
Какова роль риниофитов в царстве растений кратко
Обновлено: 04.07.2024
Риниофиты - псилофиты (Rhyniophyta), самый древний и примитивный отдел вымерших растений. Некоторые водоросли стали более совершенными и приспособились жить во влажных местах суши по берегам водоемов. Начался переход некоторых растений от водного образа жизни к наземному. Это произошло ~ 350-400 млн. лет назад.
Впервые остатки Р. были найдены в 1859 канадским геологом Д. Досоном в девонских отложениях на острове Гаспе в Канаде. Р. характеризовались примитивностью строения, верхушечным расположением спорангиев, дихотомическим или псевдомоноподиальным ветвлением спорофита, отсутствием корней и листьев и равноспоровостью. Проводящая система была представлена протостелой. Ксилема центрархная. Утолщение трахеид было лестничным, спиральным или кольчатым. Механические ткани отсутствовали. Имелись просто устроенные устьица. Спорангии были толстостенные, разные по форме. В спорангиях развивались многочисленные споры. Произрастали Р. во влажных, часто заболоченных местах, образуя заросли. Р. стали предками других низкоорганизованных ныне живущих групп высших растений — моховидных, плауновидных, хвощевидных, папоротниковидных.
Риниофиты - псилофиты (Rhyniophyta), самый древний и примитивный отдел вымерших растений. Некоторые водоросли стали более совершенными и приспособились жить во влажных местах суши по берегам водоемов. Начался переход некоторых растений от водного образа жизни к наземному. Это произошло ~ 350-400 млн. лет назад.
Впервые остатки Р. были найдены в 1859 канадским геологом Д. Досоном в девонских отложениях на острове Гаспе в Канаде. Р. характеризовались примитивностью строения, верхушечным расположением спорангиев, дихотомическим или псевдомоноподиальным ветвлением спорофита, отсутствием корней и листьев и равноспоровостью. Проводящая система была представлена протостелой. Ксилема центрархная. Утолщение трахеид было лестничным, спиральным или кольчатым. Механические ткани отсутствовали. Имелись просто устроенные устьица. Спорангии были толстостенные, разные по форме. В спорангиях развивались многочисленные споры. Произрастали Р. во влажных, часто заболоченных местах, образуя заросли. Р. стали предками других низкоорганизованных ныне живущих групп высших растений — моховидных, плауновидных, хвощевидных, папоротниковидных.
Какова же роль стебля растения?Она так же важна, как роль листьев и корней.Стебель создает листьям условия, при которых они имеют возможность максимально использовать энергию солнца и воздух, откуда растения берут углекислый газ. Цветы, опыляемые ветром и насекомыми, плоды, распространяемые животными или ветром, тоже нуждаются в стебле, который, поднимая их над землей, способствует опылению или расселению.
А- миоплегия а - здоров
Р: Аа( отец болен) * аа( мать здорова)
Г: А а а
Дети : Аа(болен аа(здоров)
Ответ : 50 процентов
Спорофит-это взрослое растение ,на котором есть спарганий со спорами
спорангии-орган,в котором образуется коробочка,колосок,мешочек
гаметофит-взрослое растение или зелёная пластинка на которых образуются гаметы
фотосинтез-процесс образования в листьях на свету органических веществ(сахара,крахмала) из углекислого газа и воды
ВСЁ ОСТАЛЬНОЕ Я САМА ЕЩЁ НЕ ЗНАЮ.
Клеточная мембрана представляет собой оболочку, отделяющую содержимое клетки от внешней среды или соседних клеток. Основу клеточной мембраны составляет двойной слой липидов, в который погружены белковые молекулы, некоторые из них выполняют функцию рецепторов . Снаружи мембрана покрыта слоем гликопротеинов – гликокаликсом .
Кто является разрушителями органических веществ 1 папоротниковидным 2 хордовых животных 3 грибы и бактерии 4 членестоногие живот
Фактором . определяющим формирование природных зон является: 1)почва 2)рельеф 3)влажность воздуха 4)солнечная радиация
Скажите,пожалуйста,где расположен дыхательный центр? а)в среднем мозге б)в спинном мозге в)в продолговатом мозге г)в промежуточн

Риниофиты - псилофиты, самый древний и примитивный отдел вымерших растений.
Некоторые водоросли стали более совершенными и приспособились жить во влажных местах суши по берегам водоемов.
Начался переход некоторых растений от водного образа жизни к наземному.
Это произошло 350 - 400 млн.
Впервые остатки были найдены в 1859 канадским геологом в девонских отложениях на острове Гаспе.
Они характеризовались примитивностью строения, верхушечным расположением спорангиев, дихотомическим ветвлением спорофита, отсутствием корней и листьев и равноспоровостью.
Риниофиты стали предками других низко организованных ныне живущих групп высших растений - моховидных, плауновидных, хвощевидных, папоротниковидных.

Первыми наземными растениями были?
Первыми наземными растениями были?
Моховидные риниофиты плауновидные папоротниковидные.

Какую роль играют риниофиты?
Какую роль играют риниофиты?
Какова роль пыльцы у растений?
Какова роль пыльцы у растений.

Какова роль семян в жизни растений?
Какова роль семян в жизни растений?
Какова роль междоузлий в жизни растений?
Какова роль междоузлий в жизни растений.
Какова роль моховидных растений в природе?
Какова роль моховидных растений в природе.

Какова роль моховидных растений в природе?
Какова роль моховидных растений в природе?
Какова роль растений в природе?
Какова роль растений в природе?
Какова роль листа в жизни растения?
Какова роль листа в жизни растения?
Какова роль осенних растений для человека?
Какова роль осенних растений для человека?
Положительное : Водоросли насыщают толщу воды и воздуха над ней кислородом. Некоторые отмершие водоросли образуют осадочные породы : известняки, диатомит, трепел. Водоросли способствуют повышению плодородия почв. Из них добывают йод, бром, агар - ..
Номер 14. Биология, физиология, микология, энтомология, лепидоптерология . 1)Биология - это очень важный и интересный предмет в школе. 2)Мне очень нравится биология.

В микроскопе несколько объективов, револьерменяет их. Меняет зум рассматриваемого объекта и для того чтобы подимерсией смотреть.
Человек находится в покое и тем самым востоновляя свои силы(По - моему).

Да, у жесткокрылых есть половой димфорфизм.
Моего любимого дедушку зовут . . Он работал . . Родился . . Увлекается. . (Есть два сына). Я его люблю потому что он ( веселый) . Я его люблю и он меня тоже .

Моего дедушку зовут Виктор Давыдович. Его главные качества - доброта, мудрость, строгость и справедливость. Он очень любит меня, всегда мне помогает решать проблемы и радуется, когда я получаю отличные отметки. Он отлично учился в школе и универси..

Клетки семезачатков и яйцеклеток содержат диплоидный набор хромосом (48). В процессе мейоза, ДНК увеличивается надвое и получится 96 ДНК. Далее произойдет редукционное деление, вследствие чего получится 24 хромосомы и 48 ДНК. Ответ : 24 хромосомы ..

Жило одно Семечко у себя в цветке. Всё было сперва хорошо. Но потом вдруг подул ветер, домик сломался и Семечко вылетело наружу. Семечко испугалось, но потом решил построить новый дом. И решил его построить под землёй. Через несколько дней Семеч..
К отделу принадлежат самые древние из известных высших растений (рис. 2). Названия рода Риния (Rhynia) происходит от деревни Райни (недалеко от г. Абердина) в Шотландии, близ которой были обнаружены остатки этих примитивных растений в кремнистых породах врачом У. Макки в 1912 г. Эти отложения представляют собой пропитанные растворами кремниевой кислоты окаменевшие девонские торфа. У. Макки в свободное от медицинской практики время увлекался геологией. Свои материалы по ископаемым таксонам он передал шотландскому палеоботанику Р. Кидстону, который совместно с профессором Манчестерского университета У. Лангом опубликовали в 1917–1921 гг. результаты изучения растений ископаемой флоры. Они заложили начало систематическому изучению группы древнейших и наиболее примитивных высших растений.
ан
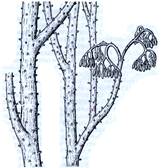
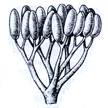



7б



Рис. 2. Риниеобразные (Rhyniophyta): 1 – риния большая (Rhynia major), (м – мезомы, р – ризоиды, рз – ризомоид, ст – стерильные теломы, фт – фертильные теломы); 2 – хорнеофитон (Horneophyton), (с – спорангии); 3а,3б – лионофитон (Lyonophyton), (3а – реконструкция гаметофита; 3б – обоеполый гаметангиофор: ан – антеридий, арх – архегоний); 4 – сциадофитон (Sciadophyton) с однополыми гаметангиофорами; 5 – спорангии хедеи (Hedeia); – синангий ярравии (Jarravia); 7а,7б – пертика (Pertica), (7а – реконструкция спорофита, 7б – сильно разветвленные спороносные оси); 8 – фрагмент псилофита первичного (Psilophyton princeps); 9 – зостеролист (Zosterophyllum); 10 – госслингия (Gosslingia); 11 – савдония (Sawdonia).
Древнейшие наземные растения раньше объединяли в один таксон под названием Псилофиты. Теперь разными исследователями предлагается 3 таксона на уровнях отделов. Многие объединяют их в отдел Риниеобразные – Rhyniophyta, насчитывающий около 20 родов и большое число видов. Единство их подтверждается рядом важных общих признаков, отражающих начальную ступень в эволюции высших растений. Единство их подкрепляемся также наличием родов, которые сочетают признаки трех отмеченных групп.
Отдел включает самые древние достоверные и по мнению большинства исследователей – самые примитивные травянистые высшие растения. Они появились приблизительно 430 млн. лет назад в силурийском периоде палеозойской эры. Геологическая история отдела заканчивается в девоне (около 370 млн. лет назад). Они дали начало другим группам высших растений.

Вегетативное тело спорофита риниофотов представлено безлистными цилиндрическими дихотомически ветвящимися, первично васкуляризованными осевыми структурами. Наземные оси выполняли в первую очередь функцию воздушного питания. Их конечные ответвления – теломы (от греч. telos – конец), а соединяющие их части, расположенные между точками ветвления – мезомы (от греч. mesos – средний). Одни теломы были стерильными, другие – фертильными и заканчивались спорангиями (рис 2). Стерильные теломы осуществляли только функцию фотосинтеза. Горизонтально расположенные на поверхности или в почве влажных местообитаний теломы получили название ризомоиды (т. е. корневищеподобные). От них отходили многочисленные простые (несептированные) одноклеточные ризоиды, осуществляя функцию минерального питания. Ризомоид – прототип корня, ризоиды – прототипы корневых волосков. Наличием проводящих пучков теломы принципиально отличались от структур водорослей.
Таким образом, теломы, мезомы и ризомоиды были первыми конструктивными элементами спорофита наземных растений. Возникновение этих элементарных органов было крупным ароморфозом в эволюции растительного мира. Это сильно повысило уровень организации растений, увеличило их пластичность и открыло более широкие возможности дальнейшего эволюционного развития.
Ветвление надземных вертикальных осей спорофита риниофитов характеризовалось равной и неравной дихотомией. Вследствие резко выраженной анизофилии, ветвление приобретало характер псевдомоноподиального. Поверхность надземных осей была голой или покрыта многочисленными выростами в виде энаций (в т.ч. шипиками, зубчиками).
Риниофиты – первые растения суши. Как наземные растения, в отличие от водорослей, они были покрыты типичной эпидермой, характеризовались наличием проводящей системы. Наружные стенки клеток эпидермы утолщены и покрыты слоем кутикулы. Имелись немногочисленные просто устроенные устьица (доказательство их наземного существования) – с двумя замыкающими клетками и устричной щелью. Они были лишены побочных клеток. Наличие устьиц в первую очередь отличает эпидерму высшего растения от наружного слоя клеток многоклеточных талломов (слоевищ) водорослей. Проводящая система слабо развита и построена по типу гаплостели, которая была началом развития всех остальных типов стели. Ксилема состояло из трахеид с кольчатыми, спиральными или реже лестничными утолщениями. Риниофты не обладали способностью к вторичному росту (камбий отсутствовал, как и перицикл). Меристема только верхушечная. В отличие от большинства сосудистых растений отсутствовали также и механические (опорные) ткани. Их функции выполняла толстая кора.
Спорангии всегда непосредственно прикреплялись к осям, или являлись их продолжением. У большинства более высоко организованных высших растений они прикрепляются к специализированным придаткам.
Спорангии риниофитов располагались верхушечно (терминально) на теломах или в боковом положении, одиночно (риния, куксония, хорнеофитон – Horneophyton, госслингия – Gosslingia) или группами (пертика – Pertica, тримерофит – Trimerophyton, зостеролист – Zosterophyllum). Они также отличались достаточно примитивным строением. Имели толстую многослойную стенку (спорангии эуспорангиатного типа), споры многочисленные, одинакового строения (растения морфологически равноспоровые). Спорангии различались по величине и форме. У наиболее примитивных видов они мало отличались от верхушек вегетативных осей. У куксонии они шаровидные, диаметром 1 мм. У ринии – продолговато-цилиндрические, длиной до 12 мм и толщиной до 4 мм. Спорангии не имели специальных приспособлений для вскрывания или они были крайне примитивно плана. У одних видов споры освобождались после разрушения стенки спорангия. У других видов спорангии вскрывались трещиной, располагавшейся продольно, поперечно (близ основания или верхушки спорангия), апикальной порой (у хорнеофитона). Нередко трещины проходили по определенным бороздам. В спорангий хорнеофитона глубоко вдавалась колонка, на которой располагался куполообразный спорангий. Интересно, но уже среди риниофитов наблюдалась тенденция к сближению спорангиев (хедея – Hedeia) и в дальнейшем к их срастанию в синангии (ярравия – Jarravia). Ярравия – нижнедевонское растение. В дальнейшем синангии будут неоднократно формироваться в различных линиях эволюции высших растений. Таким образом, спороносные структуры риниофитов, несмотря на примитивность их организации, характеризуются разнообразием морфологического строения. Эта особенность характерна для представителей большинства отделов высших растений и широко используется при классификации.
В спорангиях развивались многочисленные споры, одинакового морфологического строения (риниофиты были равноспоровыми растениями). Строение толстой оболочки спор – типичное для высших растений. Они были снабжены трехлучевым тетрадным рубцом.
Гаметофиты ринофитов существовали независимо от спорфитов и были свободно живущими растениями, однодомными (антеридии и архегонии развивались на одном растении). Под родовым названием лионофитон (Lyonophyton) описаны гаметофиты с обоеполыми гаметангиофорами. Предполагается, что они принадлежали хорнеофитону. На вершине некоторых неразветвленных осей имелось апикальное чашевидное расширение с лопастным краем. В центре чаши формировался небольшой выступ (архегониефор), к которому прикреплялись архегонии с сильно вытянутыми шейками. Булавовидные антеридии располагались по периферии чаши на внутренних стенках. Выявлены также гаметофиты с раздельнополыми гамитангиофорами (сциадофитон – Sciadophyton).
Произрастали риниофиты во влажных и болотистых местообитаниях вокруг морских и материковых водных бассейнов, а также в прибрежных мелководьях. Иногда они образовывали значительные заросли, являлись ландшафтными растениями. Возможно среди них были сухопутные растения. Некоторые были водными растениями с выступающими над водой спороносными частями.
Наиболее древней из достоверных находок высших растений является куксония. Более ранние риниофиты описаны только по спорам. Уже древнейшие силурийские риниофиты прошли длительный путь эволюционного развития. Это были довольно сложными, специализированными растениями разнообразного строения, приспособленными к жизни в наземных условиях.
Произошли они от зеленых водорослей. Наиболее древние формы еще сохраняли сходство с водорослеообразными предками. В частности, очень сходный характер их ветвления.
Многообразие признаков риниофитов явилось основой для дальнейших эволюционных направлений. Они дали начало многим независимым группам высших растений. В первую очередь плунообразным, хвощеобразным, псилотообразным и папоротникообразным, а также мохообразным (одна из гипотез). Изучение строения первенцев наземной суши имеет большое значение для эволюционной морфологии и филогении всего подцарства высших растений. Установлено, что первым органом спорофита высших растений был дихотомически разветвленный радиальный телом, строение которого, как отмечено, характерно для сухопутных растений (наличие проводящей системы, появление эпидермы, устьиц и т. д.). В ходе дальнейшего приспособления к наземной жизни от них произошли растения, обладавшие побегами и корнями.
В ходе эволюции растений из теломов формировались не только листья, но и другие органы высших растений. Например, срастание теломов, расположенных в разных плоскостях, привело к формированию стеблей с более сложным строением стели, чем протостела риниофитов. В процессе эволюции наземных растений наблюдались не только явления усложнения организации, но и явления редукции (уменьшение порядков ветвления, что приводило к упрощению строения органа). Таким путем сформировалась редуцированная листовая пластинка современных хвощей. Регрессивное развитие спорофита в целом наблюдалось у мохообразных. Одним из доказательств является наличие на спорофите некоторых печеночных и антоцеротовых мхов рудиментарных устьиц. При освоении растениями менее влажных местообитаний в процессе адаптации и последующей эволюции размеры листьев уменьшались из-за ослабления их роста. Появляются различные приспособления для ограничения транспирации. Это приводило к расширению адаптационных возможностей растений.
Некоторые листья стали выполнять функцию защиты зоны нарастания стебля и молодых зачатков листьев. Это обусловило появление почек. Сформировался побег (осевая часть – стебель, несущий листья и почки). Дифференциация и усложнение надземных органов, приведшая к формированию побегов, сопровождалась образованием корней в эволюции большинства высших растений для укрепления растения в почве и поглощения воды с растворенными минеральными веществами.
Спорофиллы и трофофиллы (вегетативные листья) согласно теломной теории возникают одновременно и независимо друг от друга. Дальнейшие их развитие шло параллельными путями. Следовательно, независимо друг от друга происходило также формирование вегетативных и спороносных побегов (стробилов). В процессе дальнейшей эволюции высших растений подземные органы совершенствовались, приобретая новые функции, видоизменялись, формируя метаморфозы (корневища, клубни, луковицы и др.). Возникают также новые ароморфозы (семена, цветки, плоды).
Как отмечено, со времени открытия, систематическое положение, классификация, таксономический ранг, объем единиц классификации риниофитов не однократно пересматривались. Это касается большинства групп ископаемых растений благодаря накоплению нового палеонтологического материала.
Некоторые исследователи разделяют риниофиты на 2 или 3 самостоятельных отдела, что не вполне оправдано. Все они связаны рядом важных общих признаков, отличающих их от более организованных высших споровых растений (плаунов, хвощей, папоротников). Они полностью лишены вторичных тканей, корней и побегов; оси всегда протостелические, стела нерасчлененная, спорангии прикрепляются непосредственно к наземным осям (терминально или латерально). Сходство прослеживается и по другим признакам (близость строения спор, ризоидов). Кроме того, существуют так называемые синтетические таксоны (тениокрада – Taeniocrada, куксония, стеганотека – Steganotheca, нотия – Nothia, реналия – Renalia и др.) в которых комбинируются признаки двух или даже трех указанных групп. Они, по-видимому, должны располагаться вместе, в составе одного отдела, в основе филогенетической системы высших растений, подчеркивая их монофилетическое происхождение.
Нередко отдел Риниеобразные условно разделяют на 2 класса – Риниеподобные и Зостеролистоподобные.
К отделу принадлежат самые древние из известных высших растений (рис. 2). Названия рода Риния (Rhynia) происходит от деревни Райни (недалеко от г. Абердина) в Шотландии, близ которой были обнаружены остатки этих примитивных растений в кремнистых породах врачом У. Макки в 1912 г. Эти отложения представляют собой пропитанные растворами кремниевой кислоты окаменевшие девонские торфа. У. Макки в свободное от медицинской практики время увлекался геологией. Свои материалы по ископаемым таксонам он передал шотландскому палеоботанику Р. Кидстону, который совместно с профессором Манчестерского университета У. Лангом опубликовали в 1917–1921 гг. результаты изучения растений ископаемой флоры. Они заложили начало систематическому изучению группы древнейших и наиболее примитивных высших растений.
Читайте также:
- Подарок воспитателю детского сада своими руками
- Рисование штрихом в детском саду в старшей группе шаблоны
- Влияние чикагской школы социальной экологии на механизмы городского развития
- Техносреда в доу развитие технические характеристики
- Особенности внимания у детей 6 7 лет которые необходимы к школьному обучению