
Механизм проведения возбуждения по нервному волокну кратко
Обновлено: 01.07.2024
Возбуждение, возникнув в одном участке мембраны возбудимой клетки, обладает способностью распространяться. Длинный отросток нейрона – аксон (нервное волокно) выполняет в организме специфическую функцию проведения возбуждения на большие расстояния.
1. Законы проведения возбуждения по нервным волокнам
Закон анатомической и физиологической непрерывности - возбуждение может распространяться по нервному волокну только в случае его морфологической и функциональной целостности.
Закон двустороннего проведения возбуждения – возбуждение, возникающее в одном участке нерва, распространяется в обе стороны от места своего возникновения. В организме возбуждение всегда распространяется по аксону от тела клетки (ортодромно).
Закон изолированного проведения – возбуждение, распространяющееся по волокну, входящему в состав нерва, не передается на соседние нервные волокна.
2. Особенности проведения местного и распространяющегося возбуждения
Локальный ответ (местное возбуждение)
распространяется по нервным волокнам с затуханием (с декрементом), т.е. амплитуда локального ответа быстро падает с увеличением расстояния от места его возникновения;
вследствие затухания локальный ответ распространяется на небольшие расстояния (не более 2 см);
местное возбуждение распространяется пассивно, без затрат энергии клетки;
механизм распространения местного возбуждения аналогичен распространению электрического тока в проводниках; такой способ распространения возбуждения называют электротоническим.
Потенциал действия (распространяющееся возбуждение)
распространяется по нервным волокнам без затухания, амплитуда потенциала действия одинакова на любом расстоянии от места его возникновения;
расстояние, на которое распространяется потенциал действия, ограничено только длиной нервного волокна;
распространение потенциала действия – активный процесс, в ходе которого изменяется состояние ионных каналов волокна, энергия АТФ требуется для восстановления трансмембранных ионных градиентов;
механизм проведения потенциала действия более сложен, чем механизм распространения местного возбуждения.
3. Миелиновые и безмиелиновые нервные волокна
Миелиновые волокна. Часть нервных волокон в ходе эмбриогенеза подвергается миелинизации: леммоциты (шванновские клетки) сначала прикасаются к аксону, а затем окутывают его (рис 4.1 А, Б). Мембрана леммоцита наматывается на аксон наподобие рулета, образуя многослойную спираль (миелиновую оболочку) (рис 4.1 В, Г). Миелиновая оболочка не является непрерывной – по всей длине нервного волокна на равном расстоянии друг от друга в ней имеются небольшие перерывы (перехваты Ранвье). В области перехватов аксон лишен миелиновой оболочки.
Безмиелиновые волокна. Миелинизация других волокон заканчивается на ранних стадиях эмбрионального развития. В леммоцит погружается один или несколько аксонов; он полностью или частично окружает их, но не образует многослойной миелиновой оболочки
4. Механизм проведения возбуждения по безмиелиновым нервным волокнам
В состоянии покоя вся внутренняя поверхность мембраны нервного волокна несет отрицательный заряд, а наружная сторона мембраны – положительный. Электрический ток между внутренней и наружной стороной мембраны не протекает, т.к. липидная мембрана имеет высокое электрическое сопротивление. Во время развития потенциала действия в возбужденном участке мембраны происходит реверсия заряда (рис. 4.2 А). На границе возбужденного и невозбужденного участка начинает протекать электрический ток (рис. 4.2 Б). Электрический ток раздражает ближайший участок мембраны и приводит его в состояние возбуждения (рис. 4.2 В), в то время как ранее возбужденные участки возвращаются в состояние покоя (рис. 4.2 Г). Таким образом, волна возбуждение охватывает все новые участки мембраны нервного волокна.

Рис. 4.2. Механизм распространения возбуждения по безмиелиновому нервному волокну. Объяснения – в тексте.
5. Механизм проведения возбуждения по миелиновым нервным волокнам
В миелинизированном нервном волокне участки мембраны, покрытые миелиновой оболочкой, являются невозбудимыми; возбуждение может возникать только участках мембраны, расположенных в области перехватов Ранвье. При развитии ПД в одном из перехватов Ранвье происходит реверсия заряда мембраны (рис 4.3 А). Между электроотрицательными и электроположительными участками мембраны возникает электрический ток, который раздражает соседние участки мембраны (рис 4.3 Б). Однако, в состояние возбуждения может перейти только участок мембраны в области следующего перехвата Ранвье (рис 4.3 В). Таким образом, возбуждение распространяется по мембране скачкообразно (сальтаторно) от одного перехвата Ранвье к другому.

Рис. 4.3. Механизм распространения возбуждения по миелиновому нервному волокну. Объяснения – в тексте.
6. Классификация нервных волокон
Нервные волокна различаются по диаметру и степени миелинизации. Чем больше диаметр нервного волокна и степень его миелинизации, тем выше скорость проведения возбуждения. Волокна с разной скоростью проведения выполняют различные физиологические функции. Нервные волокна подразделяются на 6 типов, характеристики которых приведены в таблице 4.1.
Нервные волокна всех групп обладают общими свойствами:
нервные волокна практически неутомляемы;
нервные волокна обладают высокой лабильностью, т.е. могут воспроизводить потенциал действия с очень высокой частотой.
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну.
Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности
Нервного волокна.
Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну.
В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервноговолокна на другое, то нормальное функционирование органов было бы невозможно.
Возбуждение (потенциал действия) распространяется по нервному волокну без затухания.
Периферический нерв практически неутомляем.
Механизм проведения возбуждения по нерву.
Возбуждение (потенциал действия - ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным.

Скорость распространения возбуждения пропорциональна толщине волокна и обратно пропорциональна сопротивлению среды. Проведение возбуждения зависит от соотношения амплитуды ПД и величины порогового потенциала. Этот показатель называется гарантийный фактор(ГФ) и равен 5 - 7, т.е. ПД должен быть выше порогового потенциала в 5- 7 раз. Если ГФ = 1 проведение ненадёжно, если ГФ

Схема распространения возбуждения в безмиелиновых
и миелиновых нервных волокнах.
Законы проведения возбуждения по нервам.
Нервное волокно обладает следующими физиологическими свойствами: возбудимостью, проводимостью, лабильностью.
Проведение возбуждения по нервным волокнам осуществляется по определенным законам.
Закон двустороннего проведения возбуждения по нервному волокну.
Нервы обладают двусторонней проводимостью, т.е. возбуждение может распространяться в любом направлении от возбужденного участка (места его возникновения), т. е., центростремительно и центробежно. Это можно доказать, если на нервное волокно наложить регистрирующие электроды на некотором расстоянии друг от друга, а между ними нанести раздражение. Возбуждение зафиксируют электроды по обе стороны от места раздражения. Естественным направлением распространения возбуждения является: в афферентных проводниках - от рецептора к клетке, в эфферентных - от клетки к рабочему органу.
Закон анатомической и физиологической целостности
Нервного волокна.
Проведение возбуждения по нервному волокну возможно лишь в том случае, если сохранена его анатомическая и физиологическая целостность, т.е. передача возбуждения возможна только по структурно и функционально не измененному, неповрежденному нерву (законы анатомической и физиологической целостности). Различные факторы, воздействующие на нервное волокно (наркотические вещества, охлаждение, перевязка и т. д.) приводят к нарушению физиологической целостности, т. е., к нарушению механизмов передачи возбуждения. Несмотря на сохранение его анатомической целостности проведение возбуждения в таких условиях нарушается.
Закон изолированного проведения возбуждения по нервному волокну.
В составе нерва возбуждение по нервному волокну распространяется изолированно, без перехода на другие волокна, имеющиеся в составе нерва. Изолированное проведение возбуждения обусловлено тем, что сопротивление жидкости, заполняющей межклеточные пространства, значительно ниже сопротивления мембраны нервных волокон. Поэтому основная часть тока, возникающего между возбужденным и невозбужденным участками нервного волокна, проходит по межклеточным щелям, не действуя на рядом расположенные нервные волокна. Изолированное проведение возбуждения имеет важное значение. Нерв содержит большое количество нервных волокон (чувствительных, двигательных, вегетативных), которые иннервируют различные по структуре и функциям эффекторы (клетки; ткани, органы). Если бы возбуждение внутри нерва распространялось с одного нервноговолокна на другое, то нормальное функционирование органов было бы невозможно.
Возбуждение (потенциал действия) распространяется по нервному волокну без затухания.
Периферический нерв практически неутомляем.
Механизм проведения возбуждения по нерву.
Возбуждение (потенциал действия - ПД) распространяется в аксонах, телах нервных клеток, а также иногда в дендритах без снижения амплитуды и без снижения скорости (бездекрементно). Механизм распространения возбуждения у различных нервных волокон неодинаков. При распространении возбуждения по безмиелиновому нервному волокну механизм проведения включает два компонента: раздражающее действие катэлектротона, порождаемое локальным ПД, на соседний участок электровозбудимой мембраны и возникновение ПД в этом участке мембраны. Локальная деполяризация мембраны нарушает электрическую стабильность мембраны, различная величина поляризации мембраны в смежных ее участках порождает электродвижущую силу и местный электрический ток, силовые линии которого замыкаются через ионные каналы. Активация ионного канала повышает натриевую проводимость, после электротонического достижения критического уровня деполяризации (КУД) в новом участке мембраны генерируется ПД. В свою очередь этот потенциал действия вызывает местные токи, а они в новом участке мембраны генерируют потенциал действия. На всем протяжении нервного волокна происходит процесс новой генерации потенциала действия мембраны волокна. Данный тип передачи возбуждения называется непрерывным.
Скорость распространения возбуждения пропорциональна толщине волокна и обратно пропорциональна сопротивлению среды. Проведение возбуждения зависит от соотношения амплитуды ПД и величины порогового потенциала. Этот показатель называется гарантийный фактор(ГФ) и равен 5 - 7, т.е. ПД должен быть выше порогового потенциала в 5- 7 раз. Если ГФ = 1 проведение ненадёжно, если ГФ
ЛЕКЦИЯ № 3. Физиологические свойства нервов и нервных волокон
1. Физиология нервов и нервных волокон. Типы нервных волокон
Физиологические свойства нервных волокон:
1) возбудимость – способность приходить в состояние возбуждения в ответ на раздражение;
2) проводимость – способность передавать нервные возбуждение в виде потенциала действия от места раздражения по всей длине;
3) рефрактерность (устойчивость) – свойство временно резко снижать возбудимость в процессе возбуждения.
Нервная ткань имеет самый короткий рефрактерный период. Значение рефрактерности – предохранять ткань от перевозбуждения, осуществляет ответную реакцию на биологически значимый раздражитель;
4) лабильность – способность реагировать на раздражение с определенной скоростью. Лабильность характеризуется максимальным числом импульсов возбуждения за определенный период времени (1 с) в точном соответствии с ритмом наносимых раздражений.
Нервные волокна не являются самостоятельными структурными элементами нервной ткани, они представляют собой комплексное образование, включающее следующие элементы:
1) отростки нервных клеток – осевые цилиндры;
2) глиальные клетки;
3) соединительнотканную (базальную) пластинку.
Главная функция нервных волокон – проведение нервных импульсов. Отростки нервных клеток проводят сами нервные импульсы, а глиальные клетки способствуют этому проведению. По особенностям строения и функциям нервные волокна подразделяются на два вида: безмиелиновые и миелиновые.
Безмиелиновые нервные волокна не имеют миелиновой оболочки. Их диаметр 5–7 мкм, скорость проведения импульса 1–2 м/с. Миелиновые волокна состоят из осевого цилиндра, покрытого миелиновой оболочкой, образованной шванновскими клетками. Осевой цилиндр имеет мембрану и оксоплазму. Миелиновая оболочка состоит на 80 % из липидов, обладающих высоким омическим сопротивлением, и на 20 % из белка. Миелиновая оболочка не покрывает сплошь осевой цилиндр, а прерывается и оставляет открытыми участки осевого цилиндра, которые называются узловыми перехватами (перехваты Ранвье). Длина участков между перехватами различна и зависит от толщины нервного волокна: чем оно толще, тем длиннее расстояние между перехватами. При диаметре 12–20 мкм скорость проведения возбуждения составляет 70—120 м/с.
В зависимости от скорости проведения возбуждения нервные волокна делятся на три типа: А, В, С.
Наибольшей скорость проведения возбуждения обладают волокна типа А, скорость проведения возбуждения которых достигает 120 м/с, В имеет скорость от 3 до 14 м/с, С – от 0,5 до 2 м/с.
2. Механизмы проведения возбуждения по нервному волокну. Законы проведения возбуждения по нервному волокну
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесения раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).
Существует три закона проведения раздражения по нервному волокну.
Закон анатомо-физиологической целостности.
Проведение импульсов по нервному волокну возможно лишь в том случае, если не нарушена его целостность. При нарушении физиологических свойств нервного волокна путем охлаждения, применения различных наркотических средств, сдавливания, а также порезами и повреждениями анатомической целостности проведение нервного импульса по нему будет невозможно.
Закон изолированного проведения возбуждения.
Существует ряд особенностей распространения возбуждения в периферических, мякотных и безмякотных нервных волокнах.
В периферических нервных волокнах возбуждение передается только вдоль нервного волокна, но не передается на соседние, которые находятся в одном и том же нервном стволе.
В мякотных нервных волокнах роль изолятора выполняет миелиновая оболочка. За счет миелина увеличивается удельное сопротивление и происходит уменьшение электрической емкости оболочки.
В безмякотных нервных волокнах возбуждение передается изолированно. Это объясняется тем, что сопротивление жидкости, которая заполняет межклеточные щели, значительно ниже сопротивления мембраны нервных волокон. Поэтому ток, возникающий между деполяризованным участком и неполяризованным, проходит по межклеточным щелям и не заходит при этом в соседние нервные волокна.
Закон двустороннего проведения возбуждения.
Нервное волокно проводит нервные импульсы в двух направлениях – центростремительно и центробежно.
В живом организме возбуждение проводится только в одном направлении. Двусторонняя проводимость нервного волокна ограничена в организме местом возникновения импульса и клапанным свойством синапсов, которое заключается в возможности проведения возбуждения только в одном направлении.
Возбуждение нейрона. Потенциал действия нейрона
При образовании потенциалов действия (спайное) в химических синапсах между нейронами происходит высвобождение медиаторов из пузырьков пресинаптической мембраны. Медиатор проходит через синаптическую щель и активирует рецепторы, встроенные в постсинаптическую мембрану нейрона-мишени. Рецепторы, в свою очередь, активируют медиаторозависимые ионные каналы, изменяющие поляризацию нейрона-мишени. При смещении показателей мембранного потенциала от уровня потенциала покоя (-70 мВ) в отрицательную сторону (например, — 80 мВ и более) происходит гиперполяризация мембраны, а при смещении в положительную сторону — деполяризация.
а) Электротонические потенциалы. Первоначальный ответ нейрона-мишени на воздействие возбуждающих импульсов заключается в формировании локальных ступенчатых электротонических потенциалов. Положительные злектротонические потенциалы обычно возникают в мультииолярных нейронах в результате деполяризации, обусловленной активацией медиаторозависимых ионных каналов. При низкочастотном стимулирующем воздействии по дендритам распространяются небольшие волны деполяризации, угасающие в течение 2-3 мс.
При увеличении частоты воздействия происходит ступенчатая временная суммация возбуждения, в результате чего образуется прогрессивно увеличивающаяся волна, которая распространяется на поверхность сомы. Одновременное схождение в области сомы волн возбуждения, распространяющихся по разным дендритам, приводит к пространственной суммации возбуждения. При деполяризации приблизительно на 15 мВ (и значении потенциала -55 мВ) происходит открытие потенциалозависимых ионных каналов в наиболее чувствительной области нейрона — триггерной точке (триггерной области), располагающейся в начальном сегменте аксона. При достижении уровня деполяризации (генераторного потенциала) пороговых значений, т.е. потенциала, способного запустить открытие потенциалозависимых ионных каналов, происходит формирование потенциала действия.
В триггерной зоне чувствительных нейронов черешнях и спинномозговых нервов формируется так называемый рецепторный потенциал. В этой области расположено большое количество ионных каналов, активирующихся под воздействием специфических чувствительных стимулов и вызывающих ступенчатое распространение волны деполяризации.
Определение триггерной области миелинизированных нервных волокон не вызывает затруднений: у мультиполярных нейронов эта область граничит с первым миелиновым сегментом с его проксимальной стороны, а у периферических чувствительных нейронов — с последним миелиновым сегментом с его дистальной стороны.
Тормозные (гиперполяризующие) постсинаптические потенциалы возникают при открытии ионных каналов, генерирующих выходящий ток (например, калиевых каналов). Распространение тормозных потенциалов также является угасающим.
Временная суммация.
(А) Аксон чувствительного нейрона (выделен голубым цветом) передает единичный импульс двигательному нейрону,
вызывающий возбуждающему постсинаптический потенциал (ПСП), значения которого не достигают пороговых величин, вследствие чего этот потенциал затухает.
(Б) Аксон чувствительного нейрона передает два импульса.
Происходит временная суммация импульсов с достижением в начальном сегменте аксона пороговых значений возбуждения,
что приводит к формированию потенциала действия, который в дальнейшем распространяется по аксону двигательного нейрона. (А) Ступенчатая суммация возбуждающих постсинаптических потенциалов (ПСП), за счет которой происходит формирование пикового потенциала. Пунктирной линией указана область, соответствующая графику А.
(Б) Генераторные потенциалы достаточной силы формируют множественные пиковые потенциалы. Потенциалы действия двигательных и чувствительных нейронов скелетной мускулатуры. ЦНС — центральная нервная система. Основные характеристики потенциала действия.
Возникновение фазы следовой гиперполяризации объясняют тем, что после полной инактивации потенциалозависимых натриевых каналов многие калиевые каналы остаются открытыми. Восстановление ионных градиентов обеспечивает натрий-калиевый насос.
В ходе подробного изучения натриевых каналов было обнаружено, что их регуляция осуществляется двумя путями. В состоянии покоя (-70 мВ) активационные ворота, расположенные в средней части натриевых и калиевых каналов, закрыты. Первыми на пороговое раздражение реагируют натриевые каналы: происходят открытие активационных ворот и быстрый выход ионов Na + в клетку по концентрационному и электрическому градиентам. При достижении потенциалом действия пиковых значений (+35 мВ) происходит открытие калиевых каналов, а натриевые каналы закрываются инактивационными воротами, представленными глобулярным белком. При реполяризации и восстановлении мембранного потенциала (-70 мВ) активационные натриевые ворота закрываются, а затем вновь открываются инактивационные натриевые ворота, тем самым приводя натриевые каналы в исходное состояние.
Работа потенциалозависимых калиевых каналов регулирует одноворотный механизм: этими каналами управляют только активационные ворота.
Во время фазы нарастания и в начале фазы реполяризации потенциала действия нейрон находится в абсолютно рефрактерном состоянии и не может воспринимать следующие импульсы в результате инактивации потенциалозависимых натриевых каналов. Вслед за этим периодом наступает интервал относительной рефрактерности, во время которого стимул, превышающий исходный более чем на 15 мВ, способен вызвать формирование повторного потенциала действия. Часто для достижения показателя 35 мВ генераторный потенциал производит от 50 до 100 импульсов в секунду (частота составляет от 50 до 100 Гц).
Изменения проницаемости потенциалозависимых натриевых и калиевых каналов, обеспечивающих развитие потенциала действия. Потенциалозависимые натриевые каналы в разные фазы потенциала действия.
(А) В состоянии покоя, предшествующем возбуждению, активационные ворота находятся в закрытом состоянии, а инактивационные — в открытом.
(Б) При преодолении порога возбуждения происходит открытие активационных ворот, что приводит к полному открытию ионного канала.
(В) Инактивационные ворота закрываются, что приводит к закрытию ионного канала.
(Г) При восстановлении потенциала покоя активационные ворота закрываются, а инактивационные — открываются.
Ионный канал возвращается в исходное состояние. Рефрактерные периоды.
АРП — абсолютный рефрактерный период;
ОРП — относительный рефрактерный период.
в) Передача нервных импульсов. Изменение потенциала триггерной зоны распространяется вдоль аксона в соответствии с электротонической траекторией. Положительные заряды внутренней стороны мембраны распространяются в обоих направлениях через аксоплазму. Положительные заряды внешней стороны мембраны аналогичным образом проходят через внеклеточную жидкость и нейтрализуют отрицательный заряд на внешней стороне мембраны. Прилежащий проксимальный участок мембраны достаточно рефрактерен и способен противостоять деполяризации. Одновременно с этим в прилежащем дистальном сегменте возникает локальный ответ (деполяризация), который достигает пороговых величин и распространяется далее вдоль ствола и ветвей аксона к нервным окончаниям, обеспечивая проведение потенциала действия.
Проведение импульса по немиелинизированному нервному волокну происходит непрерывно, а по миелинизированному — сальтаторно (скачкообразно). Миелиновая оболочка выполняет функцию изолятора и располагается между перехватами Ранвье, где сконцентрированы натриевые каналы. В связи с этим в каждом перехвате Ранвье последовательно возникают потенциалы действия за счет распространения положительного тока вдоль аксоплазмы миелинизированного участка аксона и его выхода в области следующего перехвата Ранвье. При распространении тока в обратном направлении по внеклеточной жидкости для повторного заряда деполяризованного участка мембраны снятие положительного заряда приводит к деполяризации следующего перехвата Ранвье.
г) Скорость проведения нервных импульсов. Скорость проведения нервного импульса по немиелинизированным нервным волокнам прямо пропорциональна диаметру аксона. В первую очередь это связано с тем, что больший объем аксоплазмы обеспечивает более высокую скорость продольного распространения возбуждения. Кроме того, чем больше диаметр аксона, тем больше площадь поверхности его плазматической мембраны, следовательно, на ней расположено большее количество ионных каналов, а деполяризация и восстановление заряда мембраны происходят быстрее. Диаметр аксонов варьирует от 0,2 до 2 мкм, а скорость проведения импульсов — от 2 до 25 м/с.
Внешний диаметр миелинизированных нервных волокон (с учетом толщины миелинового слоя) варьирует от 2 до 25 мкм. Следует отметить, что чем шире диаметр миелинизированного нервного волокна, тем длиннее миелинизированные участки, расположенные между перехватами Ранвье, в связи с чем потенциалы действия распространяются с большей скоростью, подобно спортсмену, делающему широкие шаги при беге. Скорости проведения нервных импульсов по разным видам периферических нервов описаны отдельной статье на сайте - просим пользоваться формой поиска на главной странице сайта.
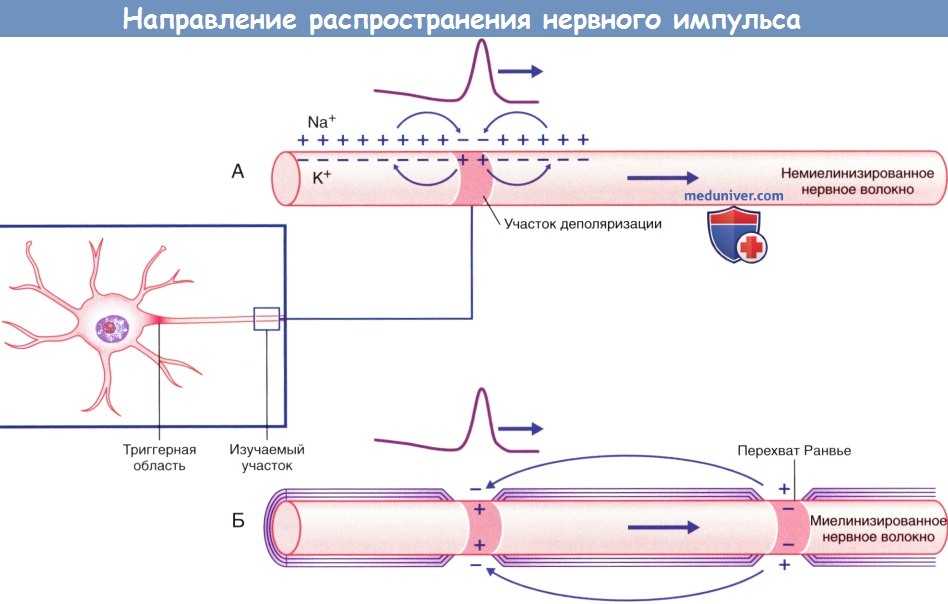
Направление распространения импульса, показанное в виде прохождения положительного заряда.
(А) Непрерывное проведение импульса по немиелинизированному нервному волокну.
(Б) Скачкообразное проведение импульса по миелинизированному нервному волокну.
д) Механизм действия местных анестетиков. При применении местных анестетиков происходит обратимая блокировка проведения нервных импульсов за счет инактивации натриевых каналов, особенно потенциалозависимых, вследствие чего деполяризация мембраны становится невозможной. Молекулы местных анестетиков связывают белок, окружающий ионные каналы. В течение времени действия анестетика происходит связывание белка, окружающего ионные каналы. Для доступа к этим белкам местные анестетики должны быть хорошо растворимы в жирах. При введении анестетика вблизи периферического нерва в первую очередь происходит инактивация немиелинизированных и тонко миелинизированных Аб-волокон. При воздействии на смешанный нерв может возникнуть временный двигательный паралич.
Большинство местных анестетиков — производные амидов (бупивакаин, лидокаин) или сложных эфиров (бензокаин, новокаин). И те, и другие вызывают местное расширение сосудов за счет прямого расслабления гладкой мускулатуры артериол, что приводит к ускорению их выведения. В связи с этим в растворы для местной анестезии часто добавляют адреналин, вызывающий сокращение гладкой мускулатуры и сужение сосудов.
е) Резюме. Электрически заряженные атомы или группы атомов называют ионами. Ионы Na + и К + являются катионами, а ионы Cl + и отрицательно заряженные белки — анионами. Клеточные мембраны представляют собой заряженные конденсаторы с потенциалом покоя величиной -70 мВ.
Неуправляемые натриевые, калиевые и хлорные каналы открыты постоянно, и движение соответствующих ионов через них происходит по градиенту концентрации. Натриевые каналы относительно малочисленны, а калиевые и кальциевые каналы представлены в большом количестве. Ионы К + сконцентрированы в цитозоле за счет притягивающего влияния белков-анионов цитоскелета и отталкивающего действия ионов Na + , расположенных на внешней части плазматической мембраны. Натрий-калиевый насос обеспечивает поддержание мембранного потенциала.
Первоначальная реакция мультиполярного нейрона на возбуждающий стимул заключается в формировании угасающих волн положительного электротонуса. В результате их временной и/или пространственной суммации происходит образование генераторного потенциала в начальном сегменте аксона. При достижении пороговых значений (-55 мВ) потенциалозависимые ионные каналы генерируют потенциал действия, распространяющийся по нервному волокну. Тормозные стимулы вызывают формирование волн отрицательного электротонуса, суммация которых приводит к гиперполяризации мембраны, тем самым отдаляя величины ее потенциала от пороговых значений.
Потенциал действия (спайк) проходит фазу нарастания (деполяризации) от начальных значений до +35 мВ, фазу убывания (реполяризации), в которую возвращается к исходным показателям, и фазу следовой гиперполяризации, в которую понижается до -75 мВ, а затем вновь возвращается к первоначальным значениям. Деполяризация мембраны происходит в результате активации потенциалозависимых натриевых каналов, в ходе которой происходит их стремительное (со скоростью + в клетку и деполяризации мембранного потенциала. При достижении мембранного потенциала +35 мВ происходит закрытие натриевых каналов при помощи инактивационных ворот.
При достижении потенциала действия максимальных значений происходит открытие потенциалозависимых калиевых каналов, и поток ионов Na + в клетку сменяется потоком ионов К + из клетки, что приводит к реполяризации и следовой гиперполяризации мембранного потенциала.
Читайте также:
- Роль воспитателя на физкультурном занятии в детском саду по фгос презентация
- Правовой статус налоговых представителей кратко
- Подпишите фотографии кратко укажите роль каждого из представленных исторических деятелей
- Жизненный цикл информационной системы кратко
- Уровни организации материи неживая природа биологический и социальный уровни кратко