
Кратко опишите роль удвоение центриолей для клеточного цикла
Обновлено: 02.07.2024
Центриоль — внутриклеточный органоид эукариотической клетки. Размер центриоли находится на границе разрешающей способности светового микроскопа.
Эти органеллы в делящихся клетках принимают участие в формированииверетена деления и располагаются на его полюсах. В неделящихся клетках (например, эпителия) центриоли часто определяют полярность клеток и располагаются вблизи комплекса Гольджи.
Термин был предложен Теодором Бовери в 1895 году. Тонкое строение центриолей удалось изучить с помощью электронного микроскопа. В некоторых объектах удавалось наблюдать центриоли, обычно расположенные в паре (диплосома), и окруженные зоной более светлой цитоплазмы, от которой радиально отходят тонкие фибриллы (центросфера). Совокупность центриолей и центросферы называют клеточным центром.
Чаще всего пара центриолей лежит вблизи ядра. Каждая центриоль построена из 27 цилиндрических элементов (тубулиновых микротрубочек), сгруппированных в 9 триплетов. Эти триплеты расположены по окружности, образуя полый цилиндр. Его длина — 0,3-0,5 мкм (равна длине каждого триплета), а диаметр — около 0,15 мкм.
В каждом триплете первая микротрубочка (А-микротрубочка) имеет диаметр около 25 нм, толщину стенки 5 нм и состоит из 13 протофиламентов. Вторая и третья микротрубочки (B и C) отличаются от A-микротрубочки тем, что они являются неполными, содержат 11 протофиламентов и вплотную примыкают к своим соседям. Каждый триплет располагается к радиусу такого цилиндра под углом около 40°.
Центриоли всегда бывают расположены в материале, не имеющем чётко выраженной структуры, который инициирует развитие микротрубочек. Эту область клетки называют центросомой. Именно она образует веретено деления, а не центриоли. Это позволяет объяснить тот факт, почему растения и грибы, не имеющие центриолей, способны образовывать веретено.
Функция центриолей остаётся неизвестной. Возможно, они участвуют в ориентации веретена согласно полюсам, к которым будет происходить деление клетки (цитокинез). Модифицированные центриоли также находятся у основания жгутиков и ресничек у простейших, там их называют базальными тельцами.
В профазе митоза две центриоли расходятся к полюсам клетки и формируют две центросомы. Центросомы в свою очередь служат ЦОМТами (центрами организации микротрубочек) веретена деления. Однако от этой общей схемы существует масса отклонений. Во многих клетках центриоли многократно удваиваются за один клеточный цикл. При созревании яйцеклеток у подавляющего большинства животных центриоли разрушаются (при этом многие белки, входящие в состав центросом, по-прежнему присутствуют в клетке).
При образовании сперматозоидов, напротив, деградирует центросома; одна из центриолей превращается в базальное тельце жгутика, а вторая сохраняется интактной. Однако у мыши и других грызунов (в отличие от остальных изученных млекопитающих), а также у улиток деградируют и обе центриоли сперматозоидов. После оплодотворения новые центриоли возникают в зиготе либо за счет удвоения центриоли, внесенной сперматозоидом, либо за счет образования заново.
Клеточный центр — это органоид эукариотических клеток.
По-другому клеточный центр называется центросомой. В большинстве клеток центросома включает две центриоли. Однако в клетках высших растений и некоторых других организмов клеточный центр есть, а центриолей (или центросомы) нет.
Обычно в неделящейся клетке бывает только одна центросома, и находится она в центральной ее области.
Центриоль — немембранный органоид.
Каждая центриоль состоит из девяти триплетов микротрубочек, которые образует белок тубулин. Триплеты соединены между собой таким образом, что создается цилиндр. Высота цилиндра относится к его диаметру как 3 : 1. Средняя высота составляет около 0,3 мкм, а диаметр — около 0,1 мкм. Две центриоли располагаются под углом 90° друг к другу.
Однако строение клеточного центра несколько сложнее.
Кроме пары центриолей в нем образуется сеть волокон и отходящих микротрубочек. Причем одна из центриолей является материнской и именно на ней формируются дополнительные образования.
Основная функция клеточного центра — это организация веретена деления. У животных и многих грибных клеток в процессе клеточного деления центриоли центросомы расходятся к различным полюсам клетки. Около каждой путем самосборки из тубулина образуется парная дочерняя центриоль (или она образуется позже, после деления).
Таким образом, в клетке оказывается два клеточных центра. От каждого в направлении к центру, к хромосомам, осуществляется сборка микротрубочек. Микротрубочки прикрепляются к центромерам хромосом и обеспечивают их равноценное расхождение к полюсам, или обеспечивают расхождение хроматид путем их отрыва друг от друга.
При расхождении происходит разборка микротрубочек с так называемого минус-конца, который находится в клеточном центре.
Трубочка уменьшается и тем самым притягивает хромосому к своему полюсу клетки.
У растений веретено деления образуется без участия центриолей.
Кроме образования веретена деления клеточный центр выполняет и другие функции.
В нем образуются микротрубочки для поддержания структуры клетки, базальные тельца ресничек и жгутиков.
Строение и роль центриолей
Центриоли — немембранные органоиды эукариотических клеток, причем их нет в клетках высших растений, ряда грибов и некоторых животных.

Каждая центриоль состоит из девяти триплетов тубулиновых микротрубочек.
Триплеты располагаются по окружности цилиндра длиной около 0,3 мкм и диаметром около 0,1 мкм.

В каждом триплете микротрубочки отличаются.
Одна из них состоит из большего числа протофиламентов, а две другие представляют собой как бы полусферы, присоединенные вторая к первой, а третья ко второй.
В паре центриоли располагаются под прямым углом друг к другу. В интерфазе находятся в центре клетки и связаны либо с ядром, либо с комплексом Гольджи.

Клеточный центр является главным центром организации микротрубочек, инициирует их рост.
Здесь же образуются жгутики и реснички.
Клеточный центр выполняет функцию организации веретена деления. Центриолей нет у растений, но веретено у них образуется. Поэтому считается, что веретено образует именно клеточный центр, а не входящие в его состав центриоли. Вероятная функция центриолей — ориентация веретена так, чтобы хромосомы расходились именно к полюсам.
Перед делением каждая центриоль из пары отходит к своему полюсу.
От центриолей, находящихся на полюсах, вырастают микротрубочки. Они прикрепляются к центромерам хромосом и обеспечивают равноценное распределение наследственного материала между дочерними клетками.
В новых клетках возле каждой центриоли возникает новая – дочерняя.
Однако бывают другие варианты: вторая центриоль пары может появляться раньше, или в клетке может быть несколько пар. Кроме того, центриоли образуют базальные тельца, представляющие собой их видоизменения, находящиеся у основания жгутиков и ресничек.
Вопрос 1. Каковы функции клеточного центра?
Клеточный центр выполняет функцию формирования внутреннего скелета клетки (цитоскелета). Цитоскелет представляет собой сеть микротрубочек, пронизывающих цитоплазму, поддерживающих форму клетки, обеспечивающих движение органоидов клетки, а также работу специализированных органоидов движения — ресничек и жгутиков.
Клеточный центр обеспечивает также и нормальное деление клетки.
Центриоли клеточного центра расходятся к полюсам делящейся клетки и образуют веретено деления, благодаря которому из одной материнской впоследствии образуются две дочерние клетки.
Центриоли представлены цилиндриками, образованными множеством микротрубочек.
Центриоли, расположенные под прямым углом друг относительно друга, находятся вблизи от ядра и образуют клеточный центр.
Вопрос 2. Каковы функции центриолей в клетке?
Центриоли входят в состав клеточного центра и обеспечивают нормальное деление клетки. Перед ее делением центриоли расходятся к полюсам, образуя веретено деления клетки.
Вопрос 3. В чем сходство и различие между ресничками и жгутиками?
У органоидов движения клетки много общего. Реснички и жгутики являются специализированными органоидами движения клетки, они образованы микротрубочками.
В основании и жгутика, и реснички лежит базальное тельце, которое укрепляет их в цитоплазме клетки. Механизм движения ресничек и жгутиков одинаков, в его основе лежит скольжение микротрубочек друг относительно друга. Сходство этих органоидов движения заключается также и в том, что на их работу расходуется энергия АТФ.
Различаются реснички и жгутики размерами. Жгутики в несколько раз длиннее ресничек.
Кроме того, реснички, изгибаясь волнообразно, обеспечивают клетке плавное, медленное передвижение. Жгутик же осуществляет вращательные движения, что позволяет клетке активно перемещаться.
Вопрос 4. Назовите примеры клеточных включений.
Временные образования в клетке называют клеточными включениями. К ним относятся гранулы крахмала, гликогена или белка, мелкие капли жира, кристаллы солей.
Центриоли (от лат. centrum – срединная точка, центр)представляют два перпендикулярно расположенных друг к другу цилиндра, стенки которых образованы микротрубочками и соединены системой связок.
Конец одного цилиндра (дочерняя центриоль) направлен к поверхности другого (материнская центриоль). Совокупность сближенных между собой материнской и дочерней центриолей называетя диплосомой.
Впервые центриоли были обнаружены и описаны в 1875 В. Флемингом. В интерфазных клетках центриоли часто располагаются возле комплекса Гольджи и ядра.
Ультрамикроскопическое строение центриолей было изучено только с помощью электронного микроскопа. Стенку центриолей составляют расположенные по окружности 9 триплетов микротрубочек, образующих полый цилиндр.
Центриолярный цикл
Строение и активность центриолей меняются в зависимости от периода клеточного цикла.
Это позволяет говорить о центриолярном цикле. В начале периода G1 от поверхности материнской центриоли начинается рост микротрубочек, которые растут и заполняют цитоплазму. По мере роста микротрубочки теряют связь с областью центриолей и могут находиться в цитоплазме длительное время. В периоде S или G2 происходит удвоение числа центриолей.
Этот процесс заключается в том, что центриоли в диплосоме расходятся и около каждой из них происходит закладка процентриолей. В начале вблизи и перпендикулярно исходной центриоли закладываются девять одиночных микротрубочек. Затем они преобразуются в девять дуплетов, а потом в девять триплетов микротрубочек новых центриолей.
Этот способ увеличения числа центриолей был назван дупликацией. Следует отметить, что удвоение числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования процентриолей.
Таким образом, в результате дупликации в клетке содержатся четыре попарно связанные центриоли. В этом периоде материнская центриоль продолжает играть роль центра образования цитоплазматических микротрубочек.
Центриоли, их строение и поведение в клеточном цикле
В периоде G2 обе материнские центриоли покрываются фибриллярным гало (зона тонких фибрилл), от которого в профазе начнут отрастать митотические микротрубочки.
В этом периоде в цитоплазме происходит исчезновение микротрубочек и клетка стремиться приобрести шаровидную форму. В профазе митоза диплосомы расходятся к противоположным полюсам клетки. От фибриллярного гало материнской центриоли отходят микротрубочки, из которых формируется веретено деления митотического аппарата.
Таким образом, центриоли являются центрами организации роста микротрубочек. В телофазе происходит разрушение веретена деления.
Следует отметить, что в клетках высших растений, некоторых водорослей, грибов, ряда простейших центры организации роста микротрубочек центриолей не имеют.У некоторых простейших центрами индукции образования микротрубочек выступают плотные пластинки, связанные с мембраной.
Цитоплазма клетки представляет собой вязкую жидкость, поэтому из-за поверхностного натяжения клетка должна иметь шаровидную форму.
Однако помимо шаровидной встречается множество других форм клеток (кубические, призматические, звездчатые, дисковидные, с разнообразными отростками и другие). Форма определяется с помощью жестких, параллельно расположенных волокон. Эти волокна называются фибриллярными структурами цитоплазмы.
К ним относятся микротрубочки, микрофиламенты и промежуточные филаменты. Эти структуры образуют цитоскелет клетки (опорно-двигательная система).Цитоскелет определяет форму клетки, участвует в передвижении клетки, во внутриклеточном транспорте органоидов и отдельных соединений.
Микротрубочки — немембранные органоиды, представляющие собой полые цилиндры длиной около 200 нм и толщиной около 25 нм.
Микротрубочки можно обнаружить в цитоплазме практически всех эукариотных клеток. В больших количествах они находятся в цитоплазматических отростках нервных клеток, фибробластов и других изменяющих свою форму клеток. Местом организации роста микротрубочек цитоскелета в интерфазной клетке является центриоль.
Микротрубочки различного происхождения (реснички простейших, клетки нервной ткани, веретено деления) имеют сходный состав и содержат белки — тубулины.
Очищенные тубулины при определенных условиях способны собираться в микротрубочки. Добавление алкалоида колхицина предотвращает самосборку микротрубочек или приводит к разборке уже существующих.
клетке тубулины существуют в двух формах — свободной и связанной. Сдвиг равновесия между этими формами приводит или к диссоциации микротрубочек, или к их росту. Ни тубулины в чистом виде, ни построенные из них микротрубочки не способны к сокращению.
В клетках микротрубочки принимают участие в создании ряда временных (цитоскелет интерфазных клеток, веретено деления) или постоянных структур (центриоли, базальные тельца, реснички, жгутики). Кроме того, микротрубочки способствуют ориентированному движению внутриклеточных компонентов.
Микрофиламенты представляют собой тонкие (около 7 нм) белковые нити, встречающиеся практически во всех типах эукариотических клеток.
Они расположены пучками или слоями в кортикальном слое цитоплазмы, непосредственно под плазмолеммой. Сеть микрофиламентов выявлена в большинстве клеток. В состав микрофиламентов входят сократительные белки: актин, миозин, тропомиозин, альфа-актинин.
Функции микрофиламентов заключаются в образовании цитоскелета и обеспечении большинства внутриклеточных форм движения (токи цитоплазмы, движение вакуолей, митохондрий).
Промежуточные филаменты имеют толщину около 10 нм и также являются белковыми структурами. Это тонкие неветвящиеся, часто располагающиеся пучками нити.
Характерно, что их белковый состав различен в разных тканях. В эпителии в состав промежуточных филаментов входит кератин. Пучки кератиновых промежуточных филаментов в эпителиальных клетках образуют тонофибриллы.
В состав промежуточных филаментов клеток мезенхимальных тканей (например, фибробластов) входит другой белок — виментин, в мышечные клетки — десмин, в нервных клетках в состав их нейрофиламентов также входит особый белок.
С момента появления клетки и до ее смерти в результате апоптоза (программируемой клеточной гибели) непрерывно продолжается жизненный цикл клетки.
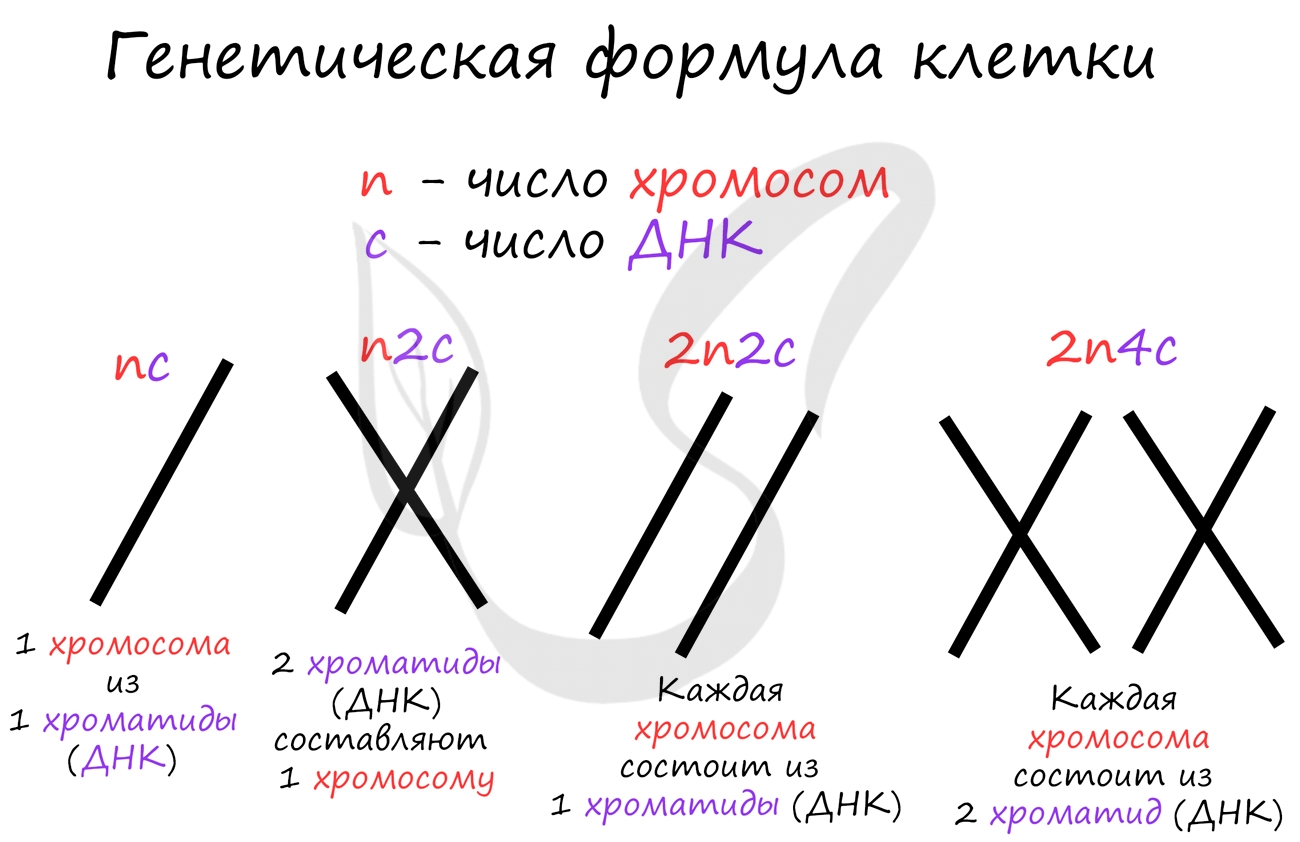
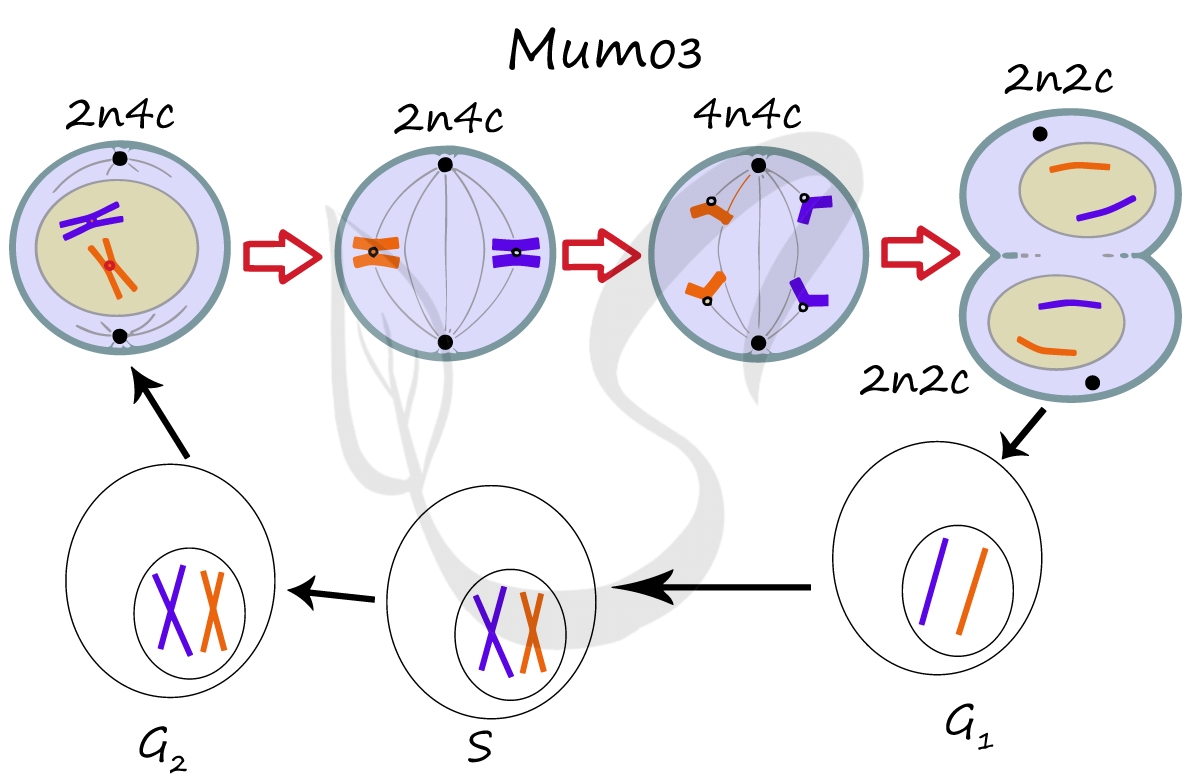
Здесь и в дальнейшем мы будем пользоваться генетической формулой клетки, где "n" - число хромосом, а "c" - число ДНК (хроматид). Напомню, что в состав каждой хромосомы может входить как одна молекула ДНК (одна хроматида) (nc), либо две (n2c).
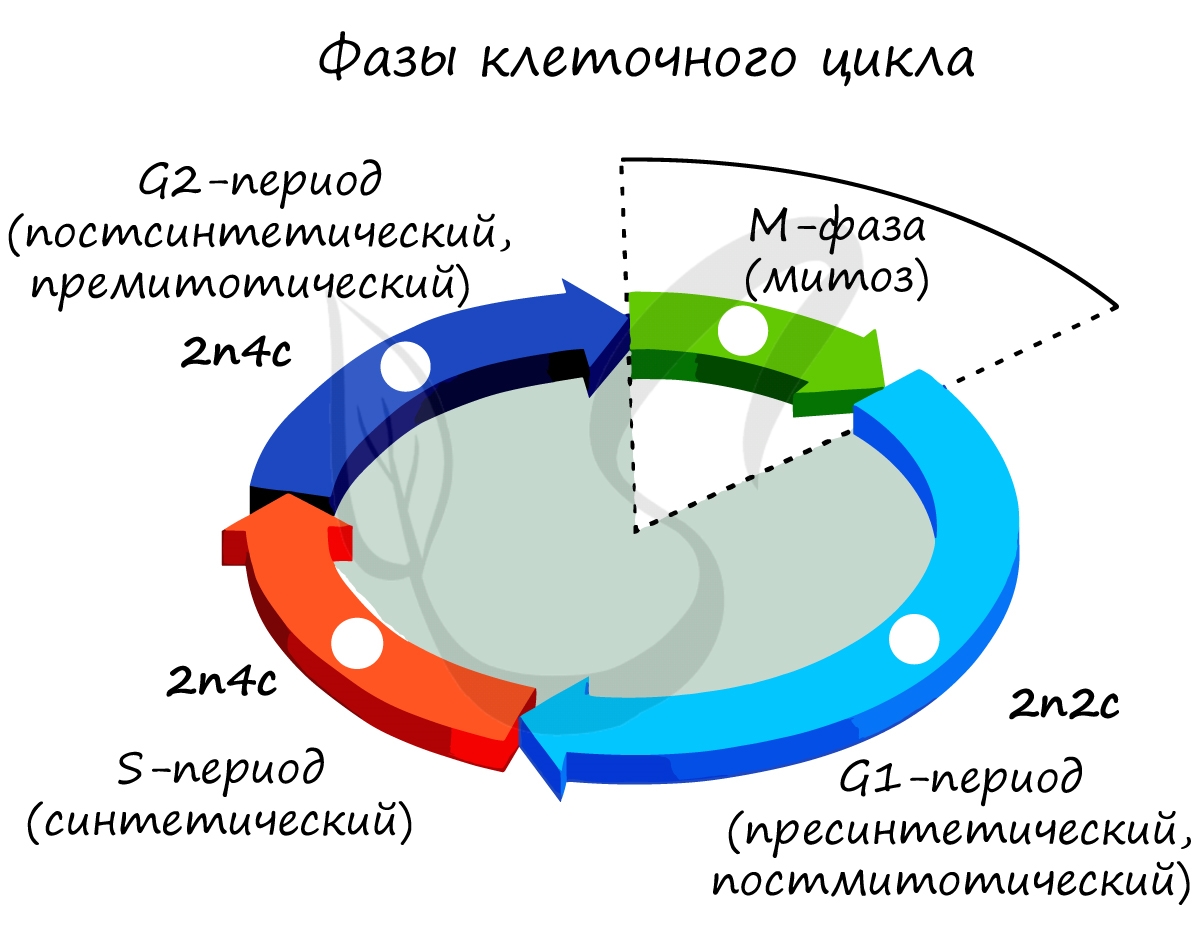
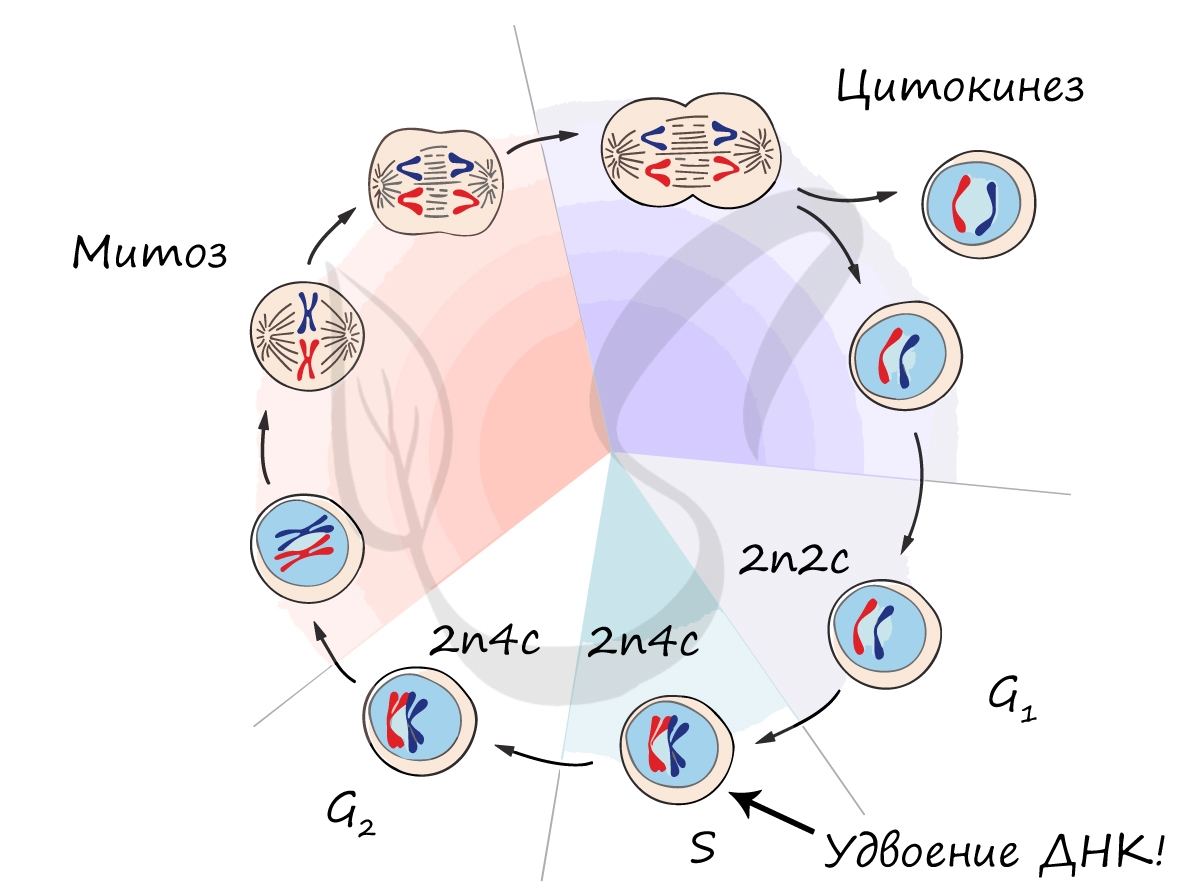
Клеточный цикл включает в себя несколько этапов: деление (митоз), постмитотический (пресинтетический), синтетический, постсинтетический (премитотический) период. Три последних периода составляют интерфазу - подготовку к делению клетки.
-
Пресинтетический (постмитотический) период G1 - 2n2c
Интенсивно образуются рибосомы, синтезируется АТФ и все виды РНК, ферменты, клетка растет.
Длится 6-10 часов. Важнейшее событие этого периода - удвоение ДНК, вследствие которого к концу синтетического периода каждая хромосома состоит из двух хроматид. Активно синтезируются структурные белки ДНК - гистоны.
Короткий, длится 2-6 часов. Это время клетка тратит на подготовку к последующему процессу - делению клетки, синтезируются белки и АТФ, удваиваются центриоли, делятся митохондрии и хлоропласты.
Митоз (греч. μίτος - нить)
Митоз является непрямым способом деления клетки, наиболее распространенным среди эукариотических организмов. По продолжительности занимает около 1 часа. К митозу клетка готовится в период интерфазы путем синтеза белков, АТФ и удвоения молекулы ДНК в синтетическом периоде.
Митоз состоит из 4 фаз, которые мы далее детально рассмотрим: профаза, метафаза, анафаза, телофаза. Напомню, что клетка вступает в митоз с уже удвоенным (в синтетическом периоде) количеством ДНК. Мы рассмотрим митоз на примере клетки с набором хромосом и ДНК 2n4c.
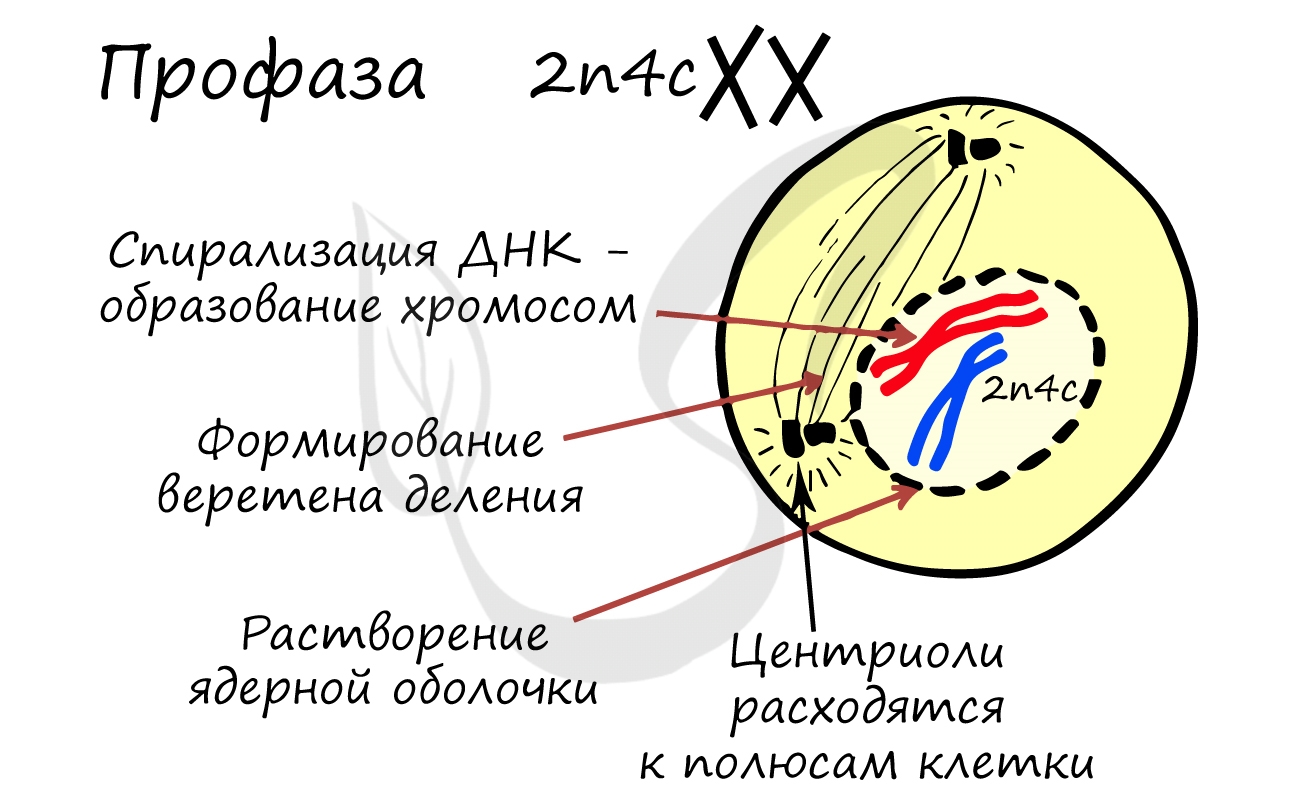
- Бесформенный хроматин в ядре начинает собираться в четкие оформленные структуры - хромосомы - происходит это за счет спирализации ДНК (вспомните мой пример ассоциации хромосомы с мотком ниток)
- Оболочка ядра распадается, хромосомы оказываются в цитоплазме клетки
- Центриоли перемещаются к полюсам клетки, образуются центры веретена деления
ДНК максимально спирализована в хромосомы, которые располагаются на экваторе клетки. Каждая хромосома состоит из двух хроматид, соединенных центромерой (кинетохором). Нити веретена деления прикрепляются к центромерам хромосом (если точнее, прикрепляются к кинетохору центромеры).
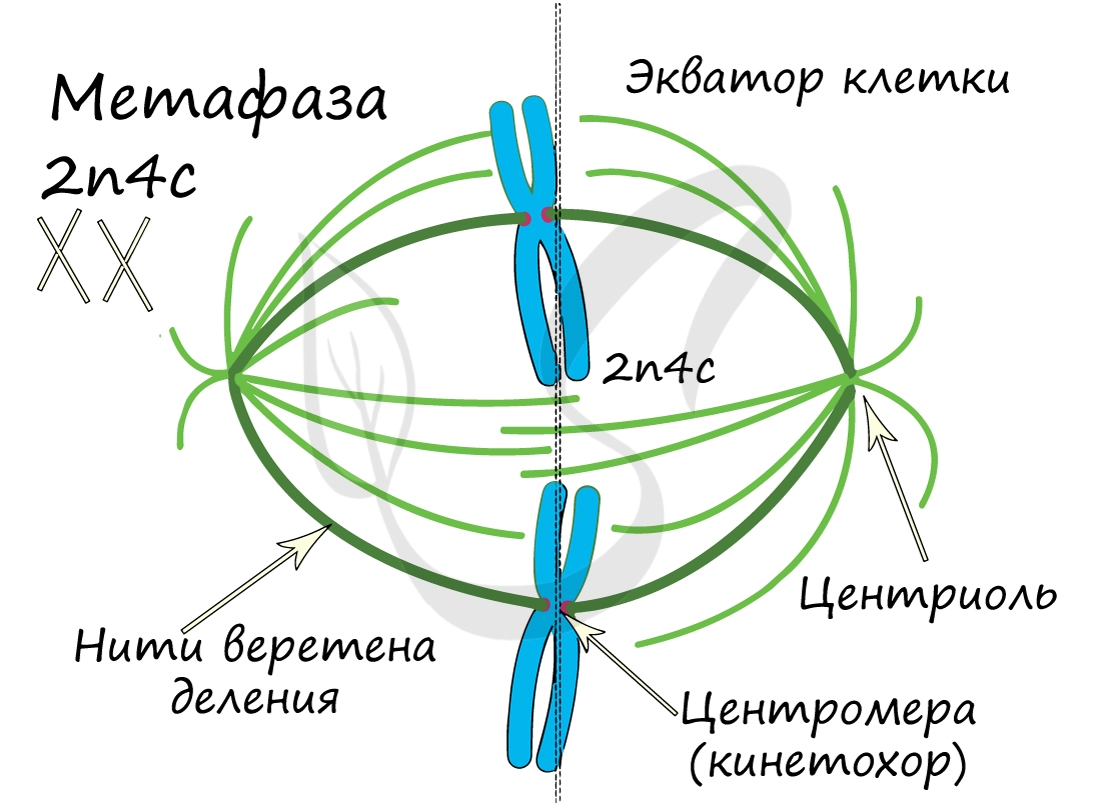
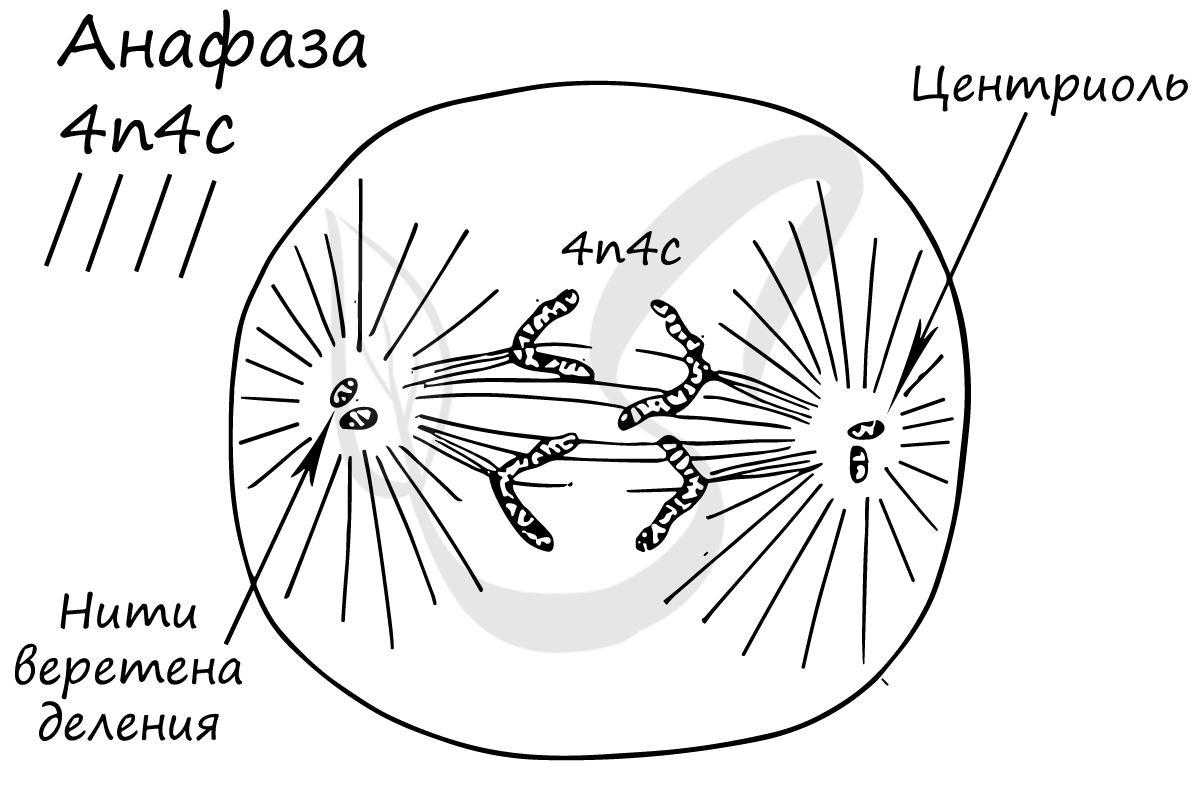
Самая короткая фаза митоза. Хромосомы, состоящие из двух хроматид, распадаются на отдельные хроматиды. Нити веретена деления тянут хроматиды (синоним - дочерние хромосомы) к полюсам клетки.
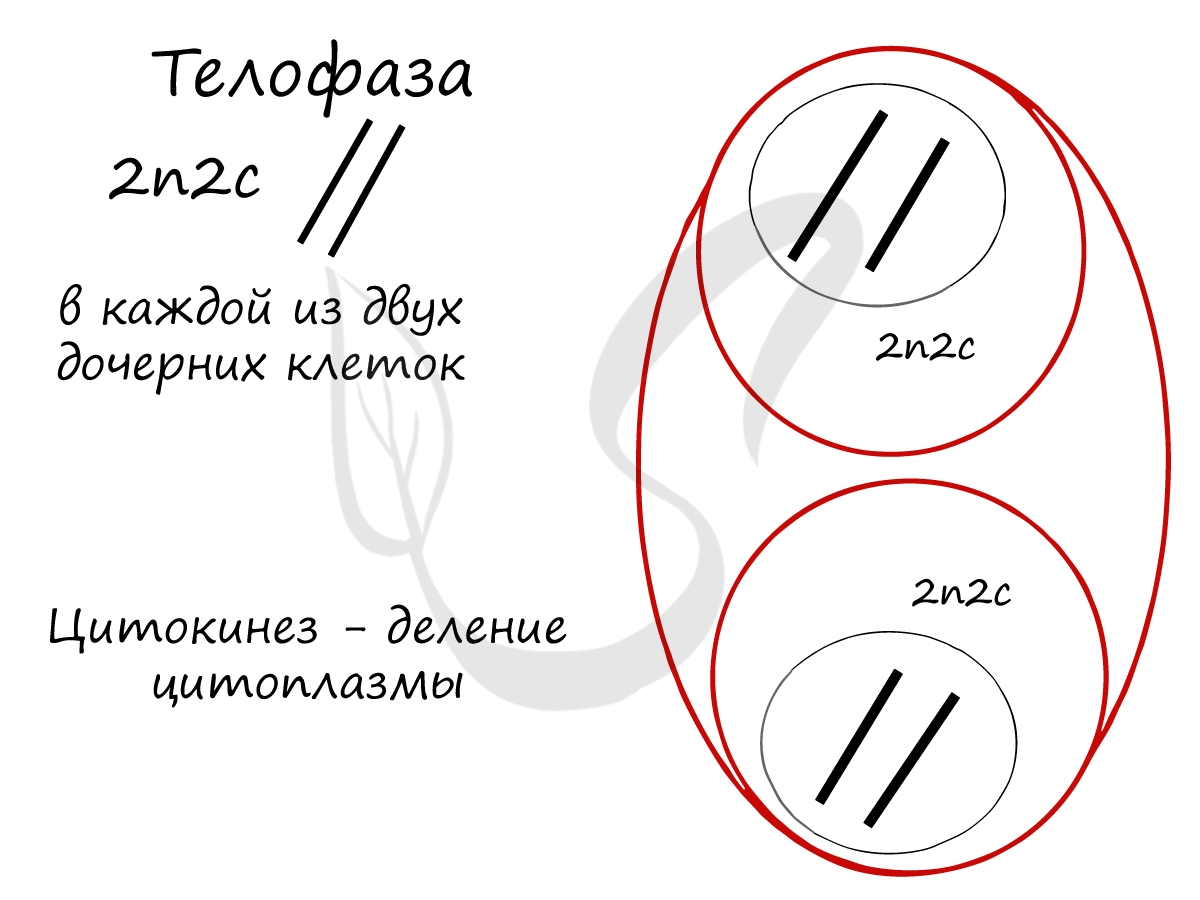
- Начинается процесс деспирализации ДНК, хромосомы исчезают и становятся хроматином (вспомните ассоциацию про раскрученный моток ниток)
- Появляется ядерная оболочка, формируется ядро
- Разрушаются нити веретена деления
В телофазе происходит деление цитоплазмы - цитокинез (цитотомия), в результате которого образуются две дочерние клетки с набором 2n2c. В клетках животных цитокинез осуществляется стягиванием цитоплазмы, в клетках растений - формированием плотной клеточной стенки (которая растет изнутри кнаружи).
Образовавшиеся в телофазе дочерние клетки 2n2c вступают в постмитотический период. Затем в синтетический период, где происходит удвоение ДНК, после чего каждая хромосома состоит из двух хроматид - 2n4c. Клетка с набором 2n4c и попадает в профазу митоза. Так замыкается клеточный цикл.
- В результате митоза образуются дочерние клетки - генетические копии (клоны) материнской.
- Митоз является универсальным способом бесполого размножения, регенерации и протекает одинаково у всех эукариот (ядерных организмов).
- Универсальность митоза служит очередным доказательством единства всего органического мира.
Попробуйте самостоятельно вспомнить фазы митоза и описать события, которые в них происходят. Особенное внимание уделите состоянию хромосом, подчеркните сколько в них содержится молекул ДНК (хроматид).
Мейоз
Мейоз (от греч. μείωσις — уменьшение), или редукционное деление клетки - способ деления клетки, при котором наследственный материал в них (число хромосом) уменьшается вдвое. Мейоз происходит в ходе образования половых клеток (гамет) у животных и спор у растений.
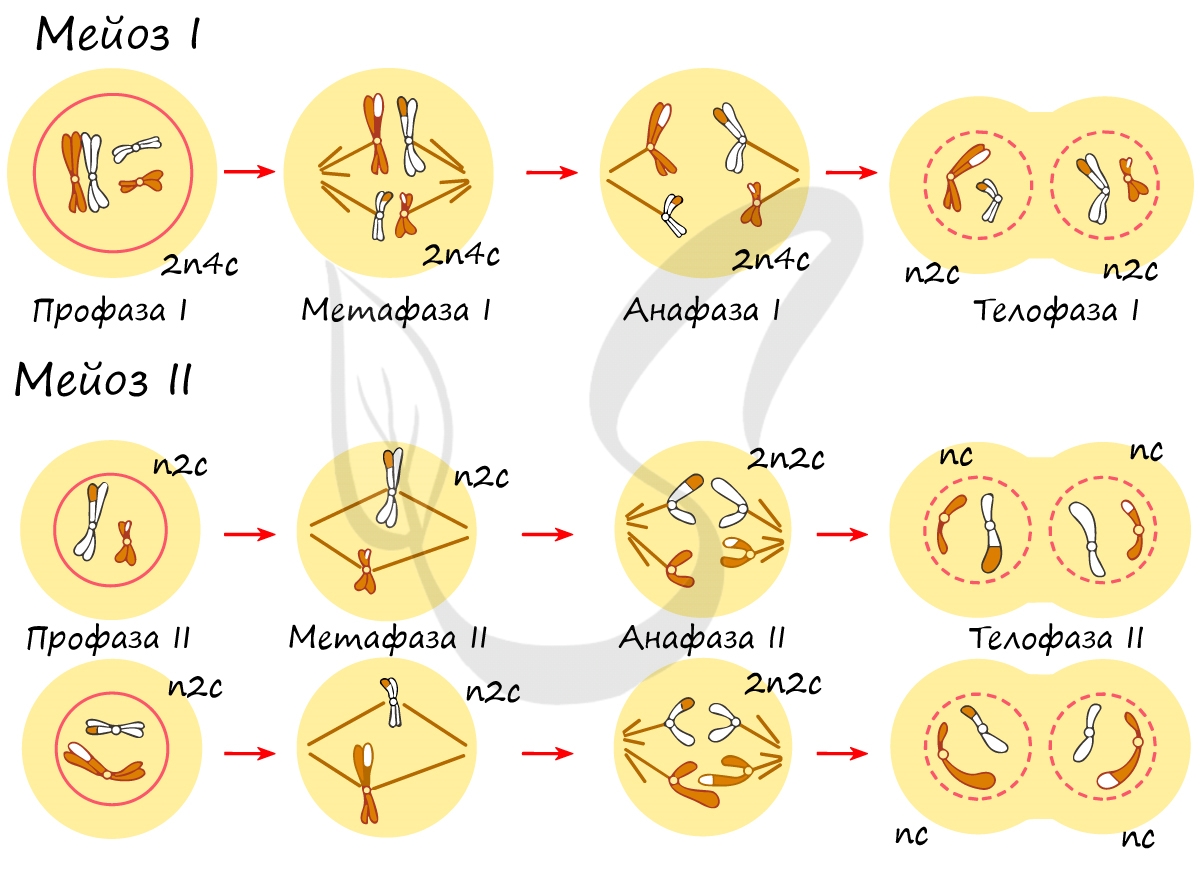
В результате мейоза из диплоидных клеток (2n) получаются гаплоидные (n). Мейоз состоит из двух последовательных делений, между которыми практически отсутствует пауза. Удвоение ДНК перед мейозом происходит в синтетическом периоде интерфазы (как и при митозе).
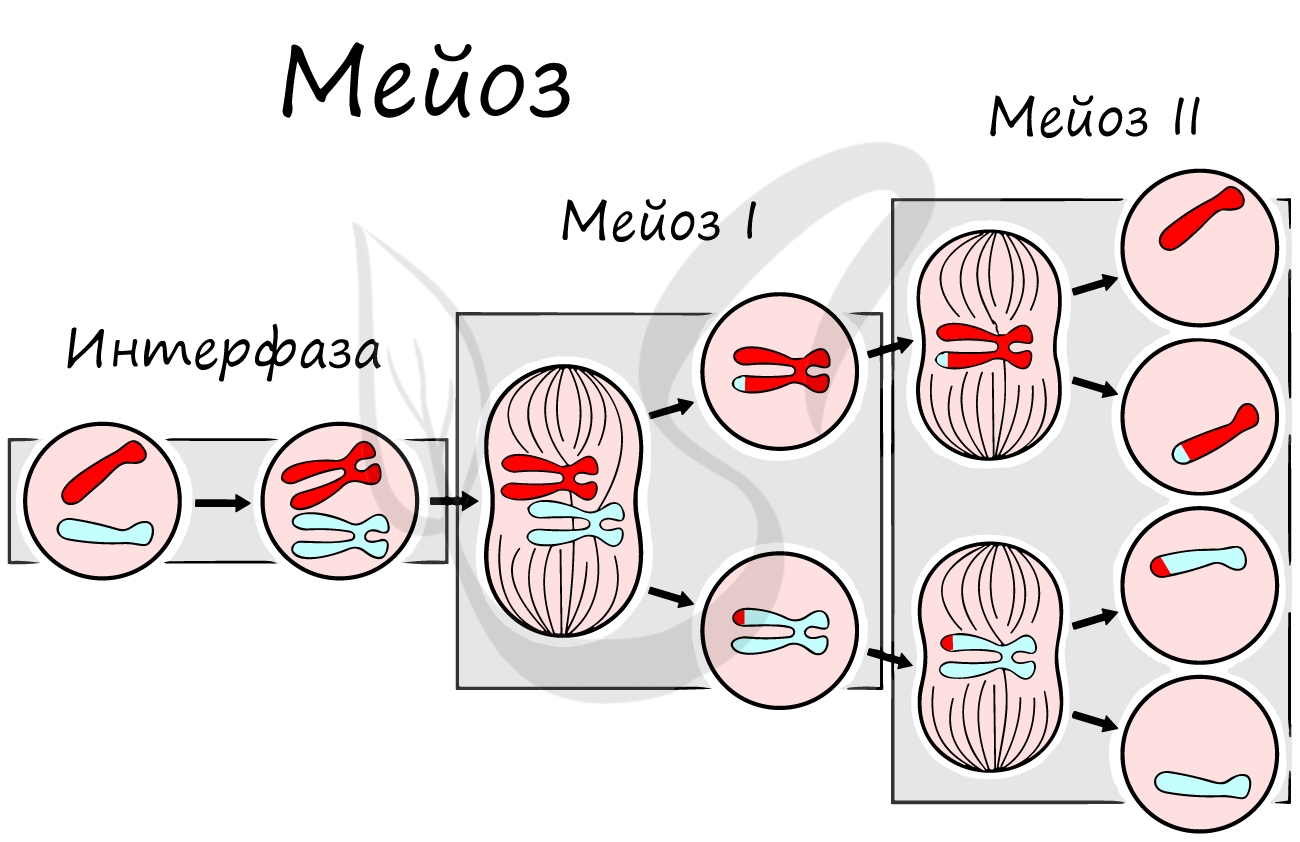
Как уже было сказано, мейоз состоит из двух делений: мейоза I (редукционного) и мейоза II (эквационного). Первое деление называют редукционным (лат. reductio - уменьшение), так как к его окончанию число хромосом уменьшается вдвое. Второе деление - эквационное (лат. aequatio — уравнивание) очень похоже на митоз.
-
Профаза мейоза I
Помимо типичных для профазы процессов (спирализация ДНК в хромосомы, разрушение ядерной оболочки, движение центриолей к полюсам клетки) в профазе мейоза I происходят два важнейших процесса: конъюгация и кроссинговер.
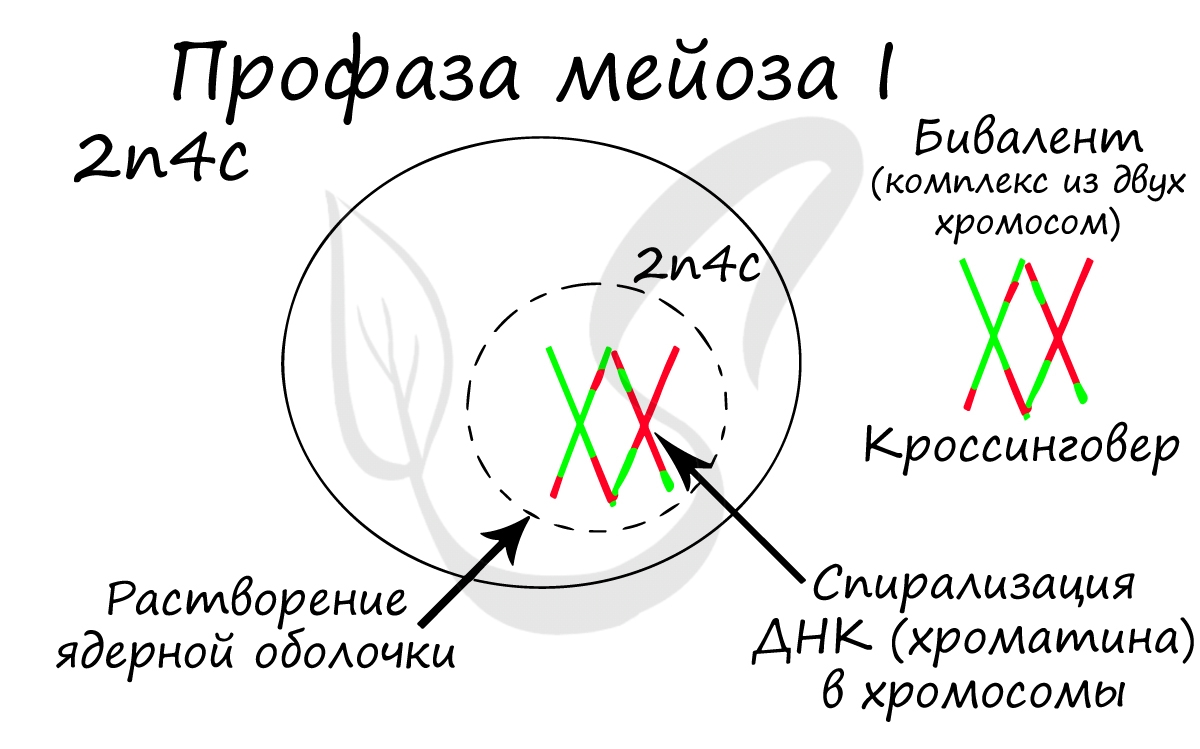
Конъюгация (лат. conjugatio — соединение) - сближение гомологичных хромосом друг с другом. Гомологичными хромосомами называются такие, которые соответствуют друг другу по размерам, форме и строению. В результате конъюгации образуются комплексы, состоящие из двух хромосом - биваленты (лат. bi - двойной и valens - сильный).
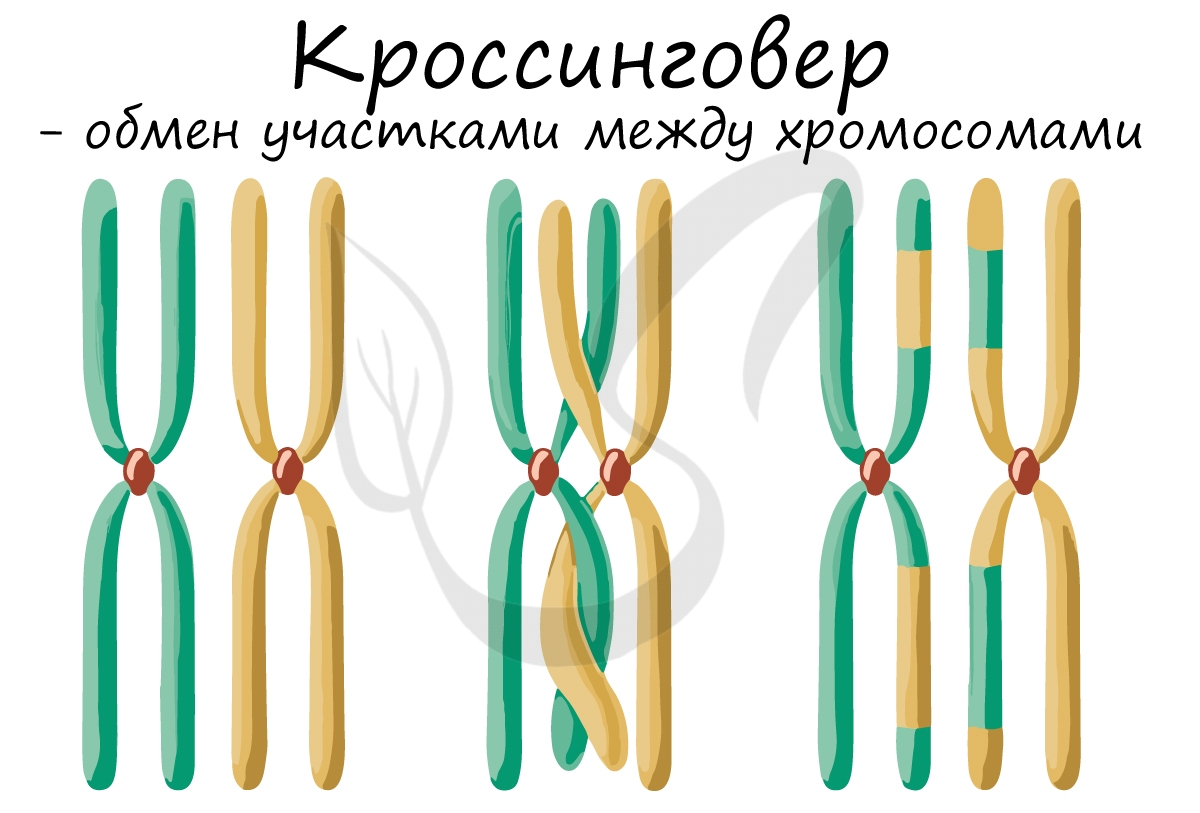
После конъюгации становится возможен следующий процесс - кроссинговер (от англ. crossing over — пересечение), в ходе которого происходит обмен участками между гомологичными хромосомами.
Кроссинговер является важнейшим процессом, в ходе которого возникают рекомбинации генов, что создает уникальный материал для эволюции, последующего естественного отбора. Кроссинговер приводит к генетическому разнообразию потомства.
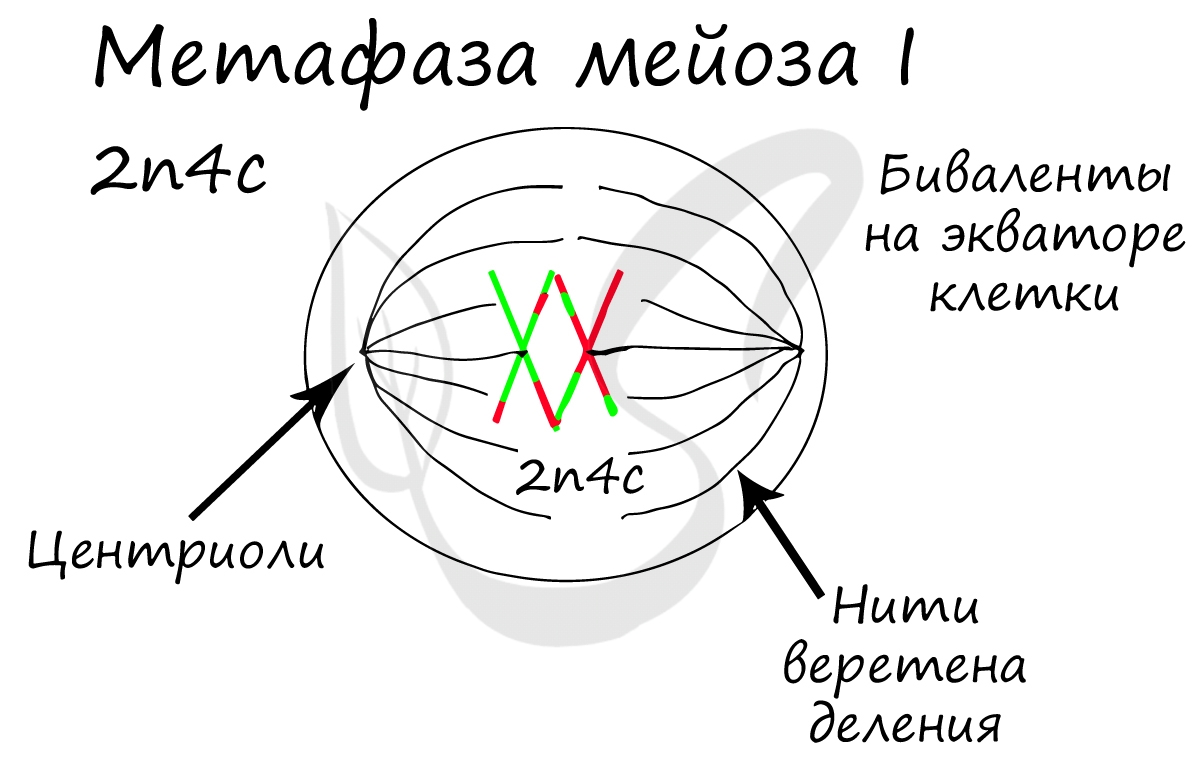
Биваленты (комплексы из двух хромосом) выстраиваются по экватору клетки. Формируется веретено деления, нити которого крепятся к центромере (кинетохору) каждой хромосомы, составляющей бивалент.
Нити веретена деления сокращаются, вследствие чего биваленты распадаются на отдельные хромосомы, которые и притягиваются к полюсам клетки. В результате у каждого полюса формируется гаплоидный набор будущей клетки - n2c, за счет чего мейоз I и называется редукционным делением.

Происходит цитокинез - деление цитоплазмы. Формируются две клетки с гаплоидным набором хромосом. Очень короткая интерфаза после мейоза I сменяется новым делением - мейозом II.
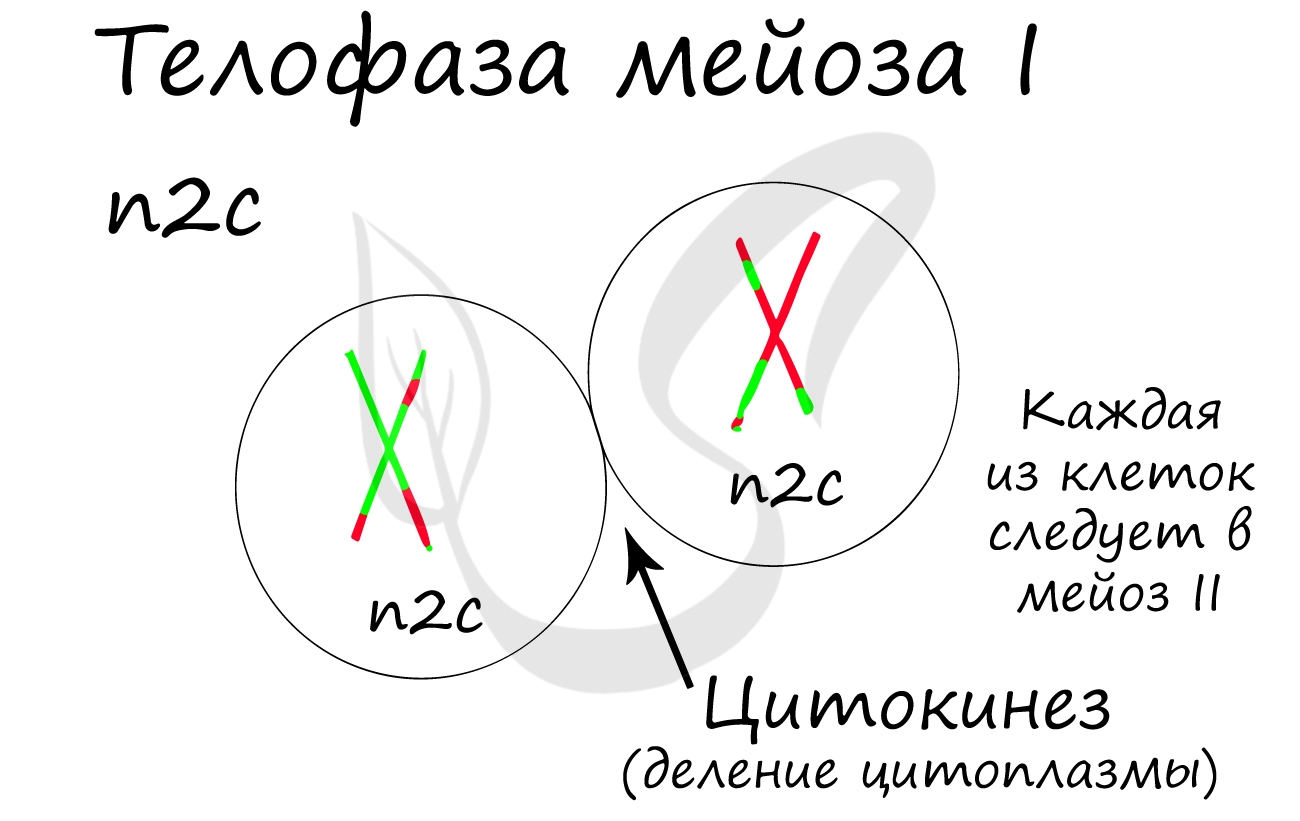
Мейоз II весьма напоминает митоз по всем фазам, поэтому если вы что-то подзабыли: поищите в теме про митоз. Главное отличие мейоза II от мейоза I в том, что в анафазе мейоза II к полюсам клетки расходятся не хромосомы, а хроматиды (дочерние хромосомы).
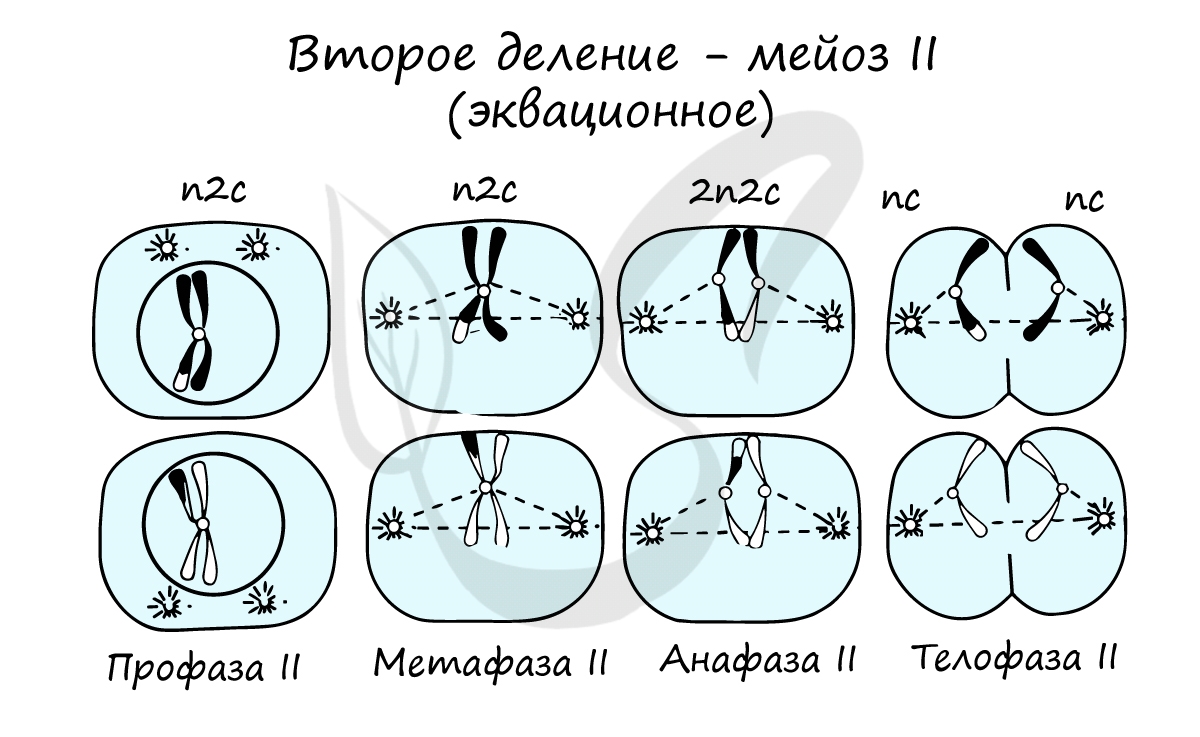
В результате мейоза I и мейоза II мы получили из диплоидной клетки 2n4c гаплоидную клетку - nc. В этом и состоит сущность мейоза - образование гаплоидных (половых) клеток. Вспомнить набор хромосом и ДНК в различных фазах мейоза нам еще предстоит, когда будем изучать гаметогенез, в результате которого образуются сперматозоиды и яйцеклетки - половые клетки (гаметы).
Сейчас мы возьмем клетку, в которой 4 хромосомы. Попытайтесь самостоятельно описать фазы и этапы, через которые она пройдет в ходе мейоза. Проговорите и осмыслите набор хромосом в каждой фазе.
Помните, что до мейоза происходит удвоение ДНК в синтетическом периоде. Из-за этого уже в начале мейоза вы видите их увеличенное число - 2n4c (4 хромосомы, 8 молекул ДНК). Я понимаю, что хочется написать 4n8c, однако это неправильная запись!) Ведь наша исходная клетка диплоидна (2n), а не тетраплоидна (4n) ;)
- Поддерживает постоянное число хромосом во всех поколениях, предотвращает удвоение числа хромосом
- Благодаря кроссинговеру возникают новые комбинации генов, обеспечивается генетическое разнообразие состава гамет
- Потомство с новыми признаками - материал для эволюции, который проходит естественный отбор
Бинарное деление надвое
Митоз и мейоз возможен только у эукариот, а как же быть прокариотам - бактериям? Они изобрели несколько другой способ и делятся бинарным делением надвое. Оно встречается не только у бактерий, но и у ряда ядерных организмов: амебы, инфузории, эвглены зеленой.
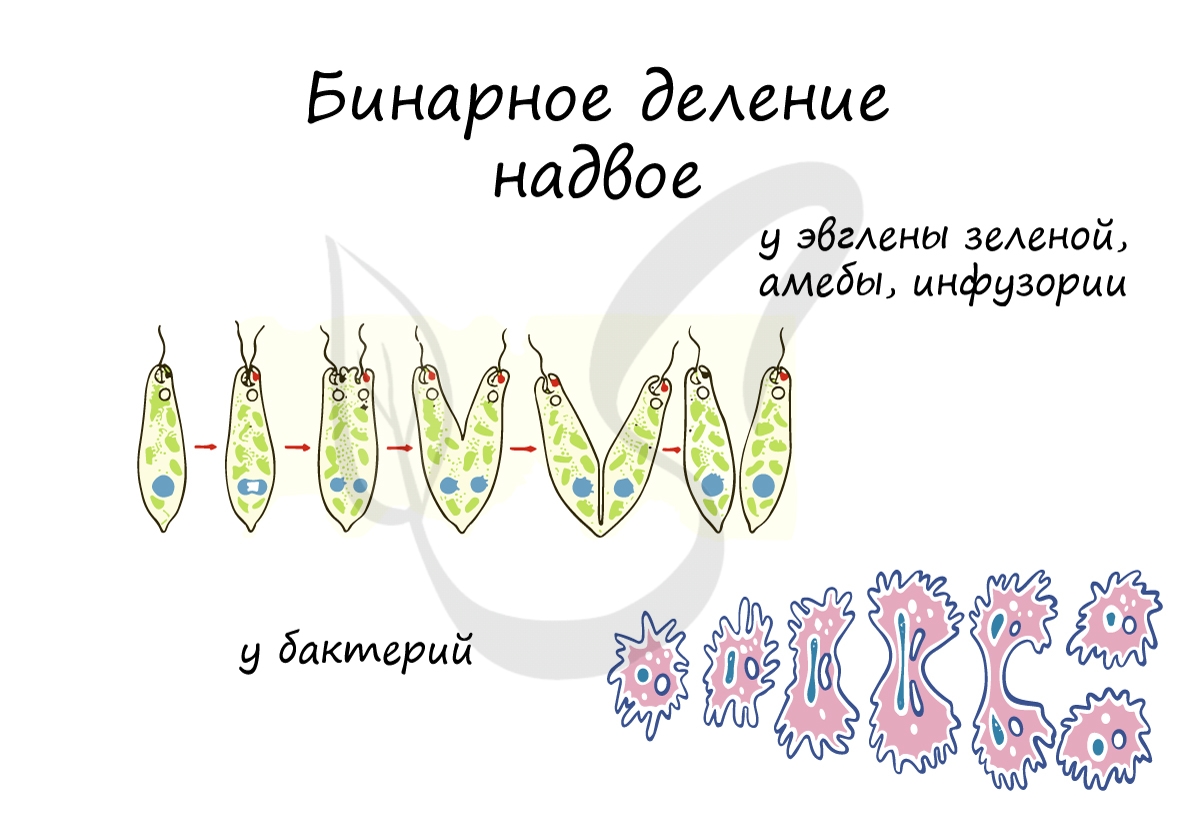
При благоприятных условиях бактерии делятся каждые 20 минут. В случае, если условия не столь благоприятны, то больше времени уходит на рост и развитие, накопление питательных веществ. Интервалы между делениями становятся длиннее.
Амитоз (от греч. ἀ - частица отрицания и μίτος - нить)
Способ прямого деления клетки, при котором не происходит образования веретена деления и равномерного распределения хромосом. Клетки делятся напрямую путем перетяжки, наследственный материал распределяется "как кому повезет" - случайным образом.
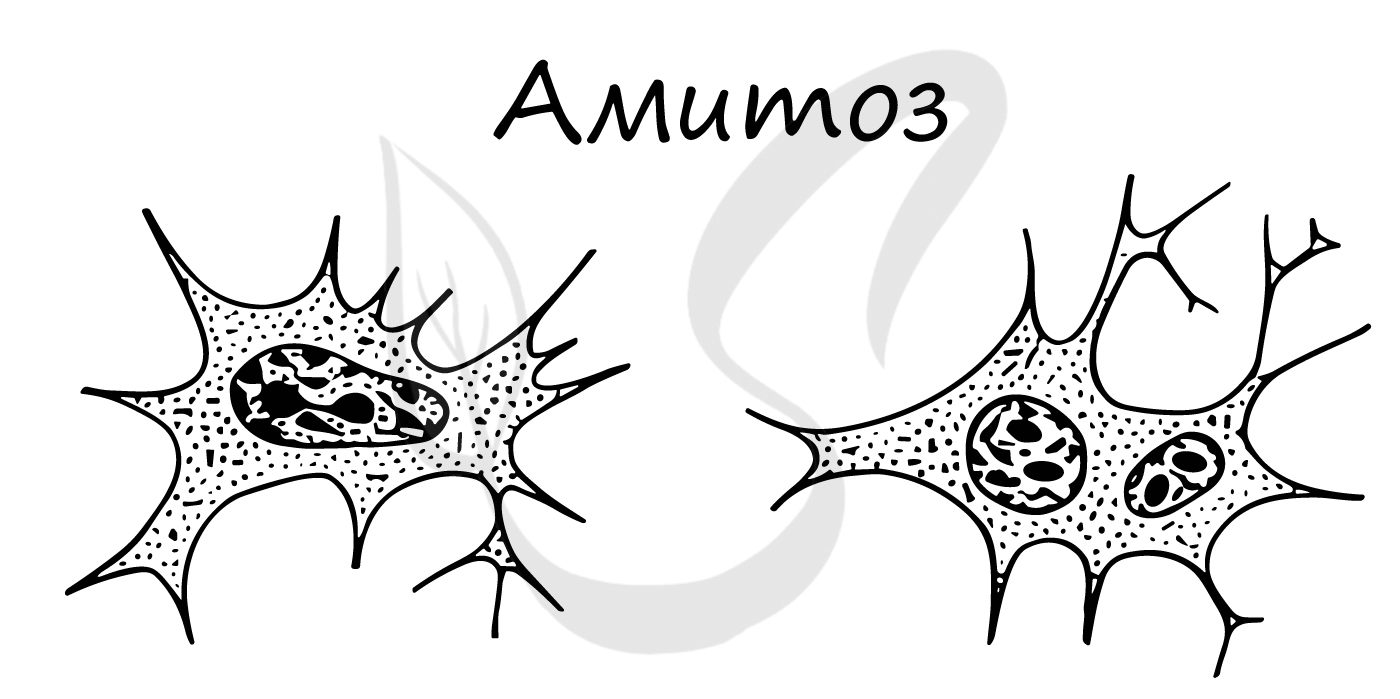
Амитоз встречается в раковых (опухолевых) клетках, воспалительно измененных, в старых клетках.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.

Центриоли встречаются в клетках животных, но отсутствуют в растительных клетках. Они помогают организовать сборку микротрубочек во время деления клеток. Центриоли расположены в клеточных структурах, известных как центросомы.
При делении клеток центросома и центриоли реплицируются и мигрируют на противоположные полюса клетки. Центриоли помогают расположить микротрубочки, которые перемещают хромосомы во время деления клеток, чтобы каждая дочерняя клетка получала соответствующее количество хромосом. Центриоли также важны для формирования клеточных структур, известных как реснички и жгутики. Эти органеллы помогают в клеточной локомоции и формируются из центриолей, называемых базальными телами.
В организмах со жгутиками и ресничками положение этих органелл определяется материнской центриолой, которая становится основным телом. Неспособность клеток использовать центриоли для создания функциональных ресничек и жгутиков связано с рядом генетических и инфекционных заболеваний.
Функции центриолей в делении клеток
Центриоли расположены за пределами, но вблизи ядра клетки. Они реплицируются во время интерфазы, до начала митоза и мейоза в клеточном цикле.
В профазе каждая центросома с центриолями мигрирует к противоположным полюсам клетки. На каждом конце клетки расположена одна пара центриолей. Митотическое веретено первоначально появляется в виде к структур, называемых астрами, которые окружают каждую пару центриолей. Микротрубочки образуют волокна веретена деления, простирающиеся от каждой центросомы, тем самым разделяя пары центриолей и удлиняя клетку.
В метафазе центриоли помогают позиционировать полярные волокна, поскольку они простираются от центросомы и располагают хромосомы вдоль метафазной пластины.
В анафазе полярные волокна, связанные с хромосомами, сокращаются и разделяют сестринские хроматиды (реплицированные хромосомы). Отделенные хромосомы вытягиваются к противоположным концам клетки полярными волокнами, простирающимися от центросомы.
В телофазе волокна веретена диспергируются по мере того, как хромосомы опираются на отдельные новые ядра. После цитокинеза каждая дочерняя клетка содержит одну центросому с одной центриольной парой.
• Центросомы расположены на полюсах веретена и участвуют в его образовании
• Центросомы участвуют в нуклеации микротрубочек и после этого часто остаются связанными с их минус-концами
• Центросомы состоят из двух центриолей, окруженных перицентриолярным материалом
• Для образования новой центросомы необходимо удвоение (дупликация) центриолей
• Дупликация центриолей контролируется клеточным циклом и скоординирована с репликацией ДНК
• Дупликация центриолей происходит в процессе образования и роста новой центриоли, которая с момента образования примыкает к существующей
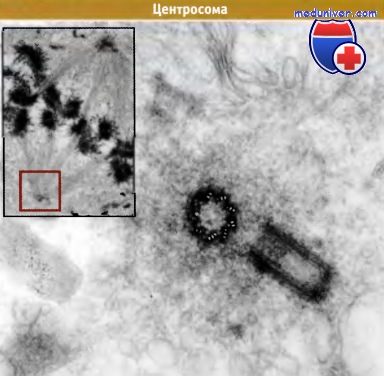
На основной фотографии помещено изображение центросомы в электронном микросокопе.
Две центриоли расположены под прямым углом друг к другу, так, что одна выглядит как круг, а другая как прямоугольник.
Вокруг первой центриоли находится скопление гранулярного материала
(сравните область, примыкающую к центриоли, с более удаленными частями цитоплазмы,
которые окрашены менее интенсивно и где заметно присутствие многих мембранных везикул).
Когда клетка приближается к митозу, в ней происходит ряд изменений. Для клеток млекопитающих наиболее заметные изменения заключаются в исчезновении из цитоплазмы длинных пучков микротрубочек и в появлении двух лучистых астральных структур, состоящих из более коротких микротрубочек, которые часто называют звездами. Этот процесс показан на рисунке ниже. Каждая из радиальных лучистых структур окружает центросому. При расхождении центросом из этих структур в клетках млекопитающих образуется веретено.
Поскольку две центросомы образуют полюса веретена, крайне важно, чтобы в момент митозе в клетке находились только две центросомы. Когда обе астральные структуры начинают взаимодействовать при образовании веретена, его структура стабилизируется хромосомами. Основную роль при этом играют их кинетохоры.
При образовании клетки каждая содержит одну центросому, которая достается ей от одного полюса веретена при предыдущем делении. В неделящихся клетках эта маленькая органелла обычно расположена поблизости от клеточного центра, где она тесно связана с ядром. В интерфазе центросома действует как центр организации микротрубочек. С его участием в цитоплазме создается сеть микротрубочек, которая пронизывает клетку. Эта сеть участвует в организации цитоплазмы и в транспорте различных материалов и органелл внутри клетки.
Центросомы образуют сеть микротрубочек, действуя как сайты их нуклеации. В пределах центросомы микротрубочка начинает расти из кольцеобразного комплекса, содержащего один из типов тубулина, называемый у-тубулином. После того как нуклеация завершилась, минус-конец микротрубочки обычно остается связанным с центросомой. Затем микротрубочка начинает удлиняться или укорачиваться за счет добавления или удаления молекул тубулина к плюс-концу, который расположен на некотором расстоянии от центросомы. В течение разных периодов времени микротрубочки остаются связанными с центросомой и в некоторых клетках отделяются от центросом под действием локализованных в них ферментов.
Связь с центросомой осуществляется посредством нескольких структурных белков, а также моторов, продвигающихся к минус-концу, включая цитоплазматический динеин и белки, относящиеся к группе кинезинов (HSET).
Каждая центросома, находящаяся в клетке, в митозе способна к образованию полюса веретена. Веретено обладает биполярной структурой, поскольку обычно клетки вступают в митоз, обладая двумя центросомами. Если в клетке находится более двух центросом и она вступает в митоз, то будет образовываться веретено, содержащее слишком много полюсов, и потомство таких клеток может характеризоваться анеуплоидностью. Это предотвращается специальными механизмами, которые позволяют центросоме в клеточном цикле реплицироваться только один раз.
Когда эти контрольные механизмы перестают функционировать, образуется слишком много центросом, которые могут привести к образованию генетических дефектов и к развитию раковых клеток и опухолей. Для того чтобы понять, каким образом обеспечивается биполярная структура веретена, необходимо выяснить строение центросомы и механизм контроля ее репликации.
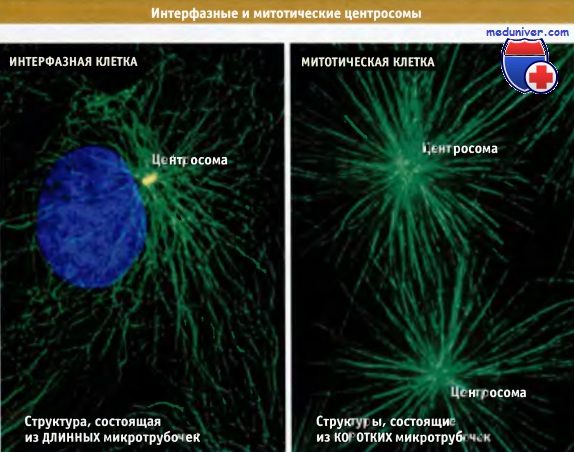
В интерфазных клетках центросома (желтая точка рядом с ядром) нуклеирует образование длинных микротрубочек, которые пронизывают цитоплазму.
В митотических клетках способность центросом к нуклеации микротрубочек возрастает,
и каждая из реплицировавшихся центросом нуклеирует плотные радиальные структуры (звезды),
состоящие из прямых коротких микротрубочек.
Микротрубочки флуоресцируют зеленым цветом, ДНК — синим.
В световом микроскопе в большинстве клеток центросомы выглядят как одна или две точки. Для того чтобы детально исследовать эти органеллы и оценить всю сложность их строения, используют электронную микроскопию (ЭМ). В электронном микроскопе видно, что центросома содержит пару структур, называемых центриоли. Каждая центриоль представляет собой круговую структуру, содержащую девять триплетов микротрубочек, распределенных равномерно по периметру цилиндра диаметром 0,3 мкм.
Еще в 1888 г. Бовери с сотрудниками пришли к выводу, что центриоль, которая была с трудом видна в их световом микроскопе, представляет собой длительно существующую и независимую органеллу, которая образуется только при делении предсуществующей центриоли. Действительно, за редким исключением, новые центриоли образуются только в связи с существующей центриолью. В некоторых клетках тесная связь между двумя центриолями сохраняется на протяжении клеточного цикла. Однако в интерфазе, во многих клетках эта связь нарушается, и две центриоли независимо перемещаются по клетке.
Каждая центриоль связана с областью диффузного материала, которая в электронном микроскопе выглядит непрозрачной. Если смотреть на центриоль со стороны торца, то непрозрачная область расположена вокруг центриоли. Этот перицентриолярный материал состоит из большого количества белков, прикрепленных к каркасу. Как правило, материнская центриоль содержит больше этого материала по сравнению с дочерней, по крайней мере до момента завершения следующего цикла репликации центриоли. Среди белков перицентриолярного материала находится несколько типов белковых моторов микротрубочек и у-тубулиновые кольца, участвующие в нуклеации микротрубочек. Сами центриоли также содержат ряд специфических структур и ферментов, некоторые из которых также присутствуют в перицентриолярном материале.
В интерфазе клеточного цикла центросомы выполняют несколько совместных функций. В большинстве клеток в фазе G1 материнская центриоль инициирует образование длинной тонкой структуры, покрытой мембраной, отходящей от поверхности клетки, которая называется первичная ресничка.
Хотя обычно на них не обращают внимания, первичные реснички можно наблюдать очень часто, и легче перечислить клетки, у которых они отсутствуют, чем те, которые ими обладают. В некоторых эпителиях эти структуры могут простираться более чем на 20 мкм от тела клетки. Поскольку эти структуры не обнаруживаются у многих трансформированных клеток, первичные реснички не играют критической роли в жизни клетки. Этот факт позволил исследователям предположить, что они просто представляют собой рудиментарные образования, подобно аппендиксу.
Однако внешние сегменты палочек и колбочек глаза — высокоспециализированные структуры, в которых происходит поглощение фотонов, образованы производными от первичных ресничек, и накапливаются данные, что первичные реснички также необходимы для процессов нормального развития и функционирования тканей.
В клетках животных количество пар центриолей определяет число центросом. Поэтому клетки контролируют число центросом, которые они содержат, за счет регулирования репликации центриолей. Только начинают появляться работы, в которых вскрываются механизмы контроля за точным удвоением центриолей, и связи этого удвоения с активностью ядра в клеточном цикле (например, с процессом репликации ДНК). В настоящее время очевидно, что наступление репликации центриолей контролируется изменениями в цитоплазме, что указывает на регулирующую роль растворимого фактора. Наряду с этим, условия, позволяющие центри-олям вступить в фазу репликации, существуют только в S-фазе клеточного цикла, когда также реплицируется ДНК.
По-видимому, основным регулятором репликации центросом является CDK2 киназа и ее активаторы циклин А и Е. Эти регуляторы активируются перед началом S-фазы и также контролируют вхождение клетки в фазу синтеза ДНК. То, что один и тот же регулятор контролирует репликацию ДНК и центриолей, обеспечивает скоординированность двух этих процессов, так что клетка входит в митоз по завешении репликации центросомы и хромосом. Хотя неизвестно, как инициируется дупликация центриолей, не вполне ясно, каким образом она ограничивается образованием одной новой центриоли из каждой предсуществующей.
Начавшись, дупликация центриолей приводит к постепенному образованию новой центриоли, примыкающей к каждой из двух, с которыми клетка вступила в S-фазу. Одна из них называется дочерней и является более молодой, поскольку образовалась в предыдущем клеточном цикле. Другая, материнская, образовалась в раннем цикле и характеризуется наличием первичной реснички. Первый признак репликации центриоли состоит в появлении двух коротких процентриолей, каждая из которых распространяется под прямым углом к стенке одной из центриолей.
Этот процесс не зависит от физического взаимодействия между двумя исходными центриолями, поскольку происходит, даже когда они разобщены. После образования процентриолей они медленно элонгируют до тех пор, пока их длина не достигнет размеров зрелой центриоли к моменту митоза. Хотя в случае материнской и дочерней центриоли процентриоли образуются и растут одинаковым образом, большая часть перицентриолярного материала остается связанным с более зрелой, материнской центриолью.
В конце концов, в процессе дупликации дочерняя центриоль накапливает новый перицентриолярный материал; частично это происходит за счет микротрубочек, которые она организует. В поздней интерфазе клетка содержит две центросомы, каждая из которых состоит из пары тесно связанных друг с другом центриолей и перицентриолярного материала. В некоторых клетках две центросомы остаются физически связанными между собой и функционируют как единое целое до момента вступления в митоз. В других клетках связь между центросомами разрушается, и они расходятся до момента вступления клетки в митоз. Время наступления момента разделения двух центросом по отношению к разрыву ядерной оболочки сильно варьирует, даже среди генетически однородных клеток, растущих в тесном контакте.
Митоз в клетке кенгуровой крысы. На левом снимке видны четыре центросомы (помечены желтыми стрелками).
Образуются три метафазных пластинки (желтые линии на центральном снимке)
и четыре группы хромосом (помечены стрелками на правом снимке).
При цитокинезе образуются четыре анеуплоидных клетки. В митотической клетке (вверху слева) центросома состоит из двух центриолей
— материнской (на этой микрофотографии представлена в поперечном сечении) и дочерней (представлена в продольной проекции)
— окруженных темной областью аморфного перицентриолярного материала.
Каждая центриоль (наверху справа) состоит из триплетов микротрубочек, образующих как бы стенку цилиндра.
На рисунке внизу видно, что центриоли расположены под прямым углом по отношению друг к другу.
Вокруг них находится много различных белков, образующих перицентриолярный материал. Электронная микрофотография основания первичной реснички.
Структура растет от материнской центриоли, которая находится под плазматической мембраной.
Между материнской и дочерней центриолями находится перицентриолярный материал плотного гранулярного вида.
Показана лишь часть клетки и первичной реснички. Центриолярный цикл в клетках млекопитающих.
Две центриоли центросомы материнской клетки дуплицируются, и затем две образующихся центросомы распределяются по дочерним клеткам.
На вставке представлены две микрофотографии реплицированных центросом до и после их расхождения в начале митоза.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Читайте также: