
Характеристика межклеточных контактов кратко
Обновлено: 02.07.2024
Межклеточные контакты — соединения между клетками, образованные при помощи белков. Межклеточные контакты обеспечивают непосредственную связь между клетками. Кроме того, клетки взаимодействуют друг с другом на расстоянии с помощью сигналов (главным образом - сигнальных веществ), передаваемых через межклеточное вещество.
Содержание
Строение межклеточных соединений
В тех тканях, в которых клетки или их отростки плотно прилежат друг к другу (эпителий, мышечная ткань и пр.) между мембранами контактирующих клеток формируются связи – межклеточные контакты. Каждый тип межклеточных контактов формируется за счет специфических белков, подавляющее большинство которых — трансмембранные белки. Специальные адапторные белки могут соединять белки межклеточных контактов с цитоскелетом, а специальные "скелетные" белки - соединять отдельные молекулы этих белков в сложную надмолекулярную структуру. Во многих случаях межклеточные соединения разрушаются при удалении из среды ионов Ca 2+ .
Функции межклеточных соединений
Межклеточные соединения возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов (межклеточное взаимодействие), а также для механического скрепления клеток друг с другом.
Через щелевые контакты могут передаваться электрические сигналы. Клетки органов и тканей вырабатывают ряд химических веществ, действующих на другие клетки (в том числе через межклеточные контакты) и вызывающих изменения в работе цитоскелета, в интенсивности обмена веществ и процессе синтеза клеткой белков.
Типы межклеточных соединений
Плазмодесмы
Микроскопические цитоплазматические мостики, соединяющие соседние клетки растений. Основная статья: Плазмодесмы
Простое межклеточное соединение
Плотное соединение (запирающая зона)
В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания.
Зона замыкания
В зоне замыкания две соседние мембраны сливаются своими наружными слоями, эта зона непроницаема для макромолекул и ионов.
Зона слипания (промежуточный контакт)
В зоне слипания мембраны разделены щелью в 10-20 нм, заполненной плотным веществом (белковой природы).
Десмосома (пятно сцепления, липкое соединение)
Десмосома представляет собой небольшую площадку, иногда слоистого вида, диаметром до 0,5 мкм. Их функциональная роль заключается главным образом в механической связи между клетками. Существуют 3 типа десмосом – точечные, опоясывающие и полудесмосомы. Десмосомой называется образованное клетками соединение, прочно склеивающее клетки. Если они образуются между клетками и внеклеточным матриксом, то они называются полудесмосомами. Количество десмосом на одной клетке может достигать 2000. Такие контакты встречаются между клетками, которые могут подвергаться трению и другим механическим воздействиям (эпителиальные клетки, клетки сердечной мышцы). Со стороны цитоплазмы к десмосомам прикрепляются промежуточные филаменты, которые формируют остов цитоплазмы, обладающий большой прочностью на разрыв. Таким образом, через десмосомы промежуточные филаменты соседних клеток объединяются в непрерывную сеть по всей ткани. Тип промежуточных филаментов зависит от типа клеток: в большинстве эпителиальных клеток они кератиновые, а в клетках сердечной мышцы – десминовые.
Нексус (щелевой контакт)
Нексус представляет собой ограниченный участок контакта двух клеточных мембран диаметром 0,5 – 3 мкм с расстоянием между мембранами 2-3 нм. Обе эти мембраны пронизаны белковыми молекулами коннексонами, содержащими гидрофильные каналы. Через эти каналы осуществляется обмен ионами и микромолекулами соседних клеток. Поэтому нексусы называют также проводящими соединениями. Их функциональная роль заключается в переносе ионов и мелких молекул от клетки к клетке, минуя межклеточное пространство. Этот тип соединения встречается во всех группах тканей.
Синапс (синаптическое соединение)
Синапсы являются особыми формами межклеточных соединений. Они характерны для нервной ткани и встречаются между нейронами (межнейронные синапсы) или между нейроном и клеткой-мишенью (нервно-мышечные синапсы и пр.). Синапсы – участки контакта двух клеток, специализированных для односторонней передачи возбуждения или торможения от одной клетки к другой. Их функция – именно передача нервного импульса с нейрона на другую нервную клетку или клетку-мишень.
В тех тканях, в которых клетки или их отростки плотно прилежат друг к другу (эпителиальная, гладкомышечная и другие) между плазмолеммами контактирующих клеток формируются связи — межклеточные контакты.
Типы межклеточных контактов:
· щелевидный или нексус;
· синаптический контакт или синапс.
Простые контакты занимают наиболее обширные участки соприкасающихся клеток. Расстояние между билипидными мембранами соседних клеток составляет 15—20 нм, а связь между клетками осуществляется за счет взаимодействия макромолекул соприкасающихся гликокаликсов. Посредством простых контактов осуществляется слабая механическая связь — адгезия, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого контакта является контакт "типа замка", когда плазмолеммы соседних клеток вместе с участком цитоплазмы как бы впячивается в друг друга (интердигитация), чем достигается большая поверхность соприкосновения и более прочная механическая связь.
Десмосомные контакты или пятна сцепления представляют собой небольшие участки взаимодействия между клетками, диаметром около 0,5 мкм. Каждый такой участок (десмосома) имеет трехслойное строение и состоит из двух десмосомэлектронноплотных участков, расположенных в цитоплазме в местах контакта клеток, и скопления электронноплотного материала в межмембранном пространстве (15 20 нм). Количество десмосом на одной клетке может достигать 2 000. Функциональная роль десмосом обеспечение механической связи между клетками.
Плотные соединения или замыкательные пластинки обычно локализуются между эпителиальными клетками в тех органах (в желудке, кишечнике и других), в которых эпителий отграничивает агрессивное содержимое этих органов (желудочный сок, кишечный сок). Плотные контакты находятся только между апикальными частями эпителиальных клеток, охватывая по всему периметру каждую клетку. В этих участках межмембранные пространства отсутствуют, а билипидные слои соседних плазмолемм сливаются в одну общую билипидную мембрану. В прилежащих участках цитоплазмы соприкасающихся клеток отмечается скопление электронноплотного материала. Функциональная роль плотных контактов — прочная механическая связь клеток, препятствие транспорту веществ по межклеточным пространствам.
Щелевидные контакты или нексусы ограниченные участки контакта соседних цитолемм, диаметром 0,5—3,0 мкм, в которых билипидные мембраны сближены на расстояние 2—3 нм, а обе мембраны пронизаны в поперечном направлении белковыми молекулами коннексонами, содержащими гидрофильные каналы. Через эти каналы осуществляется обмен ионами и микромолекулами соседних клеток, чем и обеспечивается их функциональная связь (например, распространение биопотенциалов между кардиомиоцитами, их содружественное сокращение в миокарде).
Синаптические контакты или синапсы — специфические контакты между нервными клетками (межнейронные синапсы) или между нервными и другими клетками (нервно-мышечные синапсы и другие). Функциональная роль синаптических контактов заключается в передаче возбуждения или торможения с одной нервной клетки на другую или с нервной клетки на иннервируемую клетку.
В тех тканях, в которых клетки или их отростки плотно прилежат друг к другу (эпителиальная, гладкомышечная и другие) между плазмолеммами контактирующих клеток формируются связи — межклеточные контакты.
Типы межклеточных контактов:
· щелевидный или нексус;
· синаптический контакт или синапс.
Простые контакты занимают наиболее обширные участки соприкасающихся клеток. Расстояние между билипидными мембранами соседних клеток составляет 15—20 нм, а связь между клетками осуществляется за счет взаимодействия макромолекул соприкасающихся гликокаликсов. Посредством простых контактов осуществляется слабая механическая связь — адгезия, не препятствующая транспорту веществ в межклеточных пространствах. Разновидностью простого контакта является контакт "типа замка", когда плазмолеммы соседних клеток вместе с участком цитоплазмы как бы впячивается в друг друга (интердигитация), чем достигается большая поверхность соприкосновения и более прочная механическая связь.

Десмосомные контакты или пятна сцепления представляют собой небольшие участки взаимодействия между клетками, диаметром около 0,5 мкм. Каждый такой участок (десмосома) имеет трехслойное строение и состоит из двух десмосомэлектронноплотных участков, расположенных в цитоплазме в местах контакта клеток, и скопления электронноплотного материала в межмембранном пространстве (15 20 нм). Количество десмосом на одной клетке может достигать 2 000. Функциональная роль десмосом обеспечение механической связи между клетками.
Плотные соединения или замыкательные пластинки обычно локализуются между эпителиальными клетками в тех органах (в желудке, кишечнике и других), в которых эпителий отграничивает агрессивное содержимое этих органов (желудочный сок, кишечный сок). Плотные контакты находятся только между апикальными частями эпителиальных клеток, охватывая по всему периметру каждую клетку. В этих участках межмембранные пространства отсутствуют, а билипидные слои соседних плазмолемм сливаются в одну общую билипидную мембрану. В прилежащих участках цитоплазмы соприкасающихся клеток отмечается скопление электронноплотного материала. Функциональная роль плотных контактов — прочная механическая связь клеток, препятствие транспорту веществ по межклеточным пространствам.
Щелевидные контакты или нексусы ограниченные участки контакта соседних цитолемм, диаметром 0,5—3,0 мкм, в которых билипидные мембраны сближены на расстояние 2—3 нм, а обе мембраны пронизаны в поперечном направлении белковыми молекулами коннексонами, содержащими гидрофильные каналы. Через эти каналы осуществляется обмен ионами и микромолекулами соседних клеток, чем и обеспечивается их функциональная связь (например, распространение биопотенциалов между кардиомиоцитами, их содружественное сокращение в миокарде).
Синаптические контакты или синапсы — специфические контакты между нервными клетками (межнейронные синапсы) или между нервными и другими клетками (нервно-мышечные синапсы и другие). Функциональная роль синаптических контактов заключается в передаче возбуждения или торможения с одной нервной клетки на другую или с нервной клетки на иннервируемую клетку.
Экзоцитоз. Межклеточные контакты. Виды межклеточных контактов.
Важнейшее свойство клетки и ее плазмолеммы — формирование межклеточных соединений (контактов).
Простой неспециализированный (адгезионный) контакт образуется за счет элементов гликокаликса — трансмембранными гликопротеинами (кадгеринами) взаимодействующих мембран. Слои гликокаликса удерживают мембраны клеток на расстоянии около 10-20 нм, оставляя свободной межклеточную щель для транспортных процессов ионов и низкомолекулярных соединений. Обращенные в сторону межклеточной щели молекулы кадгеринов связываются катионами кальция. Простые контакты не обеспечивают высокой прочности межклеточных взаимодействий. Иногда плазмолеммы контактирующих клеток в области простого контакта образуют интердигитации (взаимные пальцевидные внедрения участков цитоплазмы), которые придают контакту большую прочность.
Плотный (запирающий) контакт характерен для клеток однослойных эпителиев. При формировании плотного контакта внешние слои мембран в отдельных участках максимально сближаются. В точках соприкосновения мембран располагаются интегральные белки плазмолемм соседних клеток. В ряде случаев (в эпителии кишечного типа) плотные контакты формируют сплошные полосы, получившие название замыкающих пластинок. Эти контакты, помимо прочного соединения клеток, изолируют межклеточные щели и делают их плохо проницаемыми для ионов и молекул.
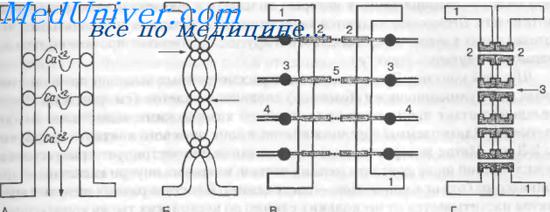
А — простой контакт (стрелки — движение веществ по межклеточному пространству); Б — плотный контакт (стрелки — зоны сближения внешнених слоев плазмолеммы); В — заякоривающий контакт (1 — цитоплазматические мембраны, 2 — связывающие трансмембранные белки - кадгерины, 3 — внутриклеточные белки сцепления — винкулин, 4 — фибриллярные белки цитоскелета, 5 — участки сцепления связывающих белков с помошью катионов кальция); Г — щелевой контакт (1 — цитоплазматические мембраны, 2 — коннексоны, 3 — каналы коннексонов)
Заякоривающий контакт. В отличие от двух предыдущих в его образовании кроме клеточных мембран участвуют фибриллярные элементы цитоскелета. К этому виду соединений принадлежат десмосомы и полудесмосомы, в формировании которых задействованы промежуточные филаменты цитоскелета.
Десмосома. В межклеточной щели в области десмосомы располагается электронно-плотный слой, образованный взаимодействующими молекулами интегральных гликопротеинов (десмоглеинов) плазмолемм соседних клеток. С помощью катионов кальция молекулы десмоглеина сцеплены в межклеточном пространстве. Со стороны гиалоплазмы в зоне десмосомы располагается электронно-плотный слой белка — десмоплакина, в который вплетаются промежуточные филаменты цитоскелета. Десмосомы являются характерными контактами эпителиальных, эндотелиальных клеток, кардиомиоцитов и других, обеспечивая прочное сцепление взаимодействующих структур.
Щелевой контакт. В отличие от всех рассмотренных выше он представляет собой коммуникационное (обменное) соединение клеток. Через щелевой контакт происходит прямой обмен химическими веществами между клетками. Плазмолеммы соседних клеток в зоне щелевого контакта сближены до 2-3 нм. Метод замораживания — скалывания демонстрирует присутствие в межклеточной щели контакта мелких частиц, имеющих внутри канальцы — это коннексоны (от англ. connection — соединение). В составе разных щелевых контактов насчитывается от нескольких единиц до нескольких тысяч коннексонов. Коннексоны насквозь пронизывают плазмолемму и в мембранах соседних клеток соединяются "стык в стык". В результате образуются сквозные каналы, сообщающие между собой внутренние среды контактирующих клеток. Коннексоны могут временно закрываться, ограничивая активность обменных процессов между клетками.
Таким образом, плазмолемма играет роль прочного механического и биологического покрова, изолируя внутреннее содержимое клетки от внешней среды. Эту функцию совместно с плазмолеммой выполняют гликокаликс и цитоплазматический кортекс актиновых филаментов. Кроме того, плазмолемма обеспечивает процессы переноса веществ из клетки во внеклеточную среду и наоборот — из внеклеточной среды внутрь клетки, а также является носителем информации о генетической принадлежности особи.
Информация на сайте подлежит консультации лечащим врачом и не заменяет очной консультации с ним.
См. подробнее в пользовательском соглашении.
Межклеточные контакты — соединения между клетками, образованные при помощи белков. Межклеточные контакты обеспечивают непосредственную связь между клетками. Кроме того, клетки взаимодействуют друг с другом на расстоянии с помощью сигналов (главным образом — сигнальных веществ), передаваемых через межклеточное вещество.

Функции межклеточных соединений
Межклеточные соединения возникают в местах соприкосновения клеток в тканях и служат для межклеточного транспорта веществ и передачи сигналов (межклеточное взаимодействие), а также для механического скрепления клеток друг с другом.
Через щелевые контакты могут передаваться электрические сигналы. Клетки органов и тканей вырабатывают ряд химических веществ, действующих на другие клетки (в том числе через межклеточные контакты) и вызывающих изменения в работе цитоскелета, в интенсивности обмена веществ и процессе синтеза клеткой белков.
Типы межклеточных соединений
Простое межклеточное соединение
Плотное соединение (запирающая зона)
В плотном соединении клеточные мембраны максимально сближены, здесь фактически происходит их слияние. Роль плотного соединения заключается в механическом сцеплении клеток и препятствии транспорту веществ по межклеточным пространствам. Эта область непроницаема для макромолекул и ионов, она ограждает межклеточные щели от внешней среды. Плотные соединения обычно образуются между эпителиальными клетками в тех органах (желудке, кишечнике и пр.), где эпителий ограничивает содержимое этих органов (желудочный сок, кишечный сок). В этих участках плотные контакты охватывают по периметру каждую клетку, межмембранные пространства отсутствуют, а соседние клеточные оболочки слиты в одну. Если же плотное сцепление происходит на ограниченном участке, то образуется пятно слипания (десмосома).Частными случаями плотного соединения являются зоны замыкания и слипания.

Плотные контакты состоят из тонких лент, пересекающихся между собой, которые полностью опоясывают клетку и контактируют с аналогичными лентами на соседних клетках. На электронных микрофотографиях заметно, что в участках плотных контактов мембраны соприкасаются одна с другой или даже сливаются. Комбинация метода замораживания-скалывания с электронной микроскопией с высоким разрешением позволила установить, что плёнки плотных контактов построены из белковых частиц диаметром 3-4 нм, которые выступают с обеих поверхностей мембраны. Также в пользу того, что в образовании плотных контактов ключевую роль играют белки, свидетельствует деление клеток под действием протеолитического фермента трипсина.
Всего в состав тесных контактов входит около 40 различных белков, как мембранных, так и цитоплазматических. Последние необходимы для прикрепления актиновых филаментов, регуляции и сигнализирования.
Мембранные белки плотных контактов можно разделить на две группы: те, которые пересекают мембрану 4 раза, и те, которые пересекают её только раз. Первая группа значительно распространена, в неё входят белки клаудины, окклюдины и трицеллюлин. Они имеют общие черты строения, в частности в них имеются четыре α-спиральных трасмембранных домена, N- и С-концы обращены к цитозолю, а домены, выступающие в межклеточное пространство, участвуют в гомо- или гетерофильных взаимодействиях с подобными белками на соседней клетке.
Основными белками плотных контактов являются клаудины (лат. claudo). Их роль была продемонстрирована на примере мышей с отсутствующим геном клаудин-1, — в эпидермисе таких животных не формируются плотные контакты и они погибают в течение дня после рождения из-за обезвоживания вследствие интенсивного испарения[1]. Клаудины также участвуют в формировании селективных каналов для транспорта ионов. В геноме человека есть гены по крайней мере 24 различных клаудинов, экспрессия которых происходит тканеспецифически.
Второе место по распространенности в плотных контактах занимают белки окклюдины (от лат. occludo — закрывать), они регулируют транспорт маленьких гидрофильных молекул и прохождение нейтрофилов через эпителий. Наибольшие концентрации третьего белка — трицеллюлина, наблюдаются в местах контакта трех клеток.
К белкам плотных контактов, пересекающим мембрану один раз, относятся JAM-A,-B,-C и-D (англ. junctional adhesion molecules) и родственные им CAR (англ. coxsackievirus and adenovirus receptor), CLMP (англ. CAR-like membrane protein) и ESAM (англ. endothelial-cell selective adhesion molecule), имеющие по два иммуноглобулинных домена, а также белки CRB3 (англ. Crumbs homologue 3) и Bves
Цитоплазматическая пластинка плотных контактов необходима для их присоединения к актиновым филаментам, регуляции сцепления клеток и параклеточного транспорта, а также для передачи сигналов от поверхности внутрь клетки. В её состав входят адаптерные, каркасные и цитоскелетные белки, а также элементы сигнальных путей (киназы, фосфатазы). Наиболее изучен белок цитоплазматической пластинки — ZO-1, он имеет несколько доменов белок-белкового взаимодействия, каждый из которых обеспечивает контакт с другими компонентами, в том числе три PDZ-домена (англ. PSD95–DlgA–ZO-1) — с клаудинами и другими адаптерными белками — ZO-2 и ZO-3, GUK-домен (англ. guanylate kinase homology) — с окклюдинами, а SH3-домен — с сигнальными белками.
С цитоплазматической стороной плотных контактов также ассоциированы комплексы белков PAR3/PAR6 и Pals1/PATJ, необходимые для установления полярности клеток и эпителиального морфогенеза.
Функции Первые исследования функций плотных контактов привели к представлению, что это статические непроницаемые структуры, необходимые для того, чтобы ограничить диффузию веществ между клетками. Впоследствии было выяснено, что они избирательно проницаемы, к тому же их пропускная способность отличается в различных тканях и может регулироваться. Также установлена ещё одна функция плотных контактов: роль в поддержании полярности клеток путем ограничения диффузии липидов и белков во внешнем слое плазматической мембраны. В первом десятилетии 21 века также накоплены данные, свидетельствующие об участии этих структур в сигнальных путях, в частности, регулирующих пролиферацию и полярность.
Регулирование парацеллюлярного транспорта
Непроницаемость плотных контактов в большинстве водорастворимых соединений может быть продемонстрирована в опыте по введению гидроксида лантана (электронно плотный коллоидный раствор) в кровеносные сосуды поджелудочной железы. Через несколько минут после инъекции ацинарные клетки фиксируются, и из них готовятся препараты для микроскопии. В таком случае можно наблюдать, что гидроксид лантана диффундирует из крови в пространство между латеральными поверхностями клеток, но не может проникнуть через плотные контакты в их верхней части. Другие опыты показали, что плотные контакты также непроницаемы для солей. Например при выращивании почек собаки MDCK (англ. Madin-Darby canine kidney) в среде с очень низкой концентрацией кальция, они формируют монослой, однако не сочетаются между собой плотными контактами. Через такой монослой могут свободно двигаться соли и жидкости. Если культуре добавить кальция, то за час формируются плотные контакты, и слой становится непроницаемым для жидкостей.
Однако не во всех тканях плотные контакты полностью непроницаемы, существуют так называемые неплотные эпителии (англ. leaky epithelia). Например, эпителий тонкого кишечника пропускает в 1000 раз больше ионов Na +, чем эпителий канальцев почек. Ионы проникают через параклеточные поры диаметром 4 Å, селективные по заряду и размеру частиц, которые формируются белками клаудинами. Поскольку эпителии различных органов эксрессируют различные наборы клаудинов, то отличается и их проницаемость для ионов. Например, специфический клаудин, присутствуюий только в почках, позволяет проходить ионам магния в процессе реабсорбции.
Межклеточное пространство эпителия может быть проницаемым и для больших частиц, например, при повторении упомянутого опыта с гидроксидом лантана на ткани эпителия тонкого кишечника кролика можно наблюдать прохождение коллоидных частиц между клетками. Крупные молекулы транспортируются через специальные пути утечки (англ. leak pathway) диаметром более 60 Å. Это важно, например, для процессов всасывания аминокислот и моносахаридов, концентрация которых в тонком кишечнике возрастает после еды достаточно для их пассивного транспорта.
Поддержание различия между апикальной и базолатеральной мембранами
Если в среду, контактирующую с апикальной частью монослоя MDCK-клеток, добавить липосомы, содержащие флуоресцентно меченые гликопротеины, некоторые из них спонтанно сливаются с клеточными мембранами. После этого флуоресценцию можно обнаружить в апикальной, но не в базолатеральной части клеток при условии целостности плотных контактов. Если же их разрушить, удалив из среды кальций, флуоресцентные белки диффундируют и равномерно распределяются по всей поверхности клетки.
Цитозольный слой мембраны имеет одинаковый липидный состав, как в апикальном, так и в базолатеральном участках, эти липиды могут свободно диффундировать. С другой стороны, липиды внеклеточного слоя двух частей клетки существенно различаются, и обмену между ними препятствуют плотные контакты. Например, все гликолипиды, как и белки заякоренные гликозилфосфатидилинозитолом, в мембранах MDCK клеток расположены исключительно в внеклеточном слое апикальной части, а фосфатидилхолин — почти исключительно в базолатеральной части.
Болезни, связанные с плотными контактами
С нарушением формирования тесных контактов связаны некоторые наследственные расстройства человека, например мутации в генах клаудина-16 и клаудина-19, которые приводят к гипомагниемии, вследствие чрезмерной потери магния с мочой. Мутации в гене клаудина-13 и трицеллюлина вызывают наследственную глухоту. Дисрегуляция некоторых белков плотных контактов связана с онкологическими заболеваниями, например экспрессия ZO-1 и ZO-2 снижается во многих типах рака. Компоненты тесных контактов также могут быть мишенями для онкогенных вирусов.
Некоторые вирусы используют мембранные белки плотных контактов для проникновения в клетку, в частности клаудин-1 является корецептором для вируса гепатита C. Другие вирусы присоединяются к белкам плотных контактов, чтобы разрушить барьер, отделяющий их от настоящих рецепторов на базолатеральной слое эпителиальных клеток, или неэпителиальных клетках.
Плотные контакты могут быть мишенью и для бактериальных патогенов, например Clostridium perfringens — возбудитель газовой гангрены, выделяет энтеротоксин (англ.), действующий на внеклеточные домены мембранных клаудинов и окклюдинов, и вызывает протечки эпителия. Helicobacter pylori — возбудитель гастрита — вводит в клетки белок CagA, взаимодействующий с комплексом ZO-1-JAM-A, считается, что это помогает бактерии преодолеть защитный барьер желудочного эпителия.
Адгезионные контакты

Адгезионные контакты (англ. adherens junctions, AJ) — якорные межклеточные контакты, ассоциированные с микрофиламентами, обеспечивающие целостность и механическую прочность ткани, в частности противостояние растяжению, придающие клеткам возможность координированно использовать актиновый цитоскелет. Адгезионные контакты относятся к гомофильным, то есть соединяют клетки одинакового типа. В их формировании принимают участие белки кадгерины и катенины.
Морфологически адгезионные контакты являются относительно простыми, в отличие от десмосом, плотных и щелевых контактов, они не имеют высокоспециализированных ультраструктур, кроме скопления актиновых филаментов. От других типов соединений клеток они отличаются относительной гибкостью и изменчивостью.
Чаще всего адгезионные контакты встречаются в эпителиальных тканях, здесь они образуют вокруг каждой клетки поясок, который называют зоной прилипания (англ. zonula adherens). Такие зоны в эпителии позвоночных животных преимущественно размещаются базальнее участка плотных контактов (англ. zonula occludens) и апикальнее десмосом (англ. macula adherens).
В зоне адгезионных контактов мембраны соседних клеток удалены друг от друга на расстояние 10-20 нм. В состав адгезионных контактов входят три основных элемента:
Схематическое изображение процесса формирования трубки из слоя эпителиальных клеток Одной из основных функций адгезионных контактов является физическое соединение клеток в единую ткань, их ослабление часто приводит к диссоциации клеток. Такого эффекта можно достичь, обработав ткань или монослойную культуру хелатирующим агентами, такими как ЭДТА, связывающими ионы кальция, вследствие чего взаимодействие между кадгеринами нарушается. Однако, хелатирующих агентов обычно недостаточно для полного разделения клеток, поскольку между ними существуют другие — кальций независимые — контакты.
Адгезионные контакты обеспечивают образование широкой межклеточной сети из пучков сократительных актиновых филаментов, расположенных параллельно мембранам клеток и соединенных между собой с помощью белков катенинов и кадгеринов. Такая организация позволяет не только противостоять механическому напряжению, но и согласовывать поведение клеток во время процессов морфогенеза. Например, координированное сокращение колец актиновых филаментов соседних клеток является необходимым для формирования трубок из слоя эпителия, в частности во время закладки нервной трубки. Одним из примеров является Shroom3-зависимое сокращение зоны прилипания, при этом актин-связывающий белок Shroom3 привлекает к участку адгезионные контакты Rho-киназы и активирует миозин-II, в результате чего и происходит сокращение.
Адгезионные контакты также задействованы в межклеточной передаче сигналов, об этом свидетельствует локализация в зрелых контактах рецепторной тирозинфосфатазы μ и белка RACK1, который взаимодействует с ней. При снижении экспрессии α-катенина наблюдается значительное увеличение темпов пролиферации эпителиальных клеток, было показано, что за этот эффект отвечает сигнальный путь инсулин / MAPK.
Соединения клеток, присутствующих в тканях и органах многоклеточных организмов, образуются сложными структурами, которые именуются межклеточными контактами . Особенно часто они обнаруживаются в эпителиях, пограничных покровных слоях.
Ученые полагают, что первичное отделение пласта элементов, связанных между собой межклеточными контактами , обеспечило формирование и последующее развитие органов и тканей.
Благодаря использованию методов электронной микроскопии удалось накопить большой объем сведений об ультраструктуре этих связей. Однако их биохимический состав, а также молекулярная структура изучены сегодня недостаточно точно.
Далее рассмотрим особенности, группы и виды межклеточных контактов .
Общие сведения
В образовании межклеточных контактов мембрана участвует очень активно. У многоклеточных за счет взаимодействия элементов формируются сложные клеточные образования. Их сохранение может обеспечиваться разными способами.
В эмбриональных, зародышевых тканях, в особенности на начальных этапах развития, клетки поддерживают связи друг с другом благодаря тому, что их поверхности обладают способностью слипаться. Такая адгезия (соединение) может быть связана со свойствами поверхности элементов.
Специфика возникновения
Исследователи полагают, что образование межклеточных контактов обеспечивается за счет взаимодействия гликокаликса с липопротеидами. При соединении всегда остается небольшая щель (ее ширина порядка 20 нм). В ней находится гликокаликс. При обработке ткани ферментом, способным нарушать его целостность или повреждать мембрану, клетки начинают обосабливаться друг от друга, диссоциируются.
Если удалить диссоциирующий фактор, клетки снова могут собраться вместе. Такое явление называют реагрегацией. Так можно разобщать клетки различных по своей окраске губок: желтых и оранжевых. В ходе экспериментов было установлено, что в соединении клеток возникает только 2 типа агрегатов. Одни состоят исключительно из оранжевых, а другие – только из желтых клеток. Смешанные суспензии, в свою очередь, самоорганизуются и восстанавливают первичную многоклеточную структуру.
Подобные результаты исследователи получили в ходе экспериментов с суспензиями разделенных эмбриональных клеток амфибий. В данном случае клетки эктодермы обосабливаются в пространстве избирательно от мезенхимы и энтодермы. Если для восстановления связей использовать ткани более поздних этапов развития зародышей, в пробирке будут самостоятельно собираться разные клеточные группы, отличающиеся органной и тканевой специфичностью, сформируются эпителиальные агрегаты, имеющие сходство с почечными канальцами.
Физиология: виды межклеточных контактов
Ученые выделяют 2 основные группы связей:
- Простые. Они могут образовывать соединения, различающиеся по форме.
- Сложные. В их число входят щелевидные, десмосомальные, плотные межклеточные контакты , а также адгезивные пояски и синапсы.
Рассмотрим их краткие характеристики.
Простые связи
Простые межклеточные контакты представляют собой участки взаимодействия надмембранных клеточных комплексов плазмолеммы. Расстояние между ними не более 15 нм. Межклеточные контакты обеспечивают прилипание элементов за счет взаимного "узнавания". Гликокаликс снабжен специальными рецепторными комплексами. Они строго индивидуальны для каждого отдельного организма.
Образование рецепторных комплексов является специфичным в пределах конкретной популяции клеток или определенных тканей. Они представлены интегринами и кадгеринами, имеющими сродство с аналогичными структурами клеток, находящихся по соседству. При взаимодействии с родственными молекулами, расположенными на прилежащих цитомембранах, происходит их слипание – адгезия.
Межклеточные контакты в гистологии
Среди адгезивных протеинов выделяют:
- Интегрины.
- Иммуноглобулины.
- Селектины.
- Кадгерины.
Некоторые белки, обладающие адгезивными свойствами, не относятся ни к одному из указанных семейств.
Характеристики семейств
Некоторые гликопротеины поверхностного клеточного аппарата относятся к основному комплексу гистосовместимости 1-го класса. Как и интегрины, они строго индивидуальны для отдельного организма и специфичны для тканевых образований, в которых располагаются. Некоторые вещества обнаруживаются только в определенных тканях. К примеру, Е-кадгерины являются специфичными для эпителия.
Интегринами называют интегральные белки, которые состоят из 2-х субъединиц – альфа- и бета-. В настоящее время выявлено 10 вариантов первых и 15 видов вторых. Внутриклеточные участки связываются с тонкими микрофиламентами с помощью специальных белковых молекул (танина или винкулина) либо напрямую с актином.
Селектины представляют собой мономерные белки. Они узнают определенные углеводные комплексы и прикрепляются к ним на поверхности клеток. В настоящее время наиболее изученными являются L, Р и Е-селектины.
Иммуноглобулиноподобные адгезивные белки по своему строению похожи на классические антитела. Некоторые из них являются рецепторами при иммунологических реакциях, другие предназначены только для реализации адгезивных функций.
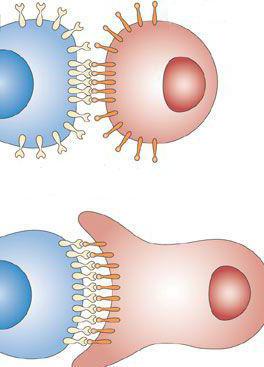
Межклеточные контакты кадгеринов возникают только при наличии кальциевых ионов. Они задействованы в образовании постоянных связей: Р и Е-кадгерины в эпителиальных тканях, а N-кадгерины – в мышечной и нервной.
Назначение
Следует сказать, что межклеточные контакты предназначены не только для простого сцепления элементов. Они необходимы для обеспечения нормального функционирования тканевых структур и клеток, в формировании которых задействованы. Простые контакты контролируют созревание и перемещение клеток, предупреждают гиперплазию (излишнее увеличение количества структурных элементов).
Разнообразие соединений
В ходе исследований установлены разные типы межклеточных контактов по форме. Они могут быть, к примеру, в виде "черепицы". Такие связи формируются в роговом слое плоского многослойного ороговевающего эпителия, в артериальном эндотелии. Известны также зубчатый и пальцевидный типы. В первом выпячивание одного элемента погружается в вогнутую часть другого. За счет этого существенно увеличивается механическая прочность соединения.
Сложные связи
Эти виды межклеточных контактов специализированы для реализации какой-то определенной функции. Такие соединения представлены небольшими парными специализированными участками плазматических мембран 2-х соседних клеток.
Существуют следующие виды межклеточных контактов:
- Запирающие.
- Сцепляющие.
- Коммуникационные.
Десмосомы
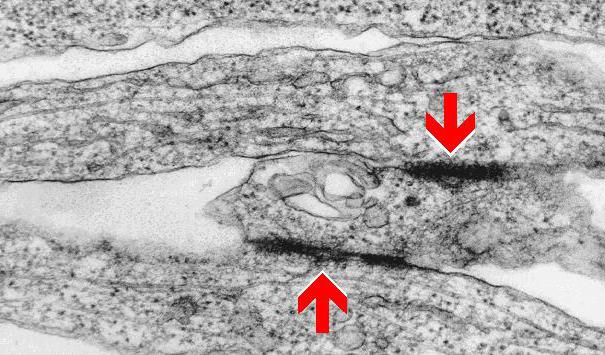
Они являются сложными макромолекулярными образованиями, посредством которых обеспечивается прочное соединение соседних элементов. При электронной микроскопии такой вид контакта заметен очень хорошо, поскольку его отличает высокая электронная плотность. Локальный участок выглядит в форме диска. Его диаметр порядка 0,5 мкм. Мембраны соседних элементов в нем расположены на расстоянии от 30 до 40 нм.
Рассмотреть участки высокой электронной плотности можно и на внутренних мембранных поверхностях обеих взаимодействующих клеток. К ним прикреплены промежуточные филаменты. В ткани эпителия эти элементы представлены тонофиламентами, которые образуют скопления – тонофибриллы. В тонофиламентах присутствуют цитокератины. Между мембранами также обнаруживается электронно-плотная зона, которая соответствует сцеплению протеиновых комплексов соседних клеточных элементов.
Как правило, десмосомы встречаются в эпителиальной ткани, но выявить их можно и в других структурах. В таком случае в промежуточных филаментах содержатся вещества, свойственные этой ткани. К примеру, в соединительных структурах присутствуют виментины, в мышцах – десмины и пр.
Внутренняя часть десмосомы на макромолекулярном уровне представлена десмоплакинами – опорными белками. С ними соединяются промежуточные филаменты. Десмоплакины, в свою очередь, скреплены с десмоглеинами с помощью плакоглобинов. Это тройное соединение проходит сквозь липидный слой. Десмоглеины связываются с белками, находящимися в соседней клетке.
Возможен, однако, и другой вариант. Прикрепление десмоплакинов осуществляется к интегральным белкам, находящимся в мембране, - десмоколинам. Они, в свою очередь, связываются с подобными протеинами соседней цитомембраны.
Поясная десмосома
Она также представлена в виде механического соединения. Однако отличительной ее чертой является форма. Выглядит поясная десмосома в виде ленты. Подобно ободку поясок сцепления охватывает цитолемму и соседние клеточные мембраны.
Этот контакт отличается высокой электронной плотностью и в области мембран, и на участке расположения межклеточного вещества.
В пояске сцепления присутствует винкулин – опорный белок, выступающий в качестве участка прикрепления микрофиламентов к внутренней части цитомембраны.
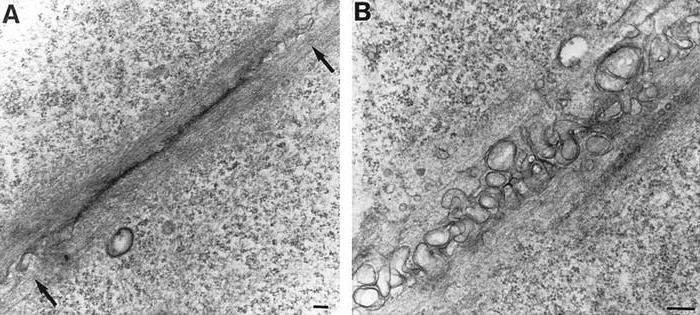
Адгезивную ленту можно обнаружить в апикальном участке однослойного эпителия. Она часто примыкает к плотному контакту. Отличительной чертой этого соединения является то, что в его структуру входят актиновые микрофиламенты. Они располагаются параллельно относительно поверхности мембраны. За счет их способности сокращаться при наличии минимиозинов и нестабильности целый пласт эпителиальных клеток, а также микрорельеф поверхности органа, которую они выстилают, могут изменять свою форму.
Щелевидный контакт
Его называют также нексусом. Как правило, так соединяются эндотелиоциты. Межклеточные контакты щелевидного типа имеют форму диска. Его протяженность составляет 0,5-3 мкм.
На участке соединения соседние мембраны находятся на расстоянии 2-4 нм друг от друга. В поверхности обеих контактирующих элементов присутствуют интегральные протеины – коннектины. Они, в свою очередь, интегрируются в коннексоны – белковые комплексы, состоящие из 6 молекул.
Коннексоновые комплексы прилегают друг к другу. В центральной части каждого расположена пора. Через нее свободно могут проходить элементы, молекулярная масса которых не превышает 2 тыс. Поры в соседних клетках плотно присоединяются друг у другу. За счет этого происходит перемещение молекул неорганических ионов, воды, мономеров, низкомолекулярных биологически активных веществ только в соседнюю клетку, а в межклеточное вещество они не проникают.
Функции нексусов
За счет щелевидных контактов передается возбуждение соседним элементам. К примеру, так проходят импульсы между нейронами, гладкими миоцитами, кардиомиоцитами и пр. За счет нексусов обеспечивается единство биореакций клеток в тканях. В нервных тканевых структурах щелевидные контакты именуются электрическими синапсами.
Задачи нексусов состоят в формировании межклеточного внутритканевого контроля над биоактивностью клеток. Кроме того, такие контакты выполняют несколько специфических функций. К примеру, без них не было бы единства сокращения сердечных кардиомиоцитов, синхронных реакций клеток гладких мышц и пр.
Плотный контакт
Его называют также запирающей зоной. Он представлен в виде участка слияния поверхностных мембранных слоев соседних клеток. Эти зоны формируют непрерывную сеть, которая "сшита" интегральными белковыми молекулами мембран соседних клеточных элементов. Эти белки формируют структуру, похожую на ячеистую сеть. Ею окружен периметр клетки в виде пояска. При этом структура соединяет соседние поверхности.
Часто к плотному контакту прилегают ленточные десмосомы. Этот участок непроницаем для ионов и молекул. Следовательно, он запирает межклеточные щели и, собственно, внутреннюю среду всего организма от внешних факторов.
Значение запирающих зон
Плотный контакт препятствует диффузии соединений. К примеру, содержимое желудочной полости защищено от внутренней среды его стенок, белковые комплексы не могут перемещаться от свободной эпителиальной поверхности в межклеточное пространство и пр. Запирающая зона способствует также поляризации клетки.
Плотные контакты являются основой разнообразных барьеров, присутствующих в организме. При наличии запирающих зон перенос веществ в соседние среды осуществляется исключительно через клетку.
Синапсы
Они представляют собой специализированные соединения, расположенные в нейронах (нервных структурах). За счет них обеспечивается передача информации от одних клеток к другим.
Синаптическое соединение обнаруживается в специализированных участках и между двумя нервными клетками, и между нейроном и другим элементом, включенным в состав эффектора либо рецептора. К примеру, выделяют нервно-эпителиальные, нервно-мышечные синапсы.
Эти контакты разделяют на электрические и химические. Первые аналогичны щелевидным связям.
Сцепление с межклеточным веществом
Клетки присоединяются за счет рецепторов цитолеммы к адгезивным белкам. К примеру, рецепторы к фибронектину и ламинину в клетках эпителия обеспечивают сцепление с этими гликопротеинами. Ламинин и фибронектин являются адгезивными субстратами с фибриллярным элементом базальных мембран (IV тип коллагеновых волокон).
Полудесмосома
Со стороны клетки ее биохимический состав и строение подобен дисмосоме. От клетки в межклеточное вещество отходят особые якорные филаменты. За счет них объединяется мембрана с фибриллярным каркасом и заякоривающие фибриллы коллагеновых волокон VII типа.
Точечный контакт
Его также называют фокальным. Точечный контакт входит в группу сцепляющих соединений. Наиболее характерным он считается для фибробластов. Клетка в таком случае сцепляется не с соседним клеточным элементами, а с межклеточными структурами. Рецепторные протеины взаимодействуют с адгезивными молекулами. К ним относят хондронектин, фибронектин и пр. Они связывают клеточные мембраны с внеклеточными волокнами.
Формирование точечного контакта осуществляется за счет актиновых микрофиламентов. Они закрепляются на внутренней части цитолеммы при помощи интегральных белков.
Читайте также: