена деления. Некоторые хромосомы имеют вторичную перетяжку или ядрышковый организатор, контролирующий образование ядрышек.
Рис. 1. Строение хромосомы
Строение хромосом.
В зависимости от места расположения центромеры относительно плеч различают типы хромосом (Рис. 2.):
Равноплечие или метацентрические, когда центромера делит хромосому пополам, т.е. плечи равные или почти равные.
Неравноплечие или субметацентрические, т.е. плечи неодинаковоой длины.
Палочковидные или акроцентрические, когда одно плечо короткое или почти незаметное.
Некоторые виды могут иметь еще и одноплечие хромосомы или телоцентрические.
В кариотипе человека есть метацетрические, субметацентрические и акроцентрические хромосомы, а телоцентрические образуются только в результате мутаций.
Правила хромосом.
1. Постоянства числа хромосом. Каждый вид имеет постоянное число хромосом. Например: человек - 46, ясень обыкновенный – 46, дрозофила – 8, шимпанзе - 48, таракан – 48, сладкий перец - 48.
2. Парности хромосом. Хромосома имеет свою гомологичную пару, т.е. число хромосом четное. Например: у человека - 23 пары и т.д.
3. Индивидуальности. Каждая хромосома имеет свою форму, величину и совокупность генов.
4. Непрерывности или преемственности. Каждая хромосома образуется от хромосомы в результате репликации.
Уровень организации вида не зависит от количества хромосом, а зависит от наследственной информации. Парные хромосомы называют гомологичными. Одна хромосома в паре происходит от материнского организма, другая - от отцовского. Хромосомы из разных пар называют негомологичными.
Вопрос 3.
В результате непрямого деления образуются клетки, похожие друг на друга и на материнскую клетку. Благодаря этому типу деления происходит:
Генетическая стабильность, т.к. хроматиды образуются в результате репликации, т.е. наследственная информация их идентична материнской.
Рост организмов, т.к. в результате митоза число клеток увеличивается.
Бесполое размножение – многие виды растений и животных размножаются в результате митотического деления.
Регенерация и замещение клеток идет за счет митозов.
Вопрос 4.
Все живые организмы на нашей планете (кроме вирусов) состоят из клеток. Клетки растений, животных и человека имеют общий план строения: мембрана, цитоплазма, ядро. Каждый органоид в клетке выполняет определенную функцию, обеспечивая жизнедеятельность клетки в целом. Несмотря на то что клетки имеют различия в строении, каждая из них обладает рядом жизненных свойств, общих для клеток растений, животных и человека. клетки многоклеточных организмов специализированы: они выполняют разные функции и образуют ткани. Это доказывает единство происхождения всего живого на нашей планете.
Примечание: половых клетках хромосомы непарные и их число в два раза меньше. Они составляют одинарные, или гаплоидный, набор, его обозначают n.Гаплоидный набор содержится в ядрах половых клеток (гамет). Если у человека диплоидный набор - 46 хромосом, то гаплоидный – 23.
В ядрах гамет из каждой пары гомологичных хромосом присутствует только одна 1 хромосома. При оплодотворении женских и мужских гамет образуется зигота.
Все хромосомы подразделяются на аутосомы и половые хромосомы. В соматических клетках присутствует двойной - диплоидный набор хромосом, в половых - гаплоидный (одинарный).
Ядрышки – непостоянная структура ядра. Они исчезают вначале деления клетки и вновь появляются к концу его. Они представляют собой мелкие тельца шаровидной формы. Ядрышки имеют большую плотность, чем ядро. В них происходит синтез рибосомальной и других видов РНК, а также образование рибосом.
Обычно в ядре клетки бывает от одного до семи ядрышек. Они хорошо видны между делениями клетки, а во время деления – разрушаются.
Функции ядра
Функция ядра связана с хромосомами.
1) Хранение генетической информации.
2) Защита генетической информации от мутагенов.
3) Синтез ДНК и всех видов РНК
4) Формирование рибосом.
5) Деление ядра лежит в основе размножения клеток.
СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОГО ЯДРА
План
Строение ядра
Функции ядра
Ядро –самый крупный клеточный компонент, хорошо видимый в световой микроскоп. Поэтому оно было открыто первым среди клеточных структур. Ядро имеется во всех клетках эукариот, за исключением зрелых эритроцитов млекопитающих.
Ядро было открыто и описано в 1833 г английским ботаником Р. Броуном.
Обычно в клетках содержится одно ядро (диаметром от 1 мкм до 1мм), но иногда встречаются многоядерные клетки. Форма ядра шаровидная или овальная.
Размер ядер зависит от количества цитоплазмы. Он может меняться под влиянием условий внешней среды и при усилении функций клетки.
Строение ядра
Ядро окружено ядерной оболочкой, отделяющей его от цитоплазмы, содержит кариоплазму (ядерный сок), одно или несколько ядрышек, хроматин.
Ядерная оболочка интерфазного ядра состоит из двух мембран (наружней и внутренней), между которыми имеется перинуклеарное пространство. В мембранах имеются поры. Через них идут обменные процессы между ядром и цитоплазмой. Наружная мембрана может переходить в стенки каналов эндоплазматической сети. На наружной ядерной мембране располагаются рибосомы.
Ядерный сок (карилимфа) – это внутреннее содержимое ядра, представляющее собой раствор белков, нуклеотидов, АТФ т различные виды РНК. В нем присутствуют также фибриллярные белки. В кариолимфе находятся ядрышки и хроматин. Ядерный сок обеспечивает взаимосвязь компонентов ядра и нормальное функционирование генетического материала.
Ядрышки – непостоянная структура ядра. Они исчезают вначале деления клетки и вновь появляются к концу его. Они представляют собой мелкие тельца шаровидной формы. Ядрышки имеют большую плотность, чем ядро. В них происходит синтез рибосомальной и других видов РНК, а также образование рибосом.
Возникновение ядрышек связано с определенными зонами хромосом, называемыми ядрышковыми организаторами. От последних зависит число ядрышек. В них содержится гены рРНК.
В жизни ядра различают два периода – между делениями (интерфаза, подготовка к делению) и во время деления клетки. В разные периоды внутреннее строение ядра меняется. Между делениями клетки ядро содержит ядерный сок и нити хроматина.
Хроматин (окрашенный материал) – плотное вещество ядра, хорошо окрашиваемое основными красителями. По своей химической природе хроматин – нуклеопротеид. Он представляет собой длинные тонкие нити ДНК, образующие комплекс с особыми белками гистонами (белки с повышенным содержанием положительно заряженных аминокислотных остатков лизина и аргинина). Различают гетерохроматин и эухроматин. Рыхлоспирализованный хроматин называют эухроматином (неокрашивается красителем). Во время деления клетки происходит спирализация хроматина. Он превращается в гетерохроматин (плотно упакованный), который хорошо окрашивается и виден в микроскоп в виде темных полос. До деления клетки нити хроматина раскручены. Во время деления они скручиваются, что сопровождается их утолщением и укорачиванием. В результате формируются особые образования – хромосомы. Белковые молекулы увеличивают толщину хромосомы и служат защитной оболочкой для ДНК. Толщина хромосомы от 100 до 200нам, диаметр ДНК 2 нм.

Хромосомы способны к размножению – ауторепродукции, т.е. образованию себе подобных.
Примечание: половых клетках хромосомы непарные и их число в два раза меньше. Они составляют одинарные, или гаплоидный, набор, его обозначают n.Гаплоидный набор содержится в ядрах половых клеток (гамет). Если у человека диплоидный набор - 46 хромосом, то гаплоидный – 23.
В ядрах гамет из каждой пары гомологичных хромосом присутствует только одна 1 хромосома. При оплодотворении женских и мужских гамет образуется зигота.
Все хромосомы подразделяются на аутосомы и половые хромосомы. В соматических клетках присутствует двойной - диплоидный набор хромосом, в половых - гаплоидный (одинарный).
Ядрышки – непостоянная структура ядра. Они исчезают вначале деления клетки и вновь появляются к концу его. Они представляют собой мелкие тельца шаровидной формы. Ядрышки имеют большую плотность, чем ядро. В них происходит синтез рибосомальной и других видов РНК, а также образование рибосом.
Обычно в ядре клетки бывает от одного до семи ядрышек. Они хорошо видны между делениями клетки, а во время деления – разрушаются.
Функции ядра
Функция ядра связана с хромосомами.
1) Хранение генетической информации.
2) Защита генетической информации от мутагенов.
3) Синтез ДНК и всех видов РНК
4) Формирование рибосом.
5) Деление ядра лежит в основе размножения клеток.
СТРОЕНИЕ И ФУНКЦИИ КЛЕТОЧНОГО ЯДРА
План
Строение ядра
Функции ядра
Ядро –самый крупный клеточный компонент, хорошо видимый в световой микроскоп. Поэтому оно было открыто первым среди клеточных структур. Ядро имеется во всех клетках эукариот, за исключением зрелых эритроцитов млекопитающих.
Ядро было открыто и описано в 1833 г английским ботаником Р. Броуном.
Обычно в клетках содержится одно ядро (диаметром от 1 мкм до 1мм), но иногда встречаются многоядерные клетки. Форма ядра шаровидная или овальная.
Размер ядер зависит от количества цитоплазмы. Он может меняться под влиянием условий внешней среды и при усилении функций клетки.
Строение ядра
Ядро окружено ядерной оболочкой, отделяющей его от цитоплазмы, содержит кариоплазму (ядерный сок), одно или несколько ядрышек, хроматин.
Ядерная оболочка интерфазного ядра состоит из двух мембран (наружней и внутренней), между которыми имеется перинуклеарное пространство. В мембранах имеются поры. Через них идут обменные процессы между ядром и цитоплазмой. Наружная мембрана может переходить в стенки каналов эндоплазматической сети. На наружной ядерной мембране располагаются рибосомы.
Ядерный сок (карилимфа) – это внутреннее содержимое ядра, представляющее собой раствор белков, нуклеотидов, АТФ т различные виды РНК. В нем присутствуют также фибриллярные белки. В кариолимфе находятся ядрышки и хроматин. Ядерный сок обеспечивает взаимосвязь компонентов ядра и нормальное функционирование генетического материала.
Ядрышки – непостоянная структура ядра. Они исчезают вначале деления клетки и вновь появляются к концу его. Они представляют собой мелкие тельца шаровидной формы. Ядрышки имеют большую плотность, чем ядро. В них происходит синтез рибосомальной и других видов РНК, а также образование рибосом.
Возникновение ядрышек связано с определенными зонами хромосом, называемыми ядрышковыми организаторами. От последних зависит число ядрышек. В них содержится гены рРНК.
В жизни ядра различают два периода – между делениями (интерфаза, подготовка к делению) и во время деления клетки. В разные периоды внутреннее строение ядра меняется. Между делениями клетки ядро содержит ядерный сок и нити хроматина.
Хроматин (окрашенный материал) – плотное вещество ядра, хорошо окрашиваемое основными красителями. По своей химической природе хроматин – нуклеопротеид. Он представляет собой длинные тонкие нити ДНК, образующие комплекс с особыми белками гистонами (белки с повышенным содержанием положительно заряженных аминокислотных остатков лизина и аргинина). Различают гетерохроматин и эухроматин. Рыхлоспирализованный хроматин называют эухроматином (неокрашивается красителем). Во время деления клетки происходит спирализация хроматина. Он превращается в гетерохроматин (плотно упакованный), который хорошо окрашивается и виден в микроскоп в виде темных полос. До деления клетки нити хроматина раскручены. Во время деления они скручиваются, что сопровождается их утолщением и укорачиванием. В результате формируются особые образования – хромосомы. Белковые молекулы увеличивают толщину хромосомы и служат защитной оболочкой для ДНК. Толщина хромосомы от 100 до 200нам, диаметр ДНК 2 нм.


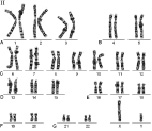
Рис. 1. Мужской кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
Как указано выше, кариотип человека состоит из 46 хромосом, которые нумеруются от 1 до 22 (аутосомы) и делятся на 7 групп, – A, B, C, D, E, F, G и половые хромосомы (гоносомы) X и Y (рис. 1 и 2).
К первой группе А относятся хромосомы 1, 2 и 3, которые хорошо отличаются друг от друга. Хромосома 1 (размер – 11 мкм) – метацентрическая, содержит вторичную перетяжку в околоцентромерном участке длинного плеча. Хромосома 2 (10,8 мкм) по размерам почти равна хромосоме 1 и является субметацентрической. Хромосома 3 (размер – 8,3 мкм) – практически метацентрическая.
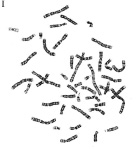
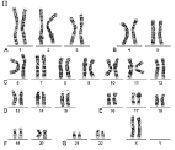
Рис. 2. Женский кариотип: I – метафазная пластинка; II – классификация по группам и нумерация хромосом
К группе В относятся хромосомы 4 и 5 (размер – 7,7 мкм каждая) – это крупные субметацентрические хромосомы, которые не отличаются друг от друга при рутинном окрашивании ни размером, ни положением центромер.
К группе С относятся хромосомы с 6 по 12 и Х. В основном, это субметацентрические хромосомы крупных и средних размеров. Наиболее крупные хромосомы из группы С – 6, 7 и Х (6,8–7,2 мкм). Хромосома Х является половой хромосомой (гоносомой). Хромосома 7 более метацентрична, чем хромосома 6. Хромосомы 8 и 9 – практически одинаковы по размеру (5,8 мкм). Хромосома 8 метацентричнее хромосомы 9, которая характеризуется регулярной вторичной перетяжкой в прицентромерном районе длинного плеча.
К группе D относятся хромосомы 13, 14 и 15 (4,2 мкм) – средних размеров акроцентрические хромосомы с почти терминальным расположением центромеры. Эти хромосомы между собой не различаются ни по размерам, ни морфологически после рутинного окрашивания. Короткое плечо всех трёх пар хромосом может формировать спутники (рис. 1, 2 и 5).
К группе Е относятся хромосомы 16, 17 и 18. Хромосома 16 (размер – 3,6 мкм) – сравнительно небольшая метацентрическая хромосома, содержащая вторичную перетяжку в длинном плече. Хромосома 17 (размер – 3,5 мкм) – сравнительно короткая субметацентрическая хромосома. Хромосома 18 (размер – 3,2 мкм) – самая короткая субметацентрическая хромосома.
Группа F представлена хромосомами 19 и 20 (размеры – 2,9 мкм). Это короткие метацентрические хромосомы, которые не отличаются между собой без дифференциального окрашивания по длине.
Хромосомы 21 и 22 (2,8 мкм) относятся к группе G. Это наиболее короткие акроцентрические хромосомы в кариотипе, которые обладают способностью формировать спутники на коротком плече (рис. 1, 2 и 5).
Хромосома Y (2,3 мкм) является маленькой акроцентрической хромосомой, сравнимой по размерам с хромосомами 21 и 22, но не имеющая спутников.
Следует отметить, что хромосомные синдромы и аномалии связаны с хромосомными (геномными) мутациями (аномалиями) в виде различных структурных перестроек хромосом или с изменением их числа (n). Численные изменения хромосом могут быть двух типов: полиплоидии – умножение хромосомного набора (3n, 4n и т. д.) или генома, кратное гаплоидному числу хромосом; анеуплоидии – увеличение или уменьшение числа хромосом, некратное гаплоидному. Структурные хромосомные (геномные) перестройки классифицируют по принципу линейной последовательности расположения генов: делеции (потеря хромосомных участков), дупликации (удвоение хромосомных участков), инверсии (перевертывание на 180° относительно нормальной последовательности хромосомных участков), инсерции (вставки хромосомных участков) и транслокации (изменение расположения хромосомных участков). Подробная информация по возможным аномалиям хромосом человека представлена в главе 3.5.

Правило непрерывности хромосом: в последовательных генерациях число и индивидуальность хромосом сохраняется благодаря способности хромосом к авторепродукции во время деления клетки.
18. Кариотип человека. Его определение. Кариограмма, принцип составления. Идиограмма, ее содержание.
1956 год - шведы Тио и Леван, англичане Форд и Хамертон установили, что ядро диплоидной клетки человека содержит 46 хромосом – это хромосомный набор или кариотип человека
Совокупность всех метафазных хромосом, расположенных относительно произвольно в экваториальной плоскости клетки, именуется метафазной пластинкой или просто хромосомным набором. После приготовления препаратов хромосом, которые можно приготовить из всех тканей и клеточных суспензий, содержащих делящиеся клетки (в зависимости от целей важно, конечно, количество метафаз), хромосомы окрашиваются, ибо только после этого их можно различить в световой микроскоп, получить микрофотографию, идентифицировать и, расположив их в определенном порядке, т.е. составив кариограмму, получить целостное представление о кариотипе конкретного человека. Кариограмма- это те же хромосомы метафазной пластинки, но расположенные упорядоченно. Принцип упорядоченности общий для всего вида и определяется идеограммой. Идиограмма - это графическое изображение гаплоидного набора хромосом (можно и диплоидного) и расположение их по группам в зависимости от формы и величины. Группы располагаются в порядке уменьшения величины входящих в них хромосом.
В современных цитогенетических лабораториях процесс составления кариограммы компьютеризирован.
Наиболее простой способ окрашивания хромосом красителем Гимза или 2%-ым ацетоорсеином, или 2%-ым ацетокармином. При этом хромосомы окрашиваются целиком, равномерно и интенсивно.
19. Денверская классификация хромосом и их Парижская номенклатура.
Наиболее простой способ окрашивания хромосом красителем Гимза или 2%-ым ацетоорсеином, или 2%-ым ацетокармином. При этом хромосомы окрашиваются целиком, равномерно и интенсивно. Окрашенные таким образом хромосомы, согласно Денверской классификации (I960), располагались в идиограмме в зависимости от их длины и нумеровались по парам от 1 до 23. Тогда же Патау предложил разбить 23 пары хромосом на 7 групп от А до G с учетом расположения центромеры. Важным признаком, уточняющим форму хромосомы, стал центромерный индекс: отношение длины короткого плеча к длине всей хромосомы, выраженное в %. Комплекс этих параметров позволял с немалой степенью точности распределить хромосомы по группам, но идентифицировать их, особенно в группах В, С, D, F и G, было невозможно.
Однако уже при стандартном (рутинном) равномерном окрашивании хромосом замечали, но оставили без внимания, некоторую неоднородность в плотности окрашивания по длине хромосом. И только позже (1968 г.), когда Касперсон с сотрудниками обнаружили, что после обработки акрихин-ипритом флуоресценция по длине хромосом распределена не равномерно, а в виде сегментов, они показали, что каждую хромосому можно надежно идентифицировать с помощью такого метода дифференциального окрашивания, ибо расположение сегментов для каждой хромосомы строго специфично. Вскоре стало ясно, что очень сходный рисунок сегментации хромосом можно получить и с помощью красителя Гимза, дополнив окрашивание некоторыми приемами. Впоследствии при разных способах обработки хромосом были обнаружены разные типы сегментов.
На Парижской конференции по стандартизации и номенклатуре хромосом человека (1971) все полученные к тому времени данные по дифференциальному окрашиванию хромосом были сопоставлены и оказалось, что все методы в принципе выявляют одни и те же структуры, но каждый специфичен в отношении определенных сегментов. И обозначать различные типы сегментов решили по методам, с помощью которых они выявляются.
Q - сегменты - флуоресцирующие после окраски акрихин-ипритом;
G - сегменты (Гимза) - выявляются при окрашивании красителем Гимза в сочетании с дополнительными процедурами; Q и G сегменты идентичны, но в большинстве лабораторий предпочитают этот метод, т.к. он не требует использования флуоресцентного микроскопа и эти препараты дольше хранятся; однако, только с помощью Q-метода можно идентифицировать Y-хромосому человека даже в интерфазном ядре;
R - сегменты - окрашиваются после контролируемой тепловой денатурации, располагаются между Q и G - сегментами;
С - сегменты - конститутивный гетерохроматин, располагается в прицентромерных районах обоих плечей хромосомы;
Т - сегменты - расположены в теломерных (концевых) районах хромосом.
Химическая природа дифференциального окрашивания еще только исследуется. Обсуждаются две основные гипотезы: первая исходит из того, что различные участки хромосом человека отличаются по количественному содержанию пар оснований аденин - тимин и гуанин - цитозин. Отсюда разная степень усвоения ими красителей. В частности, блоки с большим содержанием пар А-Т связываются преимущественно с акрихин-ипритом, следовательно, Q-сегменты соответствуют участкам, богатым А-Т - парами; R-сегменты соответствуют участкам, богатым Г-Ц - парами, которые более устойчивы к тепловой денатурации - это, однако, не объясняет всех особенностей сегментации хромосом. Вторая гипотеза, белковая, исходит из данных о том, что предварительная протеолитическая обработка перед окрашиванием красителем Гимза индуцирует появление G-сегментов, а так как разные по составу участки ДНК связаны с разными белками, можно полагать, что рисунок сегментации зависит от особенностей комплекса ДНК - белок.
И все же, что собой представляют полосы - сегменты митотических хромосом, остается загадкой. Даже небольшие тонкие полосы содержат не менее 30 гигантских петель, суммарный нуклеотидный состав которых более 1 млн., нуклеотидов. Возможно, существование таких структурных блоков связано с функционированием эукариотического генома вообще, хотя сами по себе сегменты ничего конкретного о функционировании индивидуальных генов не говорят, ибо в самой тонкой полосе, которую еще можно различить, содержится от 10 до 100 генов. Но то, что картина распределения сегментов в хромосомах почти не изменилась за долгие периоды эволюции (почти каждая хромосома человека имеет своего аналога в кариотипе шимпанзе, гориллы, орангутана), свидетельствует о большом значении пространственной организации ДНК для экспрессии соответствующих генов.
Итак, информация, полученная в результате анализа дифференциально окрашенных хромосом, позволяет представить идиограмму хромосом человека следующим образом:
Группа В, 4 и 5 хромосомы - большие субметацентрические. ЦИ 24-30%, без дифференциального окрашивания друг от друга не отличаются.
Группа С, 6-12 хромосомы и Х-хромосома - средние Субметацентрические хромосомы 6, 7, 8,11 и 12 - относительно субметацентрические, ЦИ - 27-35; 11 и 12 обнаруживают очень сходный рисунок сегментации, однако 11-я хромосома более метацентрическая; в 9-й в длинном плече часто обнаруживают вторичную перетяжку, которая не окрашивается ни акрихином, ни красителем Гимза; Х-хромосома значительно варьирует по длине, в целом сходна с самыми длинными из С-группы, ЦИ - 40,12+2,12, отличить от других при стандартном окрашивании очень трудно.
Группа D, 13-15 хромосомы - акроцентрические, ЦИ около 15 -наименьший в кариотипе человека, все они могут иметь вторичную перетяжку на коротком плече или не иметь, а следовательно, иметь спутники или не иметь, спутники могут быть очень большими, а иногда двойными; короткие плечи этих хромосом содержат ядрышковый организатор.
Группа F, 19, 20 хромосомы - мелкие метацентрические, ЦИ - 36-46, при стандартной окраске выглядят одинаково, при дифференциальной - резко отличаются.
Группа G, 21, 22, Y-хромосомы - мелкие акроцентрические, ЦИ - 13-33; 21 и 22-ая могут иметь спутники, короткие плечи имеют ядрышковый организатор; Y-хромосома обычно (но не всегда) больше, хроматиды ее длинного плеча, как правило, лежат параллельно одна другой, а у 21 и 22 - ой хромосомы они чаще образуют широкий угол; спутники в Y-хромосоме отсутствуют, ЦИ от 0 до 26. В интерфазных ядрах дистальный участок длинного плеча при окрашивании акрихин-ипритом сильно флуоресцирует и выявляется как яркое пятно, которое называется Y-хроматин.
В соответствии с Парижской номенклатурой в хромосомах идиограммы показан рисунок сегментации (G - Q , R - сегменты) - позитивные светлые G (они же Q) сегменты, негативные темные - R, районы с варьирующей окраской заштриховываются. Латинскими буквами р и q обозначаются соответственно короткое и длинное плечо, в каждом плече выделяются районы, обозначенные арабскими цифрами, районы нумеруются от центромеры к теломерным участкам хромосомы. А уже внутри района выделяются сегменты(англ. bands), обозначенные арабскими цифрами по такому же принципу, т.е. сегмент имеет свой символ, например, 1q 32- второй сегмент третьего района в длинном плече 1 -ой хромосомы (при чтении справа налево).
Если в заключение сформулировать интегральную модель хромосомы, то она состоит из единственной двойной спирали ДНК, объединенной с гистонами в нуклеосомы. Некоторые районы этой двойной спирали представлены повторяющимися последовательностями, которые могут быть рассеяны по всему геному. Участки с повторяющимися последовательностями обнаруживают признаки конститутивного гетерохроматина. Участки с уникальными последовательностями пар нуклеотидов проявляют свойства эухроматина, это транскрибирующиеся участки - т.е. собственно гены, они соответствуют светлым G- и темным R-сегментам дифференциально окрашенных хромосом.
Биосинтез белка происходит в два этапа. В первый этап входит транскрипция и процессинг РНК, второй этап включает трансляцию. Во время транскрипции фермент РНК-полимераза синтезирует молекулу РНК, комплементарную последовательности соответствующего гена (участка ДНК). Терминатор в последовательности нуклеотидов ДНК определяет, в какой момент транскрипция прекратится. В ходе ряда последовательных стадий процессинга из мРНК удаляются некоторые фрагменты, и редко происходит редактирование нуклеотидных последовательностей. После синтеза РНК на матрице ДНК происходит транспортировка молекул РНК в цитоплазму. В процессе трансляции информация, записанная в последовательности нуклеотидов, переводится в последовательность остатков аминокислот.
Транскри́пция (от лат. transcriptio — переписывание) — процесс синтеза РНК с использованием ДНК в качестве матрицы, происходящий во всех живых клетках. Другими словами, это перенос генетической информации с ДНК на РНК.
Процессинг РНК
Процессинг РНК (посттранскрипционные модификации РНК) — совокупность процессов в клетках эукариот, которые приводят к превращению первичного транскрипта в зрелую РНК
(Транскрипт — молекула РНК, образующаяся в результате транскрипции (экспрессии соответствующего гена или участка ДНК)
Примерами транскриптов являются: мРНК, рРНК, тРНК, малые РНК
Между транскрипцией и трансляцией молекула иРНК претерпевает ряд последовательных изменений, которые обеспечивают созревание функционирующей матрицы для синтеза полипептидной цепочки
Трансляция
Трансляция (от лат. translatio — перевод) — процесс синтеза белка из аминокислот на матрице информационной (матричной) РНК (иРНК, мРНК), осуществляемый рибосомой.
У прокариот мРНК может считываться рибосомами в аминокислотную последовательность белков сразу после транскрипции, а у эукариот она транспортируется из ядра в цитоплазму, где находятся рибосомы. Скорость синтеза белков выше у прокариот и может достигать 20 аминокислот в секунду [1] . Процесс синтеза белка на основе молекулы мРНК называется трансляцией.
Рибосома содержит 2 функциональных участка для взаимодействия с тРНК: аминоацильный (акцепторный) и пептидильный (донорный). Аминоацил-тРНК попадает в акцепторный участок рибосомы и взаимодействует с образованием водородных связей между триплетами кодона и антикодона. После образования водородных связей система продвигается на 1 кодон и оказывается в донорном участке. Одновременно в освободившемся акцепторном участке оказывается новый кодон, и к нему присоединяется соответствующий аминоацил-т-РНК.
Во время начальной стадии биосинтеза белков, инициации, обычно метиониновый кодон узнаётся малой субъединицей рибосомы, к которой при помощи белковых факторов инициации присоединена метиониновая транспортная РНК (тРНК). После узнавания стартового кодона к малой субъединице присоединяется большая субъединица и начинается вторая стадия трансляции — элонгация. При каждом движении рибосомы от 5' к 3' концу мРНК считывается один кодон путём образования водородных связей между тремя нуклеотидами (кодоном) мРНК и комплементарным ему антикодоном транспортной РНК, к которой присоединена соответствующая аминокислота. Синтез пептидной связи катализируется рибосомальной РНК (рРНК), образующей пептидилтрансферазный центр рибосомы. Рибосомальная РНК катализирует образование пептидной связи между последней аминокислотой растущего пептида и аминокислотой, присоединённой к тРНК, позиционируя атомы азота и углерода в положении, благоприятном для прохождения реакции. Ферменты аминоацил-тРНК-синтетазы присоединяют аминокислоты к их тРНК. Третья и последняя стадия трансляции, терминация, происходит при достижении рибосомой стоп-кодона, после чего белковые факторы терминации гидролизуют последнюю тРНК от белка, прекращая его синтез. Таким образом, в рибосомах белки всегда синтезируются от N- к C-концу.
Число, размеры и форма хромосом являются специфическими признаками для каждого вида живых организмов. Так, в клетках рака-отшельника содержится по 254 хромосомы, а у комара – только 6. Соматические клетки человека содержат 46 хромосом. Совокупность всех структурных и количественных особенностей полного набора хромосом характерного для клеток конкретного вида живых организмов называется кариотипом.
Кариотип будущего организма формируется в процессе слияния двух половых клеток (сперматозоида и яйцеклетки). При этом объединяются их хромосомные наборы. Ядро зрелой половой клетки содержит половинный набор хромосом (для человека – 23). Подобный одинарный набор хромосом, аналогичный таковому в половых клетках, называется гаплоидным и обозначается – п. При оплодотворении яйцеклетки сперматозоидом в новом организме воссоздается специфический для данного вида кариотип, включающий у человека 46 хромосом. Полный состав хромосом обычной соматической клетки является диплоидным (2п).
В диплоидном наборе каждая хромосома имеет аналогичную по размеру и расположению центромеры другую парную хромосому. Такие хромосомы называются гомологичными. Гомологичные хромосомы не только похожи друг на друга, но и содержат гены, отвечающие за одни и те же признаки.
Правила хромосом
Существует 4 правила хромосом:
Правило постоянства числа хромосом. Соматические клетки организма каждого вида в норме имеют строго определенное число хромосом (например, у человека – 46, у дрозофилы – 8).
Правило парности хромосом. Каждая хромосома в диплоидном наборе имеет гомологичную - сходную по размерам, расположению центромеры и содержанию генов.
Правило индивидуальности хромосом. Каждая пара хромосом отличается от другой пары размерами, расположением центромеры и содержанием генов.
Правило непрерывности хромосом. В процессе удвоения генетического материала новая молекула ДНК синтезируется на основе информации старой молекулы ДНК (реакция матричного синтеза – каждая хромосома происходит от хромосомы).
Классификация хромосом человека
Хромосомы подразделяют на аутосомы (одинаковые у обоих полов) и гетерохромосомы, или половые хромосомы (разный набор у мужских и женских особей). Например, кариотип человека содержит 22 пары аутосом и две половые хромосомы – ХХ у женщины и XY y мужчины (44,ХУ и 44,XYсоответственно). Соматические клетки организмов содержат диплоидный (двойной) набор хромосом, а гаметы – гаплоидный (одинарный).
Идиограмма – это систематизированный кариотип, в котором хромосомы располагаются по мере уменьшения их размеров. Точно расположить хромосомы по размеру удается далеко не всегда, так как некоторые пары имеют близкие размеры. Поэтому в I960 г. была предложена Денверская классификация хромосом, которая помимо их размеров учитывает форму, положение центромеры, наличие вторичных перетяжек и спутников (рис. 3). Согласно этой классификации, 23 пары хромосом человека разбили на 7 групп – от А до G. Важным признаком, облегчающим классификацию, являетсяцентромерный индекс (ЦИ), который отражает отношение (в процентах) длины короткого плеча к длине всей хромосомы.
Различают следующие группы хромосом:
Группа В (хромосомы 4 и 5). Это большие субметацентрические хромосомы, их центромерный индекс 24-30. Они не различаются между собой при обычном окрашивании.
Группа С (хромосомы 6-12). Хромосомы среднего размера, субметацентрические, их центромерный индекс 27-35. В 9-й хромосоме часто обнаруживается вторичная перетяжка. К этой группе относят и Х-хромосому.
Группа D (хромосомы 13-15). Хромосомы акроцентрические, сильно отличаются от всех других хромосом человека, их центромерный индекс около 15. Все три пары имеют спутники.
Группа Е (хромосомы 16-18). Хромосомы относительно короткие, метацентрические или субметацентрические, их центромерный индекс от 26 до 40. В длинном плече 16-й хромосомы в 10% случаев выявляется вторичная перетяжка.
Группа F (хромосомы 19 и 20). Хромосомы короткие, субметацентрические, их центромерный индекс 36-46.
Группа G (хромосомы 21 и 22). Хромосомы маленькие, акроцентрические, их центромерный индекс 13-33. К этой группе относят и У-хромосому.
Рис. 3. Денверская классификация хромосом человека
В основе Парижской классификации хромосом человека (1971 г.) лежат методы специального дифференциального их окрашивания, при которых в каждой хромосоме выявляется характерный только для нее порядок чередования поперечных светлых и темных сегментов (рис. 4). Различные типы сегментов обозначают по методам, с помощью которых они выявляются наиболее четко. Данные методы позволяют четко дифференцировать хромосомы человека внутри групп.
Короткое плечо хромосом обозначают латинской буквой р, а длинное – q. Каждое плечо хромосомы разделяют на районы, нумеруемые от центромеры к теломерам. В некоторых коротких плечах выделяют один такой район, а в других (длинных) – до четырех. Полосы внутри районов нумеруются по порядку от центромеры. Локализация генов не всегда известна с точностью до полосы. Так, местоположение гена ретинобластомы обозначают 13q, что означает локализацию его в длинном плече тринадцатой хромосомы.
Основные функции хромосом состоят в хранении, воспроизведении и передаче генетической информации при размножении клеток и организмов.
Читайте также: