
Множественные аллели и полигенное наследование на примере человека кратко
Обновлено: 05.07.2024
В множественные аллели это различные вариации, которые может содержать данный ген. У всех генов есть два аллеля, которые определяют генетические особенности живых организмов.
В генетике разные аллели одного и того же гена, которые представляют множественные аллелизмы, известны как аллельные серии, и члены одного и того же аллельного ряда могут иметь разную степень доминирования по отношению к другим членам этого ряда.
Одна из ветвей генетики, отвечающая за изучение генов с множественными аллелями, - это хорошо известная популяционная генетика, очень полезная для анализа генетического состава видов, будь то животные, растения или микроорганизмы.
Концепция множественных аллелей
Концепция множественных аллелей в какой-то мере применима чисто популяционным образом, поскольку индивидуум, рассматриваемый с генетической точки зрения, имеет ряд аллелей гена, эквивалентных его хромосомной нагрузке.
Другими словами, диплоидные организмы (2n, с двумя наборами хромосом), такие как, например, млекопитающие, имеют только две альтернативные формы каждого гена, поскольку они наследуют гомологичную хромосому от каждого из своих двух родительских особей во время полового размножения. .
Растения, которые являются классическим примером организмов с более чем двумя наборами гомологичных хромосом (полиплоидов), обладают, индивидуально говоря, таким же количеством аллелей гена, как и его число плоидности, то есть четыре аллеля тетраплоидов (4n) , шесть для гексаплоидов (6n) и так далее.
Понимая это, можно гарантировать, что ген имеет несколько аллелей, если он имеет больше, чем количество аллелей, эквивалентное его хромосомной нагрузке в популяции. Многие авторы придерживаются мнения, что большинство генов в популяции представлено множественными аллелями, которые являются результатом различных видов генных вариаций.
Наследование множественных аллелей
Учитывая, что эта концепция является популяционной, наследование гена с множественными аллелями не отличается от наследования генов, которые имеют только две альтернативные формы, поскольку у диплоидного индивида, например, только через половое размножение. Будут переданы две формы одного и того же гена, по одной на каждую гомологичную хромосому.
Единственное реальное отличие от генов с множественными аллелями и генов, которые существуют только в двух альтернативных формах, заключается в том, что с первым можно достичь значительно большего разнообразия генотипов и фенотипов для определенного признака.
Количество генотипов, возникших в популяции и обусловленных наличием генов с множественными аллелями, является функцией количества аллелей, существующих для каждого данного гена.
Таким образом, если имеется 2, 3, 4 или 5 различных аллелей для одного и того же гена в популяции, соответственно будут наблюдаться 3, 6, 10 или 15 возможных генотипов.
Таким образом, гены с множественными аллелями в популяции следуют принципам сегрегации, предложенным Менделем, поэтому их наследование не отличается от генов с двумя аллелями.
Примеры
В литературе можно найти различные примеры символов, кодируемых множеством аллелей в естественных популяциях. Среди наиболее цитируемых - определение группы крови у людей, цвета шерсти кроликов, цвета глаз у плодовых мух и модели оперения у уток.
Группы крови АВО у человека
Локус, к которому принадлежит ген ABO, определяет группу крови человека. Было описано, что для этого локуса человеческие популяции имеют три возможных аллеля, которые кодируют три разных антигена, определяющих группу крови.
Три аллеля локуса ABO известны как:
– яК, который кодирует антиген А,
– яB, который кодирует антиген B,
– я, который не кодирует какой-либо антиген.
Отношения доминирования между этими тремя аллелями яК> я; яB> я; яК= ЯB (кодоминантность). Оба аллеля К как аллель B доминируют над аллелем я, но они кодоминантны друг с другом; значит, у человека с группой крови AB есть аллель К и один аллель B.
Ввиду того, что аллель я рецессивен, люди с одной группой крови (фенотипом) ИЛИ имеют два аллеля я.
Цвет меха у кроликов
Цвет шерсти кроликов определяется аллельным рядом локуса C. Аллели в этой серии: C, cch, cчас Y c, которые определяют однородную темную окраску, светло-серый (шиншилла), альбинос с темными конечностями и полностью альбинос, соответственно.
Преобладание этих аллелей в порядке от наиболее доминантного к рецессивному, как написано: C> cch> cчас> cСледовательно, может быть 10 различных генотипов, которые порождают только четыре конкретных фенотипа.
Особенности окраски оперения уток
Локус, определяющий структуру оперения уток кряквы, имеет несколько аллелей. Аллель M это тот, который кодирует "дикий" образец, но есть два других аллеля: аллель Mр, что дает образец, известный как "ограниченный", и аллель м¸, который создает узор, известный как «сумеречный" (Темно).
Доминантный аллель Mр, за которым следует аллель M и рецессивный мd, из которых получается шесть возможных комбинаций, дающих начало шести фенотипам.
Полигенное наследование, также известное как количественное наследование, относится к одному унаследованному фенотипическому признаку, который контролируется двумя или более различными генами.
В системе, которая отличается от менделевской генетика где моногенные признаки определяются различными аллелями одного ген полигенетические признаки могут отображать ряд возможных фенотипов, определяемых рядом различных генов и взаимодействиями между ними.
Черты, которые определяются полигенным наследованием, являются не просто следствием доминирования и рецессивности и не проявляют полное доминирование как в менделевской генетике, где один аллель доминирует или маскирует другого. Вместо, полигенные признаки экспонат неполное доминирование так что фенотип в потомстве представлена смесь фенотипов, отображаемых у родителей. Каждый из генов, который вносит вклад в полигенный признак, оказывает одинаковое влияние, и каждый из аллелей оказывает аддитивное влияние на исход фенотипа.
Из-за моделей режима наследования, физические черты, которые контролируются полигенным наследованием, такие как цвет волос, рост и кожа цвет, а также невидимые черты, такие как кровь Давление, интеллект, аутизм и долговечность происходят по непрерывному градиенту со многими вариациями количественных приращений.
Полигенное наследование не следует путать с последствиями, вызванными несколько аллелей, В случае нескольких аллелей ген содержит несколько разных вариантов аллелей в одном и том же локусе каждого хромосома Например, три разных аллеля, которые контролируют группу крови – A, B & O.
Вероятность того, что потомство унаследует определенную характеристику от своих родителей, может быть определена с помощью квадрата пуннета, однако в действительности может быть большое количество разных генов, контролирующих один признак фенотипа, поэтому это становится трудно продемонстрировать. К счастью, распределение фенотипов, определяемых посредством полигенного наследования, обычно вписывается в нормальное распределение вероятностей, при этом у большинства потомков обнаруживается промежуточный фенотип двух родителей.
Используя упрощенный пример полигенетического признака, контролируемого только тремя генами, это становится легче визуализировать.
Примеры полигенного наследования
Цвет кожи
Пигмент меланин отвечает за темное окрашивание кожи, и существует по крайней мере три гена, которые контролируют цвет кожи человека. Используя гипотетический пример, где производство меланина контролируется участвующими аллелями (обозначаемыми здесь как A, B и C), что приводит к темному цвету кожи, и, следовательно, светлый цвет кожи создается не способствующими аллелями (обозначаемым здесь как a, b и в) можно увидеть, как спектр разных цветов кожи может привести к потомству.
Здесь важно помнить, что при полигенном наследовании аллели не проявляют доминирования над другими, скорее, каждый способствующий аллель дает аддитивный эффект, а не маскирующий эффект, и поэтому способ взаимодействия аллелей отличается от такового в менделевской генетике. Аддитивный эффект означает, что каждый способствующий аллель производит одну единицу цвета.
В примере с использованием двух родителей, гетерозиготный для каждого из генов, продуцирующих меланин (AaBbCc x AaBbCc), можно увидеть, как аддитивные эффекты и комбинации аллелей приводят ко всем возможным генотипам.

В этом упрощенном примере имеется 64 возможных комбинации аллелей, в результате которых получается 7 различных оттенков кожи.
Тоны кожи, которые встречаются с наименьшей вероятностью, являются следствием полностью гомозиготных генотипов. Самый светлый цвет кожи, 0 (aabbcc), в котором отсутствуют какие-либо аллели, способствующие пигменту меланина, или самый темный цвет кожи, 6 (AABBCC), который содержит все возможные способствующие аллели; каждый из этих фенотипов встречается с вероятностью 1/64.
Поскольку количество способствующих аллелей изменяется в комбинациях аллелей, единицы пигмента меланина увеличиваются и уменьшаются; вероятность появления второго самого светлого или самого темного тона кожи (1 или 5) составляет 6/64, третьего самого светлого или самого темного тона кожи (2 или 4) составляет 15/64, а наиболее промежуточный тон кожи (3) является наиболее распространенным. в 20/64. Как показано на гистограмме выше, этот шаблон соответствует нормальному распределению.
Высота человека
Рост человека является чрезвычайно сложным паттерном наследования, поскольку его контролируют более 400 генов, поэтому крайне сложно предсказать, какой будет высота потомства; у двух невысоких родителей может быть высокий ребенок, тогда как у двух высоких родителей может быть невысокий ребенок, а у родителей совершенно разного роста может быть высокий, низкий или средний ребенок.
Кроме того, рост известен как многофакторная черта, что означает, что черта подвержена влиянию множества генов, а также влиянию окружающей среды. Например, факторы, относящиеся к общему здоровью растущего ребенка, такие как доступ к еде и подверженность болезням, могут существенно повлиять на конечный рост человека. Подавляющее большинство наших признаков являются многофакторными, поэтому зачастую трудно оценить влияние отдельных генов на возникающий фенотип
- Менделевская генетика – Набор теорий, предложенных Грегором Менделем, которые пытаются объяснить закономерности наследования генетических характеристик на основе простых экспериментов по разведению с участием отдельных генов на парах хромосом.
- Генотип – генетический состав человека организм,
- Фенотип – Физические и биологические характеристики, выраженные в человеке в зависимости от их генотипа.
- эпистаз – взаимодействия между отдельными генами, в которых один маскирует эффект другого.
викторина
1. При полигенном наследовании признаки определяются:A. Несколько аллелей в одном локусеB. Взаимодействие нескольких геновC. Два доминантных аллеля на генеD. Один ген маскируется другим
Ответ на вопрос № 1
В верно. Полигенные признаки определяются взаимодействиями между несколькими разными генами, которые демонстрируют неполное доминирование.
2. Сколько разных комбинаций аллелей можно получить от двух родителей, которые гетерозиготны по полигенному признаку, контролируемому тремя разными генами с двумя парами аллелей?A. 5B. 7C. 54D. 64
Ответ на вопрос № 2
D верно. Каждый родитель может дать 8 различных комбинаций аллелей (ABC, ABc, AbC, aBC, Abc, aBc, abC, abc), поэтому два родителя вместе могут создать 64 различные комбинации, что приведет к 7 возможным различным фенотипам.
3. Какой из следующих фенотипов вряд ли будет многофакторной чертой, подверженной влиянию окружающей среды?A. Риск рака легкихB. Синдром ДаунаC. Высокая масса телаD. Шизофрения
Ответ на вопрос № 3
В верно. Возникновение синдрома Дауна у человека, скорее всего, будет определяться в основном генетическими факторами, так как это состояние с рождения, в то время как на другие состояния, вероятно, будут влиять его воспитание или окружение.
Определение понятия и сущность полигенного наследования, примеры полимерии. Особенности формирования полигенных признаков. Тип аддитивного действия генов. Формы кумулятивной полимерии и сущность доминантного эпистаза. Концепция полигенных болезней.
| Рубрика | Биология и естествознание |
| Вид | статья |
| Язык | русский |
| Дата добавления | 14.11.2014 |
| Размер файла | 31,4 K |
Студенты, аспиранты, молодые ученые, использующие базу знаний в своей учебе и работе, будут вам очень благодарны.
1. Сущность полигенного наследования
Наряду с отдельными генами, представленными множеством форм, существуют и полигенные признаки, т.е. признаки, контролируемые многими генами, находящимися в разных участках хромосомы, а иногда даже и в разных парах хромосом. У человека среди известных нам примеров этого рода можно назвать такие признаки, как рост, умственные способности, телосложение, а также цвет волос и цвет кожи.
Взаимодействие нескольких неаллельных генов с одинаковым действием называется полимерией, или однозначным действием генов.
Примером полимерии является наследование цвета кожи у человека. Этот признак определяется четырьмя генами, ответственными за выработку пигмента меланина. Чем больше активных генов, запускающих синтез меланина, имеется в клетке, тем темнее ее окраска. Эти гены локализованы в четырех парах разных хромосом. У людей с самой темной окраской кожи (негры) имеется восемь аллелей этих генов (поскольку клетки диплоидны): A1A1A2A2A3A3A4A4 (гены действуют одинаково, поэтому их обозначают одной и той же буквой). У человека с самой светлой кожей нет ни одного активного аллеля: а1а1а2а2а3а3а4а4. Дети таких двух людей получат четыре активных (доминантных) аллеля от одного из родителей и цвет их кожи будет промежуточным. Генотип детей будет: A1a1, а2А2, a3A3, а4A3.
В зависимости от числа доминантных генов в генотипе может формироваться более светлый или более темный цвет кожи. Такой тип взаимодействия генов называется кумулятивной полимерией.
Полигенные признаки с трудом поддаются изучению, потому что непросто вычленить эффект каждого отдельного гена в данном фенотипическом признаке, отделив его от эффекта других генов. Влияние среды еще больше запутывает дело; скажем, в нашем примере с цветом кожи люди могут быть более или менее смуглыми в зависимости от интенсивности загара. эпистаз полигенный наследование кумулятивный
- непрерывные (чаще - количественные) признаки, представленные в человеческих популяциях (семьях) множеством фенотипических классов, между которыми нет четкой границы.
Среда всегда оказывает существенное влияние на формирование ПП. Под влиянием среды может появиться даже новый фенотипический класс.
Наследуемость (Н) - >50% 2 или Кн, который рассчитывается по формуле: Кн = G/Е x 100%, где Кн - коэффициент наследования, G - наследственные факторы, Е - факторы окружающей среды. В таблице 4 приведены значения ряда коэффициентов наследования мультифакториальных признаков и заболеваний.
* Степень выраженности признака или тяжесть течения болезни у пробанда. Чем сильнее выражен признак или тяжелее протекает заболевание у больного родственника, тем выше риск его развития у здоровых родственников.
* Общность генов у пробанда и его родственников (или близкая степень родства с больным родственником). Чем больше общих генов у больного и его родственников, тем выше риск развития у последних признака или заболевания. Например, популяционная частота псориаза составляет 0,75%. У родственников I степени родства частота его развития - 5,6%, у родственников II степени родства - 3,0-3,5%, у родственников III степени родства - 1,75%, у родственников IV степени родства - 0,75%.
* Редко поражаемый пол. Мультифакториальный признак или заболевание проявляется чаще у лиц редко поражаемого пола (критерий, названный эффектом Картера). Например, врожденный пилоростеноз у мальчиков встречается в 2-5 раз чаще, чем у девочек, т.е. в данном случае женский пол - редко поражаемый пол. Однако частота этой болезни у будущих детей пораженных пилоростенозом девочек достигнет 10-20%, тогда как у будущих детей пораженных пилоростенозом мальчиков - только 2-6%. Другой пример - язвенная болезнь желудка и двенадцатиперстной кишки, как правило, проявляющаяся у лиц мужского пола и гораздо реже - у лиц женского пола. Однако ее частота у детей больной женщины выше, чем у детей больного мужчины.
* Число больных родственников. Чем больше в родословной родственников, имеющих мультифакториальный признак или заболевание, тем выше риск его развития у потомков
Кумулятивная полимерия. Значительная часть признаков у эукариот, наследуемых полигенно, находится под контролем не двух-трех, а большего числа генов (их количество пока еще трудно определить).
Тип аддитивного действия генов, каждый из которых оказывает свою, часто небольшую, долю влияния на признак, называется кумулятивной полимерией. Используя решетку Пеннета, можно подсчитать частоты доминантных генов среди генотипов второго поколения. Для этого в каждой из 64 клеток вместо генотипа записывается число присутствующих в нем доминантных аллелей. Определив частоты доминантных аллелей, можно убедиться, что генотипы с числом доминантных генов 6,5,4,3, 2, 1,0 встречаются 1,6,15,20,15,6,1 раз соответственно. Эти данные представлены в виде графика на рисунке. На горизонтальной оси указано число доминантных генов в генотипе, а на вертикальной - частоты их встречаемости. С увеличением числа генов, определяющих один признак, этот график приближается к идеальному нормальному распределению.
К количественным относятся признаки, варьирующие более или менее непрерывно от одной особи к другой, что позволяет распределить особей по классам в соответствии со степенью выраженности признака. На рисунке приведен пример распределения по росту у мужчин. Эта выборка разделена на 7 классов с 5 см-интервалом. Мужчины со средним ростом (171-175 см) составляют большую часть выборки. С наименьшей частотой встречаются мужчины, которые включены в класс с ростом 156--160 см и 186--190 см. С увеличением выборки и с уменьшением классового интервала график может приблизиться к нормальному распределению по росту. Фенотипическая изменчивость без разрывов в проявлении, представленная на графике нормального распределения признака, называется непрерывной. Непрерывная изменчивость количественных признаков зависит от двух причин: 1) от генетического расщепления по большому числу генов, 2) от влияния среды, как причины модификационной изменчивости. Впервые датский генетик Иогансен показал, что непрерывная изменчивость такого количественного признака как масса бобов фасоли Phaseolus vulgaris зависит как от генетических, так и средовых факторов. Путем инбридинга в течение ряда поколений он вывел несколько чистых (гомозиготных) линий, различающихся по средней массе бобов. Например, средняя масса бобов в линии 1 была 642 мг, в линии 13 --454 мг, в линии 19 -- 351 мг. Далее Иоган-сен вел отбор крупных и мелких бобов в каждой линии с 1902 по 1907 г. Вне зависимости от массы родительских семян средняя масса бобов после 6 лет отбора была такой же, как и в исходной линии. Так влинии № 13 при массе родительских семян от 275 мгдо 575 мг средняя масса семян в потомстве сохранилась на том же уровне ±450 мг. При этом в каждой линии масса бобов варьировала от минимальных до максимальных значений, а наиболее многочисленным был класс со средней массой, что характерно для количественных признаков. Отбор в чистых линиях оказался невозможен .
Кумулятивная полимерия имеет 2 формы:
1. Аддитивная полигения без порога действия.
формирование признака (болезни).
Предел развития признака (норма реакции) детерминируется общим числом полигенов в генотипе.
Аддитивная полигения с порогом действия
Признак (болезнь) проявляется в том случае, если в генотипе окажется пороговая величина генов.
Так наследуются : гипертоническая болезнь, нормальное развитие верхней губы.
Некумулятивная полимерия. Наряду с кумулятивной (аддитивной) полимерией известны случаи наследования по типу некумулятивной (неаддитивной) полимерии, когда характер проявления признака не меняется в зависимости от числа доминантных полимерных генов.
Эпистаз - подавление действия гена, находящегося в одной неаллельной паре, действием гена из другой неаллельной пары, например подавление геном А гена В, т.е. A >B или A >bb. Выделяют доминантный и рецессивный эпистаз.
Доминантный эпистаз: доминантный аллель одной неаллельной пары, находящийся в гомозиготном (АА) или гетерозиготном (Аа) состоянии, подавляет проявление неаллельного к нему доминантного аллеля другой аллельной пары, находящейся в состоянии АА или Аа. Гены, дающие доминантный эффект, называются эпистатическими генами или супрессорами (ингибиторами). 0ни могут быть как доминантными, так и рецессивными. Подавляемые гены именуются гипостатическими генами.
Если гены, находящиеся в других неаллельных парах, усиливают доминантное действие эпистатических генов, то они называются генами-модификаторами (интенсификаторами).
Такой тип взаимодействия характерен для неаллельных генов, участвующих в регуляции онтогенеза, например генов иммунного ответа (генная сеть - 2190 генов; см. главу 15) или генов эритропоэза (генная сеть - 200 генов).
Возможны два варианта доминантного эпистаза:
* гомозиготы с рецессивными аллелями (аа) отличаются по фенотипу от гомозигот с доминантными аллелями (АА);
* гомозиготы по доминантным аллелям (АА) не отличаются по фенотипу от гомозигот по рецессивным аллелям (аа).
Кроме рецессивного эпистаза, выделен двойной рецессивный эпистаз; при нем у рецессивных генов собственное фенотипическое проявление, а в двойных гомозиготах рецессивные аллели подавляют друг друга: аа >bb, bb >аа.
4. Наследование полигенное
Тип наследования признаков, обусловленных действием многих генов, каждый из которых оказывает лишь слабое действие. Фенотипически проявление полигенно обусловленного признака зависит от условий внешней среды. У потомков наблюдается непрерывный ряд вариаций количественного проявления подобного признака, а не появление четко различающихся по фенотипу классов. В ряде случаев при блокировании отдельного гена признак не проявляется вообще, несмотря на его полигенную обусловленность. Это свидетельствует о пороговом проявлении признака.
Комплементарность, эпистаз и полимерия (о которых Вы спрашивали в другом вопросе) и есть разные случаи полигенного наследования.
В некоторых семьях встречаются болезни, наследование которых отличается от наследованияхромосомных или моногенных болезней . Эти болезни называют полигенными, или мультифакториальными . Основные сведения о природе полигенных болезней были получены в популяционных исследованиях и с помощью близнецового метода. Оказалось, что конкордантность однояйцевых близнецов по проявлению полигенных болезней и семейный повторный риск таких болезней выше, чем следовало бы ожидать при их случайном распределении, но ниже, чем должно было бы быть при их менделевском наследовании (даже при условии неполной пенетрантности или низкой экспрессивности генетического дефекта).
Давно известно, что распространенные пороки развития (например, гипертоническая болезнь ,ИБС , сахарный диабет , язвенная болезнь и шизофрения ) и врожденные нарушения (например,расщелина губы , расщелина неба , позвоночная расщелина, миеломенингоцеле в сочетании санэнцефалией и врожденные пороки сердца ) носят семейный характер, что свидетельствует об их зависимости от наследственных факторов. Это - полигенные болезни. Их этиология очень разнообразна. Расщелину губы и неба иногда вызывают дефекты одного гена, иногда хромосомные аномалии, но чаще всего - многочисленные наследственные и внешние факторы. Аналогично ИБС изредка обусловлена дефектами одного гена (например, при семейной гиперхолестеринемии ), но чаще всего - множеством факторов.
Концепция полигенных болезней предполагает, что каждый конкретный случай вызывается взаимодействием множества наследственных и внешних факторов, что приводит к семейной предрасположенности к болезни, но без четкого менделевского наследования.
Полигенные болезни обусловлены взаимодействием множества независимых генов. Человек, унаследовавший определенную комбинацию генов, предрасположен к болезни, развитию которой способствуют также неблагоприятные внешние факторы. Другой член семьи имеет такую же предрасположенность к болезни если он унаследовал сходную комбинацию генов. Так как у родных братьев и сестер 50% аллелей одинаковые, вероятность того, что кто-либо из них имеет ту же комбинацию генов, составляет (1/2) в степени n, где n -число генов, необходимых для проявления признака (предполагается, что сцепление генов отсутствует).
- при полигенном наследовании часто наблюдается неравномерное распределение признака между полами;
- повторный риск полигенной болезни зависит от пола, тяжести болезни, степени генетической предрасположенности (различающейся в разных семьях), частоты болезни в данной семье, распространенности болезни среди населения.
Поскольку точное число генов, обусловливающих полигенные признаки, неизвестно, степень риска оценивают исходя из эмпирических данных, то есть числа больных в описанных семьях.
В отличие от моногенных заболеваний , когда 25 или 50% родственников больного первой степени также больны, полигенные болезни обычно выявляют у 5-10% таких родственников. Более того, риск возникновения полигенных болезней в разных семьях неодинаков, причем обычно риск заболевания тем выше, чем больше родственников больны и чем тяжелее протекает у них болезнь. Например, риск заболевания у братьев и сестер ребенка с односторонней расщелиной губы составляет 2,5%, а с двусторонней расщелиной губы - 6%.
Полагают, что полигенная этиология характерна для многих болезней, которые развиваются после подросткового периода. Болезни, развивающиеся в более позднем возрасте, в среднем меньше зависят от наследственных факторов. Выявлено девять полигенных болезней, в которых роль наследственных факторов снижается с возрастом.
Подобные документы
Неаллельные гены как гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Комплементарность: понятие, примеры. Доминантное и рецессивное взаимодействие неаллельных генов. Понятие о кумулятивной и некумулятивной полимерии.
презентация [1,1 M], добавлен 07.12.2013
Понятие "неаллельные гены". Исследование фенотипического проявления признаков при дигибридном скрещивании особей в случае различных форм взаимодействия неалельных генов – комплементарности, доминантного и рецессивного эпистаза, гипостаза и криптомерии.
презентация [1,7 M], добавлен 14.05.2015
История формирования и основные положения хромосомной теории. Изучение закономерностей сцепленного наследования генов. Определение биологического значения кроссинговера. Открытие Т. Морганом явления наследования, сцепленного с полом у дрозофилы.
реферат [28,5 K], добавлен 05.12.2010
Основные типы взаимодействия неаллельных генов. Комплементарное взаимодействие на примере наследования формы гребня у кур. Расщепление по фенотипу. Эпистатическое взаимодействие генов. Доминантный эпистаз на примере наследования масти у лошадей.
презентация [121,3 K], добавлен 12.10.2015
Сущность генеалогического метода и его применение в генетике человека. Особенности наследования различных признаков. Гипотеза и ход исследования родословной. Генетические закономерности наследования признаков человека и сравнение результатов с гипотезой.
Множественный аллелизм. Генетика групп крови человека
Множественный аллелизм
Наследование групп крови системы ABO
Понятие об антигенных системах групп крови
Взаимодействие генов
Тестовые задания
Множественный аллелизм
Развитие признака определяется двумя аллелями одного гена (А и а), которые занимают идентичные локусы гомологичных хромосом. Иногда ген имеет не два, а большее число аллелей, которые возникают в результате мутации. Многократное мутирование одного и того же гена образует серию множественных аллелей, а само явление называется явлением множественного аллелизма. Оно имеет широкое распространение: окраска шерсти у кроликов, цвет глаз у дрозофилы, система групп крови АВО у человека.
Имеются определенные закономерности множественного аллелизма:
— каждый ген может иметь большое число аллелей;
— любой аллель может возникнуть в результате прямой и обратной мутации любого члена серии множественных аллелей или от аллеля дикого типа;
— в диплоидном организме могут одновременно находиться два любых аллеля из серии множественных аллелей;
— аллели находятся в сложных доминантно-рецессивных отношениях между собой: один и тот же аллель может быть доминантным по отношению к одному аллелю и рецессивным по отношению к другому, а между иными аллелями доминирование может отсутствовать, и наблюдается кодоминирование и др.;
— члены серии множественных аллелей наследуются так же, как и пара аллелей, т. е. наследование подчиняется менделевским закономерностям (кроме кодоминирования);
— разные сочетания аллелей в генотипе обуславливают различные фенотипические проявления одного и того же признака;
— серии аллелей увеличивают комбинатов ну ю изменчивость.
Наследование групп крови системы ABO
Примером множественного аллелизма у человека является наличие трех аллелей гена, определяющего наследование групп крови системы АВО.
• система определяется тремя аллелями одного гена I (IA, 1в, 1°); ген I расположен в 9-й хромосоме: 9q34;
• из всей серии аллелей одновременно в генотипе диплоидного организма находятся два аллеля (I°I0, IAIA, IAI°, 1в1в и др.);
• аллели IA, 1в доминантны по отношению к аллелю 1° — полное доминирование, между собой аллели 1А и 1в — кодоминантны;
• доминантный аллель гена может проявлять свое действие в гомо- (IAIA, IBIB) и гетерозиготном организмах (1А1°, 1в 1°), а рецессивный аллель гена — только в гомозиготном организме (1° 1°);
• различные сочетания аллелей в генотипе дают разные фенотипы: 4 группы крови I (0), II (А), III (В), IV (АВ), которые различаются между собой антигенными свойствами эритроцитов. Антигены (агглютиногены) находятся на поверхности эритроцитов (гликокаликс);
• особенностью системы является наличие в сыворотке крови спецефических антител (агглютининов), разноименных по отношению к собственным агглютиногенам (они одновременно находятся в крови);
• разнообразие групп крови обеспечивает фенотипический полиморфизм в популяциях человека по данному признаку.
Ген I обладает 100% пенетрантностью.
Группы крови являются примером однозначной нормы реакции организма (группа крови не изменяется в течение жизни ни при каких изменениях среды).
Понятие об антигенных системах групп крови
В Европе у 85% людей на поверхности эритроцитов имеется антиген (белок), который называется резус-фактор (они условно называются резус-положительными), у 15% — в эритроцитах такого антигена нет (они условно называются резус-отрицательными). Наличие антигена определяют три тесно сцепленных гена С, Д, К (хромосома 1р35), они наследуются совместно как один ген и обозначаются одним символом Rh. Резус-фактор наследуется по доминантному типу (наличие резус-фактора RhRh, Rhrh, его отсутствие — rhrh). Иногда при переливании крови и некоторых вариантах браков возникает резус-несовместимость. Классическим примером несовместимости по резус-фактору матери и плода является вариант, когда в организме резус-отрицательной матери развивается резус-положительный плод.
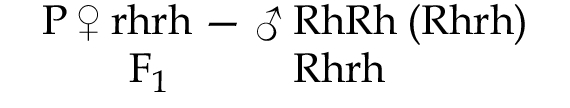
Кровь матери и плода разделена (плацента — биологическая мембрана), эритроциты через мембрану не проникают. Во время родов эритроциты плода, имеющие антиген, могут попасть в кровоток матери и вызвать образование антител. Антитела свободно перемещаются через плаценту из организма матери в организм плода. Эритроциты второго ребенка с момента начала кроветворения в эмбриогенезе уже будут подвергаться воздействию образовавшихся антител матери (реакция антитело-антиген на ПАК эритроцитов) и разрушаться (гемолиз эритроцитов). В результате гемолиза эритроцитов возникает гемолитическая болезнь, симптомами которой являются анемия, желтуха, водянка. Для спасения ребенка после рождения ему осуществляют обменное переливание одногрупповой крови, при котором восстанавливается дыхательная функция крови и удаляются токсические продукты.
Для профилактики гемолитической болезни у последующих детей женщине до родов или сразу после родов вводят антирезусные антитела, которые связывают антигены (эритроциты плода), попавшие в кровь (эффект Кларка — профилактика резус-несовместимости). Иммунная система матери остается интактной.
Определение групп крови систем АВО и РЕЗУС используется при изучении близнецов (определение конкордантности по изучаемому признаку в парах монозиготных и дизиготных близнецов), судебно-медицинской экспертизе, переливании крови, трансплантации, установлении отцовства и др.
Для изучения систем групп крови нужно знать иммуногенетику: строение антигенов, взаимоотношения между антигенами и антителами, возможность агглютинации эритроцитов при несовместимости по группам крови донора и реципиента, беременной и плода.
Взаимодействие генов
В настоящее время в эритроцитах обнаружено более 200 различных агглютиногенов, 140 из которых объединены в 20 систем (групп), а остальные являются общими или индивидуальными. Это определяет антигенную неповторимость людей (и в этом смысле каждый человек имеет свою группу крови). Данные системы агглютиногенов отличаются от системы АВО тем, что не содержат в плазме естественных агглютининов, подобных а и Р агглютининам. Среди систем агглютиногенов, кроме системы АВО, наиболее важны Rh; MN, S, Р, А, Levis, Kell, Duppi, Kidd и др. В каждой из этих систем имеется один или несколько агглютиногенов, составляющих разные комбинации.
Например, система Kell состоит из двух агглютиногенов Кик, они образуют 3 группы крови: КК, Кк, кк. Все эти системы агглютиногенов имеют значение лишь при частых переливаниях крови, поэтому в обычной медицинской практике повторно переливать кровь больному от одного и того же донора не рекомендуется.
Читайте также:
- Взаимодействие с родителями по оздоровлению детей раннего возраста в условиях доу и семьи
- Достижения современной космонавтики кратко
- Социальные и философские основы бунта раскольникова кратко
- Сценарий новогоднего праздника во 2 младшей группе детского сада волшебный новый год
- Фон бюлов роль в истории кратко