
Мост функции кратко рефлее ирая
Обновлено: 02.07.2024
Жевательный рефлекс проявляется сокращениями и расслаблениями жевательных мышц в ответ на поступление афферентных сигналов от сенсорных рецепторов внутренней части губ и полости рта через волокна тройничного нерва к нейронам ядра тройничного нерва. Эфферентные сигналы к жевательным мышцам передаются через двигательные волокна лицевого нерва.
Роговичный рефлекс проявляется закрытием век обоих глаз (морганием) в ответ на раздражение роговицы одного из глаз. Афферентные сигналы от сенсорных рецепторов роговицы передаются по сенсорным волокнам тройничного нерва к нейронам ядра тройничного нерва. Эфферентные сигналы к мышцам век и круговой мышце глаза передаются через двигательные волокна лицевого нерва.
Слюноотделительный рефлекс проявляется отделением большего количества жидкой слюны в ответ на раздражение рецепторов слизистой полости рта. Афферентные сигналы от рецепторов слизистой полости рта передаются по афферентным волокнам тройничного нерва к нейронам его верхнего слюноотделительного ядра. Эфферентные сигналы предаются от нейронов этого ядра к эпителиальным клеткам слюнных желез через языкоглоточный нерв.
Слезоотделительный рефлекс проявляется усиленным слезовыделением в ответ на раздражение роговицы глаза. Афферентные сигналы передаются по афферентным волокнам тройничного нерва к нейронам верхнего слюноотделительного ядра. Эфферентные сигналы к слезным железам передаются через волокна лицевого нерва.
Глотательный рефлекс проявляется осуществлением координированного сокращения мышц, обеспечивающих глотание при раздражении рецепторов корня языка, мягкого неба и задней стенки глотки. Афферентные сигналы передаются по афферентным волокнам тройничного нерва к нейронам моторного ядра и далее к нейронам других ядер ствола мозга. Эфферентные сигналы от нейронов ядер тройничного, подъязычного, языкоглоточного и блуждающего нервов передаются к иннервируемым ими мышцам языка, мягкого нёба, глотки, гортани и пищевода.
Координация работы жевательных и других мышц
Жевательная мускулатура может развивать высокую степень напряжения. Мышца с поперечным сечением в 1 см 2 при сокращении развивает силу в 10 кг. Сумма поперечного сечения жевательных мышц, поднимающих нижнюю челюсть на одной стороне лица, равна в среднем 19,5 см 2 , а на обеих сторонах — 39 см 2 ; абсолютная сила жевательных мышц составляет 39 х 10 = 390 кг.
Жевательные мышцы обеспечивают смыкание челюстей и поддерживают закрытое состояние рта, не требующие развития в мышцах существенного напряжения. В то же время при пережевывании грубой пищи или усиленном смыкании челюстей жевательные мышцы способны развивать предельные напряжения, превышающие выносливость пародонта отдельных зубов к оказываемому на них давлению и вызывающие болевые ощущения.
Из приведенных примеров очевидно, что у человека должны существовать механизмы, с помощью которых поддерживается тонус жевательных мышц в покое, инициируются и координируются сокращения и расслабления различных мышц во время жевания. Эти механизмы необходимы для достижения эффективности жевания и предотвращения развития избыточного напряжения мышц, которое могло бы привести к возникновению болевых ощущений и других неблагоприятных последствий.
Жевательные мышцы относятся к поперечно-полосатой мускулатуре, поэтому им присущи те же свойства, что и другим исчерченным скелетным мышцам. Их сарколемма обладает возбудимостью и способностью проводить потенциалы действия, возникающие при возбуждении, а сократительный аппарат обеспечивает сокращение мышц вслед за их возбуждением. Жевательные мышцы иннервируются аксонами а-мотонейронов, формирующими двигательные порции: нижнечелюстного нерва — ветви тройничного нерва (жевательные, височные мышцы, переднее брюшко двубрюшной и челюстно-подъязычная мышцы) и лицевого нерва (вспомогательные — шилоподъязычная и двубрюшная мышцы). Между окончаниями аксонов и сарколеммой волокон жевательных мышц имеются типичные нервно-мышечные синапсы, передача сигналов в которых осуществляется с помощью ацетилхолина, взаимодействующего с н-холинорецегггорами постсинаптических мембран. Таким образом, для поддержания тонуса, инициирования сокращения жевательных мышц и регуляции его силы используются те же принципы, что и в других скелетных мышцах.
Удерживание в покос закрытого состояния рта достигается за счет наличия тонического напряжения в жевательной и височной мышцах, которое поддерживается рефлекторными механизмами. Под действием се массы нижняя челюсть постоянно растягивает рецепторы мышечных веретен. В ответ на растяжение в окончаниях нервных волокон, ассоциированных с этими рецепторами, возникают афферентные нервные импульсы, передающиеся по чувствительной порции волокон тройничного нерва к нейронам мезэнцефалического ядра тройничного нерва и поддерживают активность моторных нейронов. Последние постоянно посылают поток эфферентных нервных импульсов к экстрафузальным волокнам жевательных мышц, создающим напряжение достаточной силы для удерживания закрытого состояния рта. Активность моторных нейронов ядра тройничного нерва может подавляться под влиянием тормозных сигналов, посылаемых по кортикобульбарным путям из области нижней части первичной моторной коры. Это сопровождается снижением потока эфферентных нервных импульсов к жевательным мышцам, их расслаблением и открытием рта, имеющим место при произвольном открытии рта, а также во время сна или наркоза.
Жевательные и другие движения нижней челюсти осуществляются с участием жевательных, мимических мышц, языка, губ и других вспомогательных мышц, иннервируемых различными черепными нервами. Они могут быть произвольными и рефлекторными. Жевание может быть эффективным и достигать своей цели при условии тонкой координации сокращения и расслабления участвующих в нем мышц. Функцию координации выполняет центр жевания, представленный сетью сенсорных, моторных и интернейронов, расположенных преимущественно в стволе мозга, а также в черной субстанции, таламусе и коре большого мозга.
Информация, поступающая в структуры жевательного центра от вкусовых, обонятельных, термо-, механо- и других сенсорных рецепторов, обеспечивает формирование ощущений имеющейся или поступившей в полость рта пищи. Если параметры ощущений о поступившей пище не соответствуют ожидаемым, то в зависимости от мотивации и чувства голода может развиться реакция отказа от ее приема. При совпадении параметров ощущений с ожидаемыми (извлеченными из аппарата памяти) в центре жевания и других двигательных центрах мозга формируется моторная программа предстоящих действий. В результате реализации моторной программы обеспечивается придание телу определенной позы, осуществление, согласованное с движением рук, открывание и закрывание рта, откусывание и введение пиши в рот, вслед за которыми инициируются произвольные и рефлекторные компоненты жевания.
Предполагается, что в нейронных сетях центра жевания имеется сформированный в процессе эволюции генератор двигательных команд, посылаемых к моторным нейронам ядер тройничного, лицевого, подъязычного черепных нервов, иннервирующих жевательные и вспомогательные мышцы, а также к нейронам моторных центров ствола и спинного мозга, инициирующих и координирующих движения рук, откусывание, пережевывание и проглатывание пищи.
Жевательные и другие движения приспосабливаются к консистенции и другим особенностям пищи. Ведущую роль в этом играют сенсорные сигналы, посылаемые в центр жевания и непосредственно к нейронам ядра тройничного нерва по волокнам мезэнцефалического тракта и, в частности, сигналы от проприорецепторов жевательных мышц и механорецепторов периодонта. Результаты анализа этих сигналов используются для рефлекторной регуляции жевательных движений.
При усиленном смыкании челюстей происходит избыточная деформация пародонта и механическое раздражение рецепторов, расположенных в периодонте и (или) в десне. Это приводит к рефлекторному ослаблению давления за счет снижения силы сокращения жевательных мышц. Существует несколько рефлексов, с помощью которых жевание тонко приспосабливается к характеру принимаемой пищи.
Массетерный рефлекс инициируется сигналами проприорецепторов основных жевательных мышц (особенно m. masseter), ведущими к повышению тонуса чувствительных нейронов, активации а-моторных нейронов мезэнцефалического ядра тройничного нерва, иннервирующих мышцы, поднимающие нижнюю челюсть. Активация моторных нейронов, повышение частоты и количества эфферентных нервных импульсов в двигательных нервных волокнах тройничных нервов способствуют синхронизации сокращения двигательных единиц, вовлечению в сокращение высокопороговых моторных единиц. Это ведет к развитию сильных фазических сокращений жевательных мышц, обеспечивающих подъем нижней челюсти, смыкание зубных дуг и повышение жевательного давления.
Пародонтомышечные рефлексы обеспечивают контроль за силой жевательного давления на зубы во время сокращений мышц, поднимающих нижнюю челюсть, и сжатии челюстей. Они возникают при раздражении механорецепторов пародонта, чувствительных к изменению жевательного давления. Рецепторы расположены в связочном аппарате зуба (периодонте), а также в слизистой оболочке десны и альвеолярных гребней. Соответственно выделяют два вида пародонтомышечных рефлексов: периодонтомышечный и о гингивомышечный.
Периодомтомышечный рефлекс защищает периодонт от избыточного давления. Рефлекс осуществляется во время жевания при помощи собственных зубов в ответ на раздражение механорецептров периодонта. Выраженность рефлекса зависит от силы давления и чувствительности рецепторов. Афферентные нервные импульсы, возникшие в рецепторах при их механическом раздражении высоким жевательным давлением, развиваемым при пережевывании твердой пищи, передаются по афферентным волокнам чувствительных нейронов гассерова ганглия к нейронам чувствительных ядер продолговатого мозга, затем — в таламус и кору большого мозга. От корковых нейронов эфферентная импульсация по коргико- бульбарному пути поступает в жевательный центр, моторные ядра, где вызывает активацию а-мотонейронов, иннервирующих вспомогательные жевательные мышцы (опускающие нижнюю челюсть). Одновременно активируются тормозные интернейроны, снижающие активность а-моторных нейронов, иннервирующих основные жевательные мышцы. Это приводит к уменьшению силы их сокращений и жевательного давления на зубы. При надкусывании пищи с очень твердым компонентом (например, скорлупы орехов или косточек) могут возникать болевые ощущения и остановка акта жевания для удаления твердого тела из полости рта во внешнюю среду или его перемещения на зубы с более устойчивым пародонтом.
Гингивомышечный рефлекс осуществляется в процессе сосания и (или) жевания у новорожденных или у пожилых людей после потери зубов, когда сила сокращений основных жевательных мышц контролируется механорецепторами слизистой оболочки десны и альвеолярных гребней. Особое значение этот рефлекс приобретает у людей, пользующихся съемными протезами (при частичной или полной адентии), когда передача жевательного давления осуществляется непосредственно на слизистую оболочку десны.
Важное значение в регуляции сокращения основных и вспомогательных жевательных мышц имеет артикуляционно-мышечный рефлекс, возникающий при раздражении механо-рецепторов, расположенных в капсуле и связках височно-нижнечелюстных суставов.
Мост располагается выше продолговатого мозга и выполняет сенсорные, проводниковые, двигательные, интегративные рефлекторные функции.
В состав моста входят ядра лицевого, тройничного, отводящего, преддверно-улиткового нерва (вестибулярные и улитковые ядра), ядра преддверной части преддверно-улиткового нерва (вестибулярного нерва): латеральное (Дейтерса) и верхнее (Бехтерева). Ретикулярная формация моста тесно связана с ретикулярной формацией среднего и продолговатого мозга.
Важной структурой моста является средняя ножка мозжечка. Именно она обеспечивает функциональные компенсаторные и морфологические связи коры большого мозга с полушариями мозжечка.
Сенсорные функции моста обеспечиваются ядрами преддверно-улиткового, тройничного нервов. Улитковая часть преддверно-улиткового нерва заканчивается в мозге в улитковых ядрах; преддверная часть преддверно-улиткового нерва — в треугольном ядре, ядре Дейтерса, ядре Бехтерева. Здесь происходит первичный анализ вестибулярных раздражений их силы и направленности.
Чувствительное ядро тройничного нерва получает сигналы от рецепторов кожи лица, передних отделов волосистой части головы, слизистой оболочки носа и рта, зубов и конъюнктивы глазного яблока. Лицевой нерв иннервирует все мимические мышцы лица. Отводящий нерв иннервирует прямую латеральную мышцу, отводящую глазное яблоко кнаружи.
Двигательная порция ядра тройничного нерва иннервирует жевательные мышцы, мышцу, натягивающую барабанную перепонку, и мышцу, натягивающую небную занавеску.
Проводящая функция моста. Обеспечивается продольно и поперечно расположенными волокнами. Поперечно расположенные волокна образуют верхний и нижний слои, а между ними проходят идущие из коры большого мозга пирамидные пути. Между поперечными волокнами расположены нейронные скопления — ядра моста. От их нейронов начинаются поперечные волокна, которые идут на противоположную сторону моста, образуя среднюю ножку мозжечка и заканчиваясь в его коре.
В покрышке моста располагаются продольно идущие пучки волокон медиальной петли. Они пересекаются поперечно идущими волокнами трапециевидного тела, представляющие собой аксоны улитковой части преддверно-улиткового нерва противоположной стороны, которые заканчиваются в ядре верхней оливы. От этого ядра идут пути боковой петли, которые направляются в заднее четверохолмие среднего мозга и в медиальные коленчатые тела промежуточного мозга.
В покрышке мозга локализуются переднее и заднее ядра трапециевидного тела и латеральной петли. Эти ядра вместе с верхней оливой обеспечивают первичный анализ информации от органа слуха и затем передают информацию в задние бугры четверохолмий.
В покрышке также расположены длинный медиальный и тектоспинальный пути.
Собственные нейроны структуры моста образуют его ретикулярную формацию, ядра лицевого, отводящего нервов, двигательной порции ядра и среднее сенсорное ядро тройничного нерва.
Ретикулярная формация моста является продолжением ретикулярной формации продолговатого мозга и началом этой же системы среднего мозга. Аксоны нейронов ретикулярной формации моста идут в мозжечок, в спинной мозг (ретикулоспинальный путь). Последние активируют нейроны спинного мозга.
Ретикулярная формация моста влияет на кору большого мозга, вызывая ее пробуждение или сонное состояние. В ретикулярной формации моста находятся две группы ядер, которые относятся к общему дыхательному центру. Один центр активирует центр вдоха продолговатого мозга, другой — центр выдоха. Нейроны дыхательного центра, расположенные в мосте, адаптируют работу дыхательных клеток продолговатого мозга в соответствии с меняющимся состоянием организма.
В основе деятельности нервной системы лежат рефлексы (лат. reflexus — отражённый). Рефлекс - ответная реакция организма на действие раздражителя.
Любой рефлекс существует на базе рефлекторной дуги - совокупности соединенных друг с другом нервных элементов, через которые последовательно проводится нервный импульс при осуществлении рефлекса. Самый прострой пример - коленный рефлекс, который часто проверяет невролог, что позволяет быстро сделать вывод о сохранности элементов рефлекторной дуги.
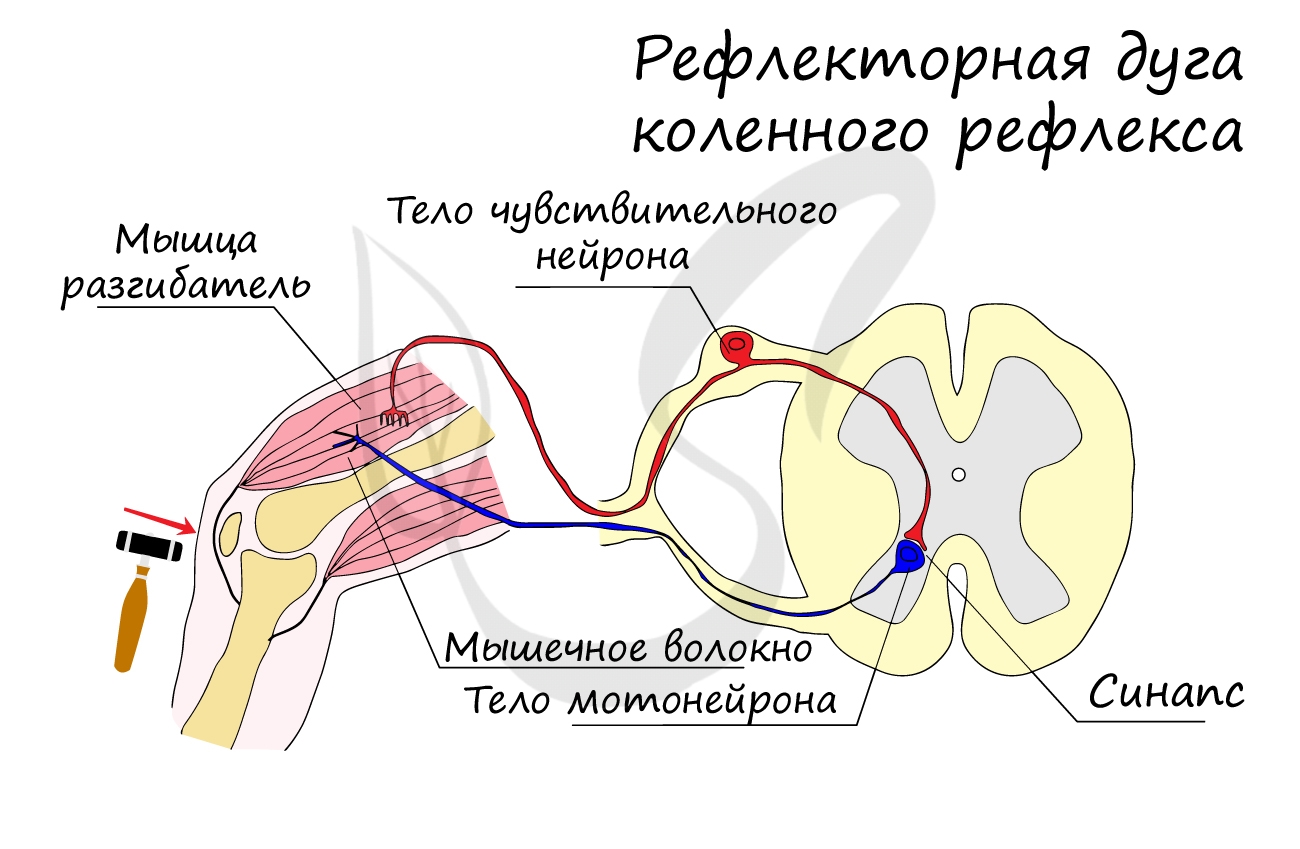
Нейроны соединяются друг с другом с помощью отростков: аксонов и дендритов, на конце которых находятся специальные контакты - синапсы, которые мы подробно изучили в статье про нервные ткани.
Устройство рефлекторных дуг
-
Чувствительное (афферентное, центростремительное)
Состоит из рецептора (может быть расположен в коже, внутренних органах, сосудах) чувствительного нейрона и идущего от этого нейрона чувствительного волокна, которое проникает в спинной мозг через задние рога.
Тело чувствительного нейрона находится в задних корешках (!) спинного мозга. Представили? А теперь представьте дендрит, идущий от кончика вашего указательного пальца до самого спинного мозга. Именно поэтому неверно считать, что дендрит - всегда "короткий" отросток, а аксон - "длинный". Данный вопрос мы обсуждали в статье про нервные ткани.
Состоит из вставочного нейрона и его отростков. Вставочный нейрон осуществляет связь между чувствительным и двигательным звеном рефлекторной дуги. Вставочные нейроны могут осуществлять связь с другими отделами ЦНС.
Тела вставочных нейронов находятся в задних рогах спинного мозга.
Представлено двигательным нейроном (эфферентным, исполнительным, мотонейроном), от которого нервные волокна идут к рабочему органу (эффектору, органу-исполнителю).
В зависимости от того, чем представлен эффектор - мышца, железа - при поступлении к нему нервных импульсов его работа активируется: мышца начинает сокращаться, железа - выделять секрет.
Двигательные нейроны лежат в передних рогах спинного мозга, откуда и выходят их отростки.
Рассмотрим схему рефлекторной дуги, на базе которой осуществляется рефлекс отдергивания руки от горячего предмета. Попробуйте сами описать путь, который проходит нервный импульс и вспомнить 3 звена рефлекторной дуги. Назовите локализацию каждого из нейронов.
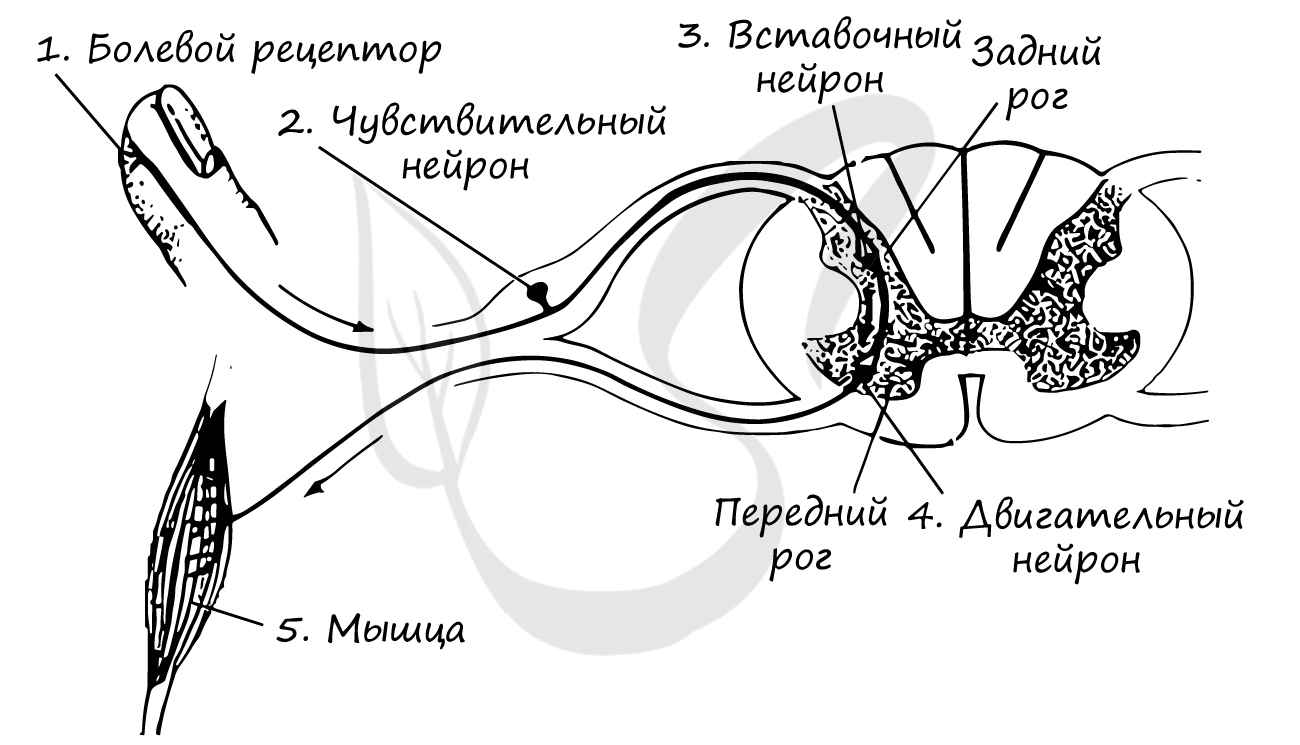
Это может показаться очевидным, но необходимо подчеркнуть, что афферентные нервные волокна входят в спинной мозг через задние корешки. Эфферентные нервные волокна выходят из спинного мозга через передние корешки.
Виды рефлекторных дуг
Рефлекторные дуги подразделяются на соматические и вегетативные. С помощью соматических рефлекторных дуг осуществляются рефлексы, обеспечивающие возможность произвольных движений (совершаемых по воле человека). С помощью вегетативных - координация деятельности внутренних органов, то есть функции, которые не поддаются нашему осознанному контролю (вспомните вегетативную нервную систему).
Ниже вы увидите схемы соматической и вегетативной рефлекторных дуг. Под картинкой будет написано существенное отличие между ними, которое вы должны запомнить, но прежде постарайтесь сами сделать вывод, изучив картинку.
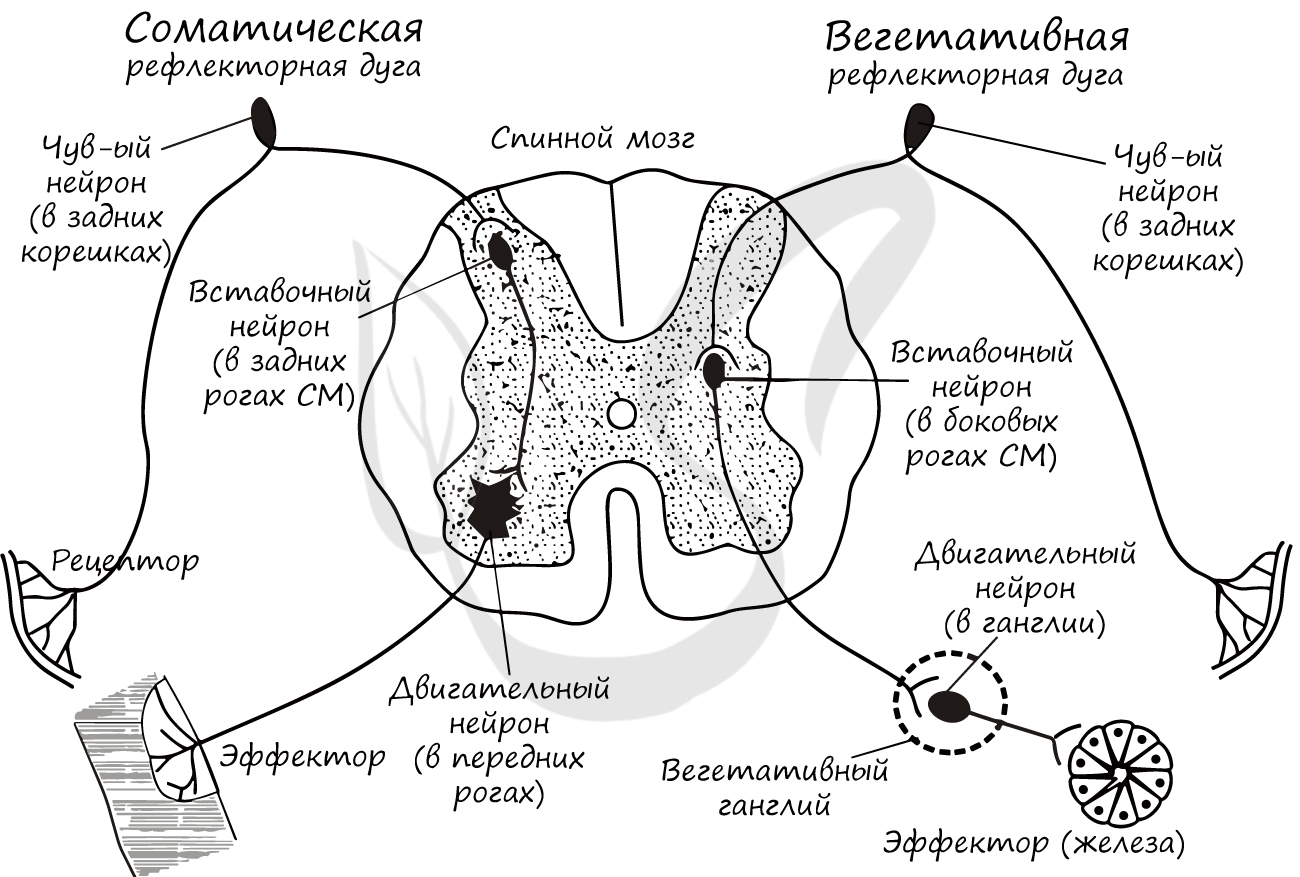
Отличием между соматической и вегетативной рефлекторными дугами в том, что в составе последней эфферентный нейрон лежит за пределами спинного мозга - в вегетативном ганглии. Данные ганглии могут располагаться по бокам от позвоночника, вблизи внутренних органов или в их стенке.
Также вы, скорее всего, обратили внимание, что вставочный нейрон вегетативной дуги локализован в другом месте - в боковых рогах спинного мозга (а не в задних, как в соматической).
Нервная регуляция
Рефлекторная дуга - фундамент, на котором осуществляется рефлекс. В нервной системе возникают не только процессы возбуждения, но и торможения, о которых мы подробнее поговорим в теме, посвященной высшей нервной деятельности. Торможение заключается в ослаблении или задержке уже возникшего возбуждения.
Таким образом, координация и регулирования процессов возбуждения и торможения - основа согласованной работы органов и систем органов, составляющих единый организм.
Заболевания
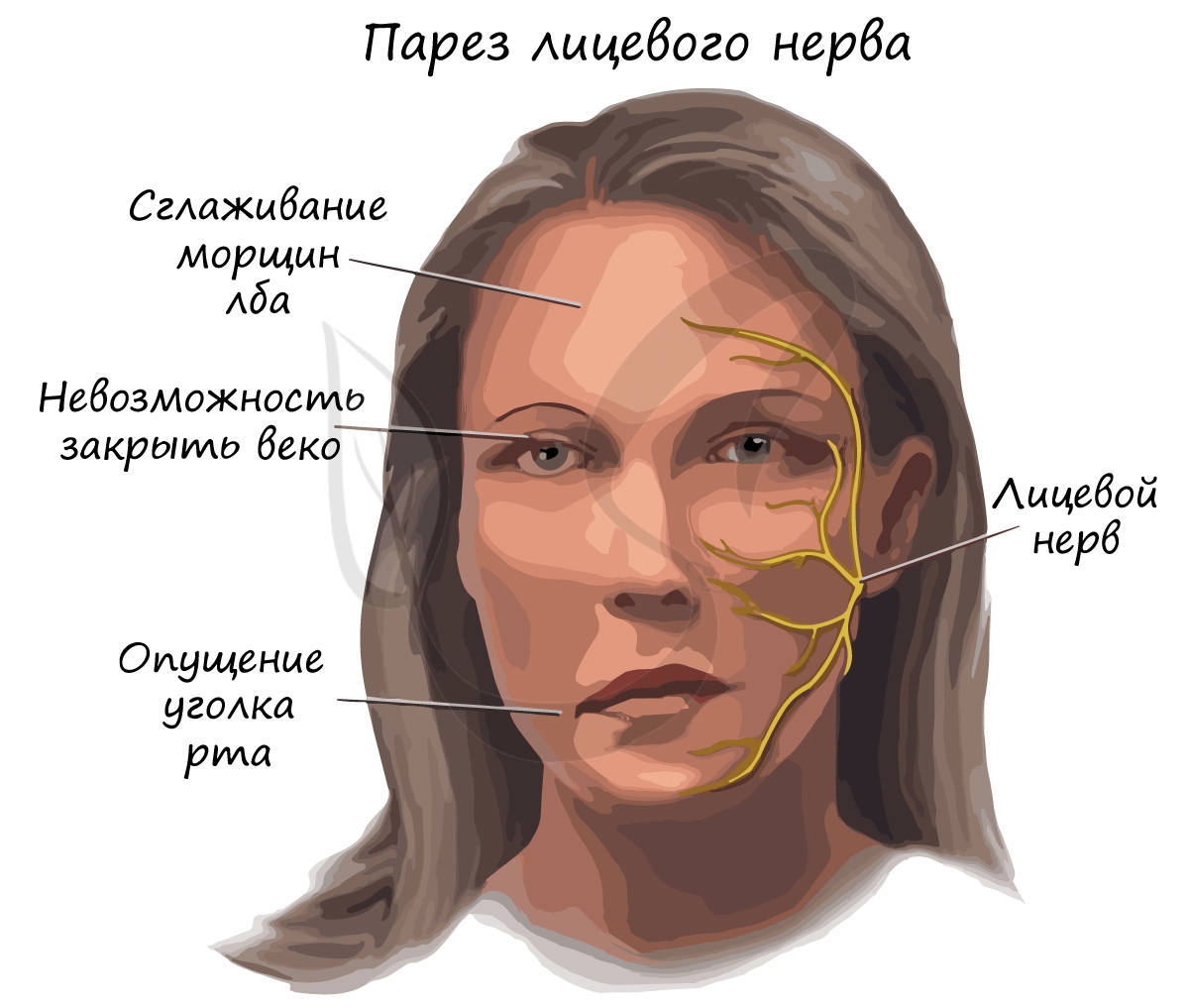
Парез (греч. πάρεσις - ослабление) - неврологический синдром, обусловленный поражением двигательного (эфферентного) пути и слабостью в конечности, или в другом органе, который данный нервный путь иннервировал. Парез проявляется снижением мышечной силы, движения в неполном объеме сохраняются.
Паралич (греч. παράλυσις - расслабление) - полное отсутствие произвольных движений, обусловленное теми же причинами, что и парез.
При переохлаждении может возникнуть парез лицевого нерва. Причиной этому служит воспаление тканей, в результате чего в узком костном канале нерв сдавливается воспаленными тканями. Нервные импульсы частично, либо полностью перестают поступать к мышцам лица, что делает невозможным для пациента движение ими.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.
Задний мозг - Ядро среднемозгового пути тройничного нерва
ЗАДНИЙ МОЗГ
, pons. Расположен между межножковой ямкой и пирамидами. Содержит, главным образом, проекционные нервные волокна и группы нейронов. Рис. А, Рис. Б.
Бульбарно-мостовая борозда
, sulcus bulbopontinus. Лежит между продолговатым мозгом и нижним краем моста. Место выхода VI, VII, VII нервов. Рис. А.
Базилярная борозда
, sulcus basilaris. Идет по средней линии вентральной поверхности моста между возвышениями, соответствующими пирамидным проводящим путям, содержит одноименную артерию. Рис. А, Рис. В.
Средняя мозжечковая ножка
, pedunculus cerebellaris medius (pontinus). Соединяет мост с мозжечком. Содержит волокна мостомозжечкового пути. Рис. А.
Мостомозжечковый треугольник
, trigonum pontocerebellarе. Важная с клинической точки зрения область, расположенная между мостом, продолговатым мозгом и мозжечком. Рис. А.
РАЗРЕЗЫ МОСТА
Передняя (базилярная) часть моста
, pars anterior (basilaris) pontis. Содержит волокна корково-мостового и мостомозжечкового путей. Рис. В.
Продольные волокна моста
, fibrae pontis longitudinales. Состоят из описанных ниже групп волокон, идущих в продольном направлении.
Корково-спинномозговые волокна
Корково-ядерные волокна
Корково-ретикулярные волокна
, fibrae corticoreticulares. Направляются от коры полушарий большого мозга к нейронам ретикулярной формации.
Корково-мостовые волокна
Поперечные волокна моста
Мостомозжечковые волокна
Ядра моста
, nuclei pontis. Расположены в вентральной части моста. Отростки их нейронов направляются к мозжечку. Рис. В.
Задняя часть моста (покрышка моста)
, pars posterior pontis (tegmentum pontis). Находится между четвертым желудочком и трапециевидным телом. Рис. В.
Шов моста
, raphe pontis. Расположен по средней линии моста. Образован волокнами мостового ядра тройничного нерва. Рис. В.
Медиальный продольный пучок
, fasciculus longitudinalis medialis. Соединяют двигательные ядра III, IV, VI, XI и вестибулярные ядра VIII черепных нервов с мотонейронами передних рогов спинного мозга, иннервирующими мышцы шеи. Рис. В.
Задний продольный [[Шютца]] пучок
, fasciculus longitudinalis dorsalis [[Schutz]]. Соединяет между собой гипоталамус, добавочное, верхнее и нижнее слюноотделительные, двойное ядра, заднее ядро блуждающего нерва, ядро одиночного пути, ядра лицевого и подъязычного нервов. Рис. В.
Медиальная петля
, lemniscus medialis. Продолжение перекреста медиальных петель. Соединяет тонкое и клиновидное ядра с ядрами таламуса. Рис. В.
Покрышечноспинномозговой путь
, tractus tectospinalis. Начинается в верхнем холмике, переходит на противоположную сторону и располагается вентрально от медиального продольного пучка. Рис. В.
Ретикулярная формация
, formatio reticularis. Расположена в задней части моста. Состоит из отдельных клеточных групп, которые пронизаны нервными волокнами, идущими в различных направлениях. Осуществляет интеграцию висцеральных функций с сокращением произвольной мускулатуры. Рис. В.
Спинномозговая петля
, lemniscus spinalis. Продолжение в краниальном направлении переднего и латерального спинно-таламических путей. Рис. В.
Спинномозговой путь тройничного нерва
, tractus spinalis nervi trigeminalis. Образован отростками нейронов узла V нерва, которые после вступления в вещество мозга спускаются вниз и заканчиваются на клетках одноименного ядра тройничного нерва. Рис. Б.
Ядро (нижнее) спинномозгового пути тройничного нерва
Мостовое ядро тройничного нерва
, nucleus pontinus nervi trigeminalis. Основное ядро V нерва, на нейронах которого заканчиваются волокна проводящие, в основном, тактильную чувствительность. Рис. Б.
Тройничная петля (тройнично-таламический путь)
Среднемозговой путь тройничного нерва
, tractus mesencephalicus nervi trigeminalis (tractus mesencephalicus trigeminalis). Образован отростками нейронов одноименного ядра и проходит в латеральной части покрышки моста. Проводник проприоцептивной чувствительности от жевательных и мимических мышц. Рис. Б, Рис. В.
Ядро среднемозгового пути тройничного нерва
, nucleus mesencephalicus nervi trigeminalis [nucleus mesencephalicus trigeminalis].Верхнее чувствительное ядро V черепного нерва в покрышке ножек мозга. Рис. Б.
Спокойное дыхание взрослого человека характеризуется последовательной сменой актов вдоха и выдоха с частотой 14-16 дыханий в минуту.
Чтобы произошел вдох, необходимо сокращение дыхательных мышц.
Импульсы к ним поступают от мотонейронов передних рогов спинного мозга.
К диафрагме – от III – IV шейных сегментов, к межреберным мышцам – от грудных сегментов спинного мозга.
Мотонейроны получают импульсы от дыхательного центра , расположенного в продолговатом мозге.
Если перерезать спинной мозг под продолговатым мозгом , то дыхание – прекращается .
Дыхательный центр
Был открыт в 1812 г. Легаллуа и в 1842 г. Флурансом, которые своими опытами доказали его локализацию в продолговатом мозге .
Н.А.Миславский в 1885 г. уточнил местоположение дыхательного центра – в области РФ (ретикулярной формации) продолговатого мозга , т.к. перерезка между грудными и шейными сегментами спинного мозга – сохраняет диафрагмальное дыхание:
- продолговатым и спинным – полностью прекращает дыхание ,
- выше продолговатого мозга – дыхание сохраняется .
Н.А.Миславским также было показано, что дыхательный центр состоит из двух отделов:
- инспираторного (отвечает на вдох),
- экспираторного (отвечает за выдох).
Они находятся в реципрокных (то есть в противоположных) отношениях.
В настоящее время установлено:
- в РФ продолговатого мозга находятся инспираторные и экспираторные нейроны ,
- эти нейроны рассеяны диффузно , среди других нейронов РФ и относительно друг друга,
- инспираторных нейронов примерно вдвое больше , чем экспираторных.
В продолговатом мозге есть еще два скопления дыхательных нейронов – дорсальные и вентральные дыхательные ядра.
Дорсальное ядро содержит, в основном, инспираторные нейроны, контролирующие сокращение диафрагмы .
В вентральном ядре содержатся как инспираторные , так и экспираторные нейроны, которые включаются при форсированном дыхании.
Все дыхательные нейроны делят на 6 групп:
- ранние инспираторные – активны в начале фазы вдоха (инспирации),
- поздние инспираторные – активны в конце вдоха,
- полные инспираторные – активны в течение всего вдоха,
- постинспираторные – максимальный разряд в начале выдоха,
- экспираторные – активны во вторую фазу выдоха,
- преинспираторные – активны перед вдохом, они включают активную экспирацию (выдох.
Значение дыхательных нейронов:
- Преинспираторные и ранние инспираторные влияют на момент начала вдоха.
- Инспираторные нейроны ДЦ генерируют дыхательный ритм (частоту и глубину дыхания), иннервируют мотонейроны.
- Постинспираторные нейроны контролируют процесс пассивного выдоха.
- Экспираторные нейроны отвечают за активный выдох, так как иннервируют мотонейроны внутренних межреберных мышц и мышц передней брюшной стенки.
Дыхательный центр – это совокупность нервных клеток, расположенных в различных отделах ЦНС и принимающих участие в регуляции дыхания .
Опыт Фредерика (1901 г.) с перекрестным кровообращением у собак
У двух собак, находящихся под наркозом, перекрестно соединяли сонные артерии и яремные вены, латерально же расположенные сосуды пережимали. При этом голова 1-й собаки снабжались кровью из туловища 2-й и наоборот.
У первой собаки кратковременно пережимали трахею, и у нее в крови уменьшалось содержание кислорода (гипоксемия) и увеличивалось содержание углекислого газа (гиперкапния). Эта кровь поступала в голову 2-й собаки, и у нее наступала одышка (диспноэ).
В результате у нее в крови увеличивалось содержание кислорода (гипероксимия) и уменьшалось содержание углекислого газа (гипокапния), и эта кровь поступала в голову 1-й собаки, и у нее наступало апноэ – остановка дыхания. (Нормальное дыхание — эйпноэ).
На состояние ДЦ влияет газовый состав крови:
При увеличении в крови напряжения углекислого газа и уменьшении кислорода, ДЦ – возбуждается и, наоборот, уменьшается , если в крови уменьшается напряжение углекислого газа и увеличивается напряжение кислорода.
Опыт Холдена
При дыхании в герметичной камере в воздухе увеличивалось содержание CO2 и уменьшалось O2 – наступала одышка . Когда CO2 поглощается натронной известью, одышка наступала намного позже, хотя содержание O2 в воздухе значительно снижалось .
Одышка наступает:
- При снижении O2 в атмосферном воздухе с 20,94% до 12%, т.е. на 9%.
- При повышении содержания CO2 в альвеолярном воздухе на 0,17% вентиляция удваивается.
Главный активатор ДЦ – CO2
Действие карбогена
Карбоген – газовая смесь, состоящая из 96% — O2, 4% — CO2.
В сравнении с воздухом в карбогене в 4,8 раза больше O2 и в 130 раз – CO2.
Карбоген применяют при расстройствах дыхания.
Эффект карбогена связан с эффектами содержащегося в нем CO2:
- стимуляция ДЦ,
- расширение бронхов и кровеносных сосудов,
- сдвиг кривой диссоциации HbO2 вправо –> увеличение диффузии O2 из крови в ткани.
Карбоген применяется в медицинских целях для лечения горной болезни, отравления угарным газом, глаукомы, стресса, при восстановлении слуха после воздействия шума и в ряде случаев для улучшения кровоснабжения опухолей при химио- и лекарственной терапии.
В 1911 г. Винтерштейн доказал, что возбудителем ДЦ также являются ионы Н .
Ацидоз – усиливает легочную вентиляцию.
Он показал, что возбуждают ДЦ нелетучие кислоты – молочная, никотиновая и другие.
Самым сильным стимулятором дыхания являются:
Механизм действия гуморальных факторов (CO2, O2, H) на ДЦ :
- Через хеморецепторы (периферические) сосудистых рефлексогенных зон.
- Через хеморецепторы, находящиеся в продолговатом мозге (медуллярные).
Периферические хеморецепторы:
- расположены в каротидных и в аортальных тельцах,
- реагируют на (в артериальной крови):
- увеличение pCO2,
- уменьшение pO2,
- увеличение H (от есть уменьшение pH).
Хеморецепторы возбуждаются постоянно CO2 и O2 , растворенными в крови, а также H, т.к. порог для pCO2 равен 20-30 мм.рт.ст. В норме pCO2 = 40 мм.рт.ст.
Порог для pO2 равен 130-140 мм.рт.ст. В норме pO2 = 100 мм.рт.ст. Одышка же наступает при pO2 ниже 50-60 мм.рт.ст.
Таким образом, хеморецепторы постоянно посылают импульсы в ДЦ, возбуждая инспираторные нейроны, причем большую роль играют хеморецепторы каротидного синуса.
Медуллярные хеморецепторы:
- Находятся на вентролатеральной поверхности продолговатого мозга .
- Реагируют только на H и изменение напряжения CO2.
- Эти рецепторы возбуждаются позднее, поскольку требуется время для проникновения CO2 через гематоэнцефалический барьер.
- Импульсы, поступающие с медуллярных хеморецепторов в ДЦ, увеличивают прирост вентиляции на 60-80% .
Механорецепторы (рецепторы растяжения)
Находятся в:
- легких,
- дыхательных путях,
- дыхательных мышцах (проприорецепторы)
Это барорецепторы рефлексогенных зон.
Механорецепторы легких (РРЛ)
Рецепторы растяжения легких (РРЛ) – являются наиболее значимым среди всех механорецепторов.
В 1868 г. Геринг и Брейер доказали наличие в легких рецепторов, которые возбуждаются при их растяжении, то есть при вдохе. Они являются чувствительными окончаниями блуждающих нервов, которые направляют свои импульсы в ДЦ.
Геринг и Брейтер раздували легкие и наблюдали прекражение вдоха (инспираторно-тормозной рефлекс).
Таким образом, этот рефлекс способствует смене вдоха на выдох. Он называется рефлексом Геринга-Брейера и является рефлексом саморегуляции дыхания.
При перерезке блуждающих нервов, дыхание становится редким и глубоким, альвеолы расширяются до максимального предела, т.к. вдох не тормозится. В этом случае, смене вдоха на выдох будет способствовать пневмотаксический центр (ПТЦ).
В настоящее время известно, что в легких существует 3 разновидности механорецепторов :
- РРЛ медленноадаптирующиеся,
- РРЛ быстроадаптирующиеся или ирритантные,
- Юкстаальвеолярные рецепторы капилляров.
РРЛ медленноадаптирующиеся:
- Они расположены в ГМК (гладкомышечных клетках) дыхательных путей.
- Возбуждаются при вдохе.
- С них осуществляется рефлекс Геринга-Брейера.
Ирритантные рецепторы (быстроадаптирующиеся):
- Расположены в слизистой дыхательных путей.
- Реагируют на механические и химические стимулы.
- Быстро адаптирующиеся.
- Длительное раздражение этих рецепторов приводит к хроническому бронхиту.
Физиологическое значение при вдыхании токсических веществ: Сужение бронхов -> вентиляция альвеол -> поступление этих веществ в альвеолы и кровь.
- расположены в паренхиме легких, в альвеолярных перегородках, прилегающих к капиллярам,
- стимулируются, главным образом, растяжением легочных сосудов,
- быстро реагируют на введение химических веществ в легочные сосуды,
- стимуляция может вызвать апноэ, затем учащение дыхания, уменьшение давления, брадикардию и бронхоспазм.
Возбуждение механорецепторов верхних ДП вызывает возникновение дыхательных рефлексов.
Защитные дыхательные рефлексы:
- Чихание – с рецепторов слизистой носа.
- Кашель – с ирритантных рецепторов слизистой гортани, трахеи, бронхов.
- Рефлекс ныряльщиков – остановка дыхания при действии воды на носовые ходы.
- Остановка дыхания во время акта глотания .
- Рефлексогенная задержка дыхания – сужение голосовой щели, бронхоконстрикция при вдыхании дыма, газов, едких веществ.
Механорецепторы межреберных мышц и диафрагмы (проприорецепторы) – они осуществляют обратную связь дыхательных мышц с ДЦ.
Барорецепторы рефлексогенных зон – возбуждаются они при увеличении АД и дыхание при этом угнетается. При уменьшении АД – дыхание усиливается.
Ритмическая смена дыхательных фаз
- Генератор центральной инспираторной активности (возбуждения) – сокр. ЦИА (В) – представлен α-инспираторными нейронами дорсального ядра. Они возбуждаются от хеморецепторов (центральных и сосудистых рефлексогенных зон). Чем больше раздражение хеморецепторов (ХР), тем больше скорость ЦИА .
- Механизм выключения инспирации – состоит из β-инспираторных нейронов и инспираторно-тормозных (ИТ), т.е. экспираторных нейронов. β-инспираторные нейроны возбуждаются афферентными сигналами от РРЛ. На механизм выключения вдоха влияют также нисходящие импульсы от ПТЦ.
Хеморецепторы возбуждаются постоянно и постоянно посылают импульсы в продолговатый мозг, возбуждая α-инспираторные нейроны. Они возбуждаются и посылают импульсы к мотонейронам спинного мозга.
Мотонейроны возбуждаются и посылают импульсы к мышцам. Они сокращаются и наступает вдох .
При вдохе объем легких увеличивается и возбуждаются РРЛ, которые посылают возбуждающие импульсы по чувствительным волокнам блуждающего нерва к β-инспираторным нейронам. В результате суммации импульсов от α-инспираторных нейронов и рецепторов растяжения легких достигается порог и возбуждаются и β-инспираторные нейроны благодаря влиянию вышележащих отделов ЦНС.
Инспираторно-тормозные нейроны посылают тормозные импульсы к α-инспираторным нейронам. В результате α-инспираторные нейроны тормозятся и не посылают импульсы к мотонейронам. Мышцы расслабляются, происходит выдох .
К β-инспираторным нейронам не поступают импульсы, и они тормозятся (не возбуждаются).
β-инспираторные нейроны не возбуждают инспираторно-тормозные нейроны и поэтому они не посылают импульсы к α-инспираторным нейронам. α-инспираторные нейроны вновь возбуждаются импульсами от хеморецепторов и наступает вдох .
Читайте также: