
Фотохимический этап фотосинтеза кратко
Обновлено: 05.07.2024
Задачи: Сформировать знания о реакциях пластического и энергетического обменов и их взаимосвязи; вспомнить особенности строения хлоропластов. Дать характеристику световой и темновой фазы фотосинтеза. Показать значение фотосинтеза как процесса, обеспечивающего синтез органических веществ, поглощение углекислого газа и выделение кислорода в атмосферу.
Тип урока: лекция.
Оборудование:
- Средства наглядности: таблицы по общей биологии;
- ТСО: компьютер; мультимедиапроектор.
План лекции:
- История изучения процесса.
- Эксперименты по фотосинтезу.
- Фотосинтез, как анаболический процесс.
- Хлорофилл и его свойства.
- Фотосистемы.
- Световая фаза фотосинтеза.
- Темновая фаза фотосинтеза.
- Лимитирующие факторы фотосинтеза.
Ход лекции
История изучения фотосинтеза
1630 год начало изучения фотосинтеза. Ван Гельмонт доказал, что растения образуют органические вещества, а не получают их из почвы. Взвешивая горшок с землей и ивой, и отдельно само дерево, он показал, что через 5 лет масса дерева увеличилась на 74 кг, тогда как почва потеряла только 57 г. Он решил, что пищу дерево получает из воды. В настоящее время мы знаем, что используется углекислый газ.
В 1804 году Соссюр установил, что в процессе фотосинтеза велико значение воды.
В 1887 году открыты хемосинтезирующие бактерии.
В 1905 году Блэкман установил, что фотосинтез состоит из двух фаз: быстрой – световой и ряда последовательных медленных реакций темновой фазы.
Эксперименты по фотосинтезу
3 опыт доказывает значение фотосинтеза (рис.3.)
Фотосинтез, как анаболический процесс
- Ежегодно в результате фотосинтеза образуется 150 млрд. тонн органического вещества и 200 млрд. тонн свободного кислорода.
- Круговорот кислорода, углерода и других элементов, вовлекаемых в фотосинтез. Поддерживает современный состав атмосферы, необходимый для существования современных форм жизни.
- Фотосинтез препятствует увеличению концентрации углекислого газа, предотвращая перегрев Земли вследствие парникового эффекта.
- Фотосинтез – основа всех цепей питания на Земле.
- Запасенная в продуктах энергия – основной источник энергии для человечества.
Сущность фотосинтеза заключается в превращении световой энергии солнечного луча в химическую энергию в виде АТФ и НАДФ·Н2.
Суммарное уравнение фотосинтеза:
Существует два главных типа фотосинтеза:
| анаэробный | аэробный |
| Характерен для фотосинтезирующих бактерий (подцарство Настоящие бактерии). Фотосинтезирующим пигментом у них является бактериохлорофилл. Кислород не выделяется. | Характерен для всех оксифотобактерий и зеленых растений. Фотосинтез в растениях осуществляется в хлоропластах содержащих хлорофилл. Кислород выделяется. |
Хлорофилл и его свойства
Виды хлорофилла
Хлорофилл имеет модификации а, в, с, d. Отличаются они структурным строением и спектром поглощения света. Например: хлорофилл в содержит на один атом кислорода больше и на два атома водорода меньше, чем хлорофилл а.
Все растения и оксифотобактерии имеют как основной пигмент желто-зеленый хлорофилл а, а как дополнительный хлорофилл в.
Другие пигменты растений
Некоторые другие пигменты способны поглощать солнечную энергию и передавать ее в хлорофилл, вовлекая ее тем самым в фотосинтез.
У большинства растений есть темно оранжевый пигмент – каротин, который в животном организме превращается в витамин А и желтый пигмент – ксантофилл.
Фикоцианин и фикоэритрин – содержат красные и сине-зеленые водоросли. У красных водорослей эти пигменты принимают более активное участие в процессе фотосинтеза, чем хлорофилл.
Хлорофилл минимально поглощает свет в сине-зеленой части спектра. Хлорофилл а, в- в фиолетовой области спектра, где длина волны 440 нм. Уникальная функция хлорофилла состоит в том, что он интенсивно поглощает солнечную энергию и передает ее другим молекулам.
Пигменты поглощают определенную длину волны, не поглощенные участки солнечного спектра отражаются, что обеспечивает окраску пигмента. Зеленый свет не поглощается, поэтому хлорофилл зеленый.
Пигменты – это химические соединения, которые поглощают видимый свет, что приводит электроны в возбужденное состояние. Чем меньше длина волны, тем больше энергия света и больше его способность переводить электроны в возбужденное состояние. Это состояние неустойчиво и вскоре вся молекула возвращается в свое обычное низкоэнергетическое состояние теряя при этом энергию возбуждения. Эта энергия может быть использована на флуоресценцию.
Фотосистемы
Каждая система состоит из набора вспомогательных пигментов (от 250 до 400 молекул), передающих энергию на одну молекулу главного пигмента и она называется реакционным центром. В нем энергия Солнца используется для фотохимических реакций.
 Рис.5. Фотосистемы | Фотосистема I имеет более мелкие частицы, чем фотосистема II. Частицы фотосистемы II связаны с гранами. Энергия захватывается как бы в ловушку со вспомогательных (антенных) пигментов на главный. Это может быть хлорофилл а – Р690 или Р700 (Р – пигмент, а 690-700 – максимально поглощенная длина волны в нм). Р690 и Р700 – энергетические ловушки |
| Фотосинтез | ||
| Световая фаза Фотофизический этап | Световая фаза Фотохимический этап | Темновая фаза или цикл Кальвина |
| Поглощение квантов света пигментами, идет возбуждение электронов в этих молекулах и передача возбуждения от одной молекулы к другой. | Преобразование энергии света в энергию химических связей АТФ и НАДФ.Н2. Идет в фотосинтетических мембранах. | Идет за счет энергии, которая образовалась в световой фазе. Суть процесса: включение углекислого газа в образование органических веществ. |
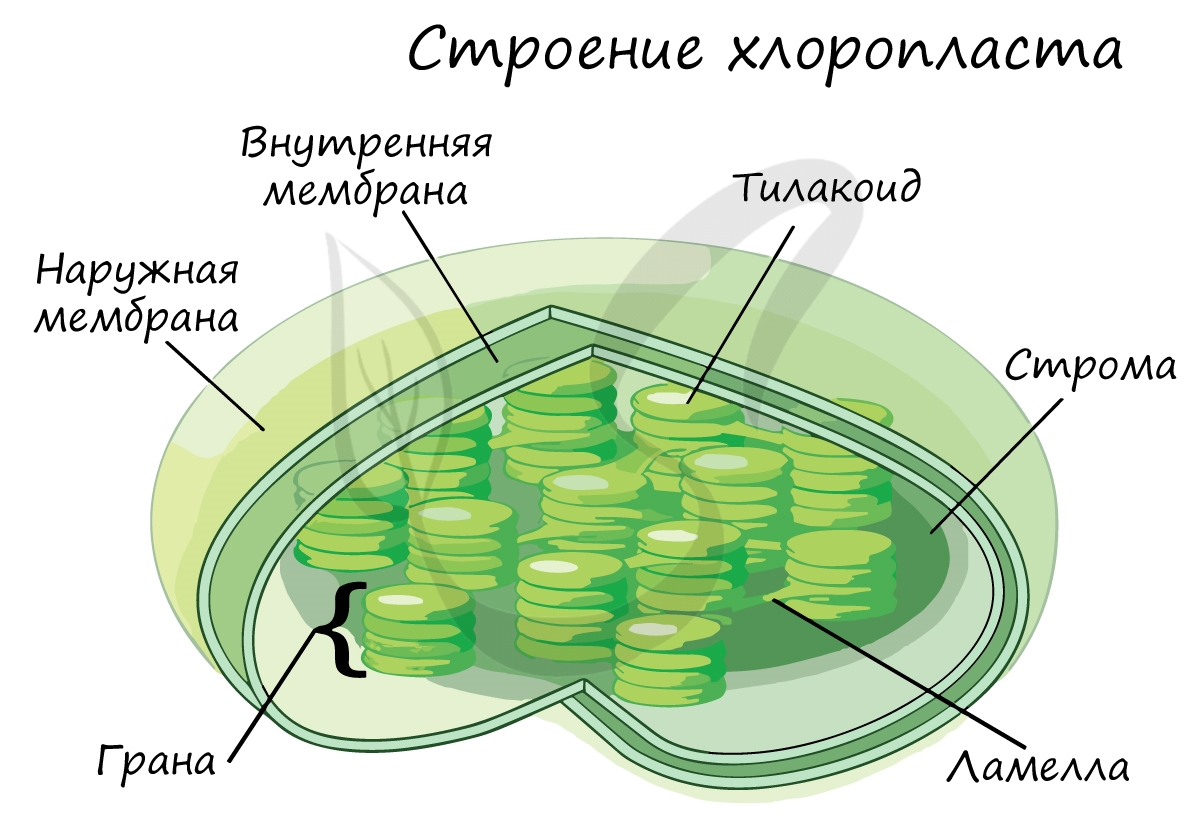
Световая фаза идет обязательно с участием света, темновая фаза и на свету и в темноте. Световой процесс происходит в тилакоидах хлоропластов, темновой – в строме, т.е. эти процессы пространственно разобщены.
Световая фаза фотосинтеза
В 1958 году Арнон и его сотрудники изучили световую фазу фотосинтеза. Они установили, что источником энергии при фотосинтезе является свет, а так как на свету в хлорофилле происходит синтез из АДФ+Ф.к. → АТФ, то этот процесс называется фосфорилированием. Оно сопряжено с переносом электронов в мембранах.
Роль световых реакций: 1. Синтез АТФ – фосфорилирование. 2. Синтез НАДФ.Н2.
Путь переноса электронов называется Z-схемой.
Z-схема. Нециклическое и циклическое фотофосфорилирование (рис. 6.)

Начало процесса. Поглощение квантов света. Квант света попадает на ФС II, находящуюся в мембранах тилакоидов гран и приводит к возбуждению пигментов – это возбуждение передается от одной молекулы антенного пигмента к другой вплоть до реакционного центра. Все электроны собираются вокруг ловушки и отдают энергию виде электронов в электроннотранспортную цепь. Электрон, поглотив фотон, отрывается от молекулы хлорофилла и переходит на более высокий энергетический уровень присоединяясь к молекулам-переносчикам. Затем он двигается по электроннотранспортной цепи переходя от одного переносчика к другому (от пластохинона к пластоцианину) постепенно растрачивая энергию. Часть этой энергии растрачивается на синтез АТФ.
Нециклическое фотофосфорилирование. Растратив энергию электрон достигает ФСI, где он опять поглощает фотон и снова поднимается еще на более высокий энергетический уровень, и пройдя через несколько переносчиков (ферредоксин) передается конечному акцептору цепи НАДФ + , который расположен на внешней стороне мембраны тилакоида.
Процесс химио-осмоса.

Рис.7. Процесс фотосинтеза
- Свет, попадая на молекулы хлорофилла, которые находятся в мембранах тилакоидов гран, приводит их в возбужденное состояние. В результате этого электроны сходят со своих орбит и переносятся с помощью переносчиков за пределы мембраны тилакоида, где накапливаются, создавая отрицательно заряженное электрическое поле.
- Протоны, образовавшиеся при фотолизе, не проникают через мембрану тилакоида и накапливаются внутри, образуя положительно заряженное электрическое поле, что приводит к увеличению разности потенциалов по обе стороны мембраны.
- При достижении критической разности потенциалов, протоны могут выходить в строму по протонному каналу. С каналами связаны ферменты АТФ-синтетазы, которые используют энергию протонов на синтез АТФ. На каждые три протона, которые проходят через канал, синтезируется одна молекула АТФ. Большая часть АТФ при фотосинтезе образуется этим путем.
- Протоны, вышедшие на поверхность мембраны тилакоида, соединяются с электронами, образуя атомарный водород, который идет на восстановление переносчика НАДФ + .
Циклический поток электронов. ФС I может работать независимо от ФСII. Под действием света, электрон выбивается из молекулы хлорофилла реакционного центра ФС I, передается к тому же акцептору, что и при нециклическом потоке, но далее идет не к НАДФ + , а по обходному пути возвращается на тоже место в ФС I. Поглощенная электроном энергия используется на синтез АТФ. Таким образом электрон двигается по кольцу. Это циклический поток.
В ходе циклического транспорта электронов не происходит образования НАДФ.Н2 и фоторазложения Н2О, следовательно и выделение О2. Этот путь используется тогда, когда в клетке избыток НАДФ.Н2, но требуется дополнительная АТФ.
Все эти процессы относятся к световой фазе фотосинтеза. В дальнейшем энергия АТФ и НАДФ.Н2 используется для синтеза глюкозы. Для этого процесса свет не нужен. Это реакции темновой фазы фотосинтеза.
Темновая фаза фотосинтеза или цикл Кальвина
Синтез глюкозы происходит в ходе циклического процесса, который получил название по имени ученого Мельвина Кальвина, открывшего его, и награжденного Нобелевской премией.

Рис. 8. Цикл Кальвина
Каждая реакция цикла Кальвина осуществляется своим ферментом. Для образования глюкозы используются: СО2, протоны и электроны от НАДФ.Н2, энергия АТФ и НАДФ.Н2. Происходит процесс в строме хлоропласта. Исходным и конечным соединением цикла Кальвина, к которому с помощью фермента рибулозодифосфаткарбоксилазы присоединяется СО2, является пятиуглеродный сахар – рибулозобифосфат, содержащий две фосфатные группы. В результате образуется шестиуглеродное соединение, сразу же распадающееся на две трехуглеродные молекулы фосфоглицериновой кислоты, которые затем восстанавливаются до фосфоглицеринового альдегида. При этом, часть образовавшегося фосфоглицеринового альдегида используется для регенерации рибулозобифосфата, и, таким образом, цикл возобновляется снова (5С3 → 3С5), а часть используется для синтеза глюкозы и других органических соединений (2С3 → С6 → С6Н12О6).
Для образования одной молекулы глюкозы необходимо 6 оборотов цикла и требуется 12НАДФ.Н2 и 18 АТФ. Из суммарного уравнения реакции получается:
Из приведенного уравнения видно, что атомы С и О вошли в глюкозу из СО2, а атомы водорода из Н2О. Глюкоза в дальнейшем может быть использована как на синтез сложных углеводов (целлюлозы, крахмала), так и на образование белков и липидов.
(С4 – фотосинтез. В 1965 году было доказано, что у сахарного тростника – первыми продуктами фотосинтеза, являются кислоты, содержащие четыре атома углерода (яблочная, щавелевоуксусная, аспарагиновая). К С4 растениям принадлежат кукуруза, сорго, просо).
Лимитирующие факторы фотосинтеза
Скорость фотосинтеза – наиболее важный фактор влияющий на урожайность с/х культур. Так, для темновых фаз фотосинтеза нужны НАДФ.Н2 и АТФ, и поэтому скорость темновых реакций зависит от световых реакций. При слабой освещенности скорость образования органических веществ будет мала. Поэтому свет – лимитирующий фактор.
Из всех факторов одновременно влияющих на процесс фотосинтеза лимитирующим будет тот, который ближе к минимальному уровню. Это установил Блэкман в 1905 году. Разные факторы могут быть лимитными, но один из них главный.
-
При низкой освещенности скорость фотосинтеза прямопропорциональна интенсивности света. Свет – лимитирующий фактор при низкой освещенности. При большой интенсивности света происходит обесцвечивание хлорофилла и фотосинтез замедляется. В таких условиях в природе растения обычно защищены (толстая кутикула, опушенные листья, чешуйки).
Фотохимические реакции фотосинтеза — это реакции, в которых энергия света преобразуется в энергию химических связей, и в первую очередь в энергию фосфорных связей АТФ. Именно АТФ является энергетической валютой клетки, обеспечивающей течение всех процессов. Одновременно под действием света происходит разложение воды, образуется восстановленный НАДФ и выделяется кислород. Энергия поглощенных квантов света стекается от сотен молекул пигментов ССК к фотохимическому реакционному центру, содержащему особую пару (димер) молекул хлорофилла а, которые характеризуются поглощением в наиболее длинноволновой части солнечного спектра и выполняют роль ловушки энергии возбуждения, блуждающего по пигментам ССК. Наряду с димером хлорофилла в фотосинтетический комплекс входят молекулы первичного и вторичного акцепторов электрона. Молекула хлорофилла, отдавая электрон первичному акцептору, окисляется. Электрон поступает в электронтранспортную цепь.
Совокупность светособирающего комплекса (ССК), фотохимического реакционного центра и связанных с ним молекул — переносчиков электрон составляет фотосистему. По современным представлениям фотосистема является важнейшим структурно-функциональным звеном фотосинтетического аппарата.
Различают два типа потока электронов — циклический и нециклический. При циклическом потоке электроны, переданные от молекулы хлорофилла первичному акцептору, возвращаются к ней обратно. При нециклическом потоке происходит фотоокисление воды и передача электрона от воды к НАДФ. Выделяемая в ходе окислительно-восстановительных реакций энергия частично используется на синтез АТФ. Процесс преобразования энергии света в энергию АТФ получил название фотосинтетического фосфорилирования (Арнон). Соответственно двум типам потока электронов различают циклическое и нециклическое фотофосфорилирование. При нециклическом потоке электронов принимают участие две фотосистемы. Как уже упоминалось, энергия квантов света стекается к молекуле пигмента П700, который входит в состав реакционного центра ФС I. Электрон в молекуле П700 переходит на более высокий энергетический уровень. Тенденция к потере электрона (окислению) выражена очень слабо. Однако при поглощении кванта света молекула П700переходит в возбужденное состояние и легко отдает электрон первичному акцептору (фотохимическая реакция). Энергия света изменяет распределение электронов в кольце молекулы хлорофилла П700 и делает один электрон доступным для переноса. При этом энергия квантов сначала переходит в энергию электронного возбуждения, а затем в энергию разделения зарядов.

После последовательной передачи четырех электронов от ВРК к П680+ происходит разложение
сразу двух молекул воды, сопровождающееся выделением одной молекулы кислорода и четырех
ионов водорода, которые попадают во внутритилакоидное пространство хлоропласта. Перенос электрона по цепи переносчиков от фотосистемы II к фотосистеме I сопровождается образованием АТФ из АДФ и неорганического Фн (АДФ + Фн —> АТФ). Суммарное уравнение процесса нециклического фотофосфорилирования может быть выражено следующим образом:
2НАДФ + 2Н20 + 2АДФ + 2Н3Р04 -> 2НАДФН + 2Н+ + 2АТФ + 02.
Таким образом, отличительными особенностями нециклического фотосинтетического фосфорилирования являются:1) окисление двух молекул воды;2) участие двух фотосистем;3) передача электронов от молекул воды (первичный донор) через электронтранспортную цепь на НАДФ (конечный акцептор). Продуктами процесса нециклического фотофосфорилирования являются восстановленный никотинамидадениндинуклеотидфосфат (НАДФН + Н+) и АТФ. Эти соединения в дальнейшем используются в темновой фазе фотосинтеза.
В циклическом процессе принимает участие лишь фотосистема I. В результате поглощения кванта света молекула П700 отдает возбужденный электрон сначала первичному переносчику, а затем от ферредоксина возвращается к П700. Суммарное уравнение циклического фотосинтетического фосфорилирования выглядит следующим образом:
АДФ + Н3Р04 + hv —> АТФ + Н20.
Существуют две гипотезы относительно механизма фосфорилирования (прямой механизм и косвенный). Согласно первой гипотезе фосфатная группа и АДФ связываются с ферментом в активном участке комплекса Fr Два протона перемещаются через канал по градиенту концентрации и соединяются с кислородом фосфата, образуя воду. Это делает фосфатную группу весьма реакционноспособной, и она присоединяется к АДФ с образованием АТФ. Согласно второй гипотезе, которая в последние годы получила экспериментальные подтверждения (косвенный механизм), АДФ и Фн соединяются в активном центре фермента спонтанно. Однако образовавшаяся АТФ прочно связана с ферментом, и для ее освобождения требуется энергия. Энергия доставляется протонами, которые, связываясь с ферментом (протонирование), изменяют его конформацию, после чего АТФ высвобождается. Продукты световой фазы, в которых аккумулирована энергия света, — АТФ и НАДФН Д. Арнон назвал ассимиляционной силой, поскольку они используются в темновую фазу, в ходе которой С02 восстанавливается до углеводов.
Синтез АТФ
Синтез АТФ осуществляется тремя способами: фотосинтетическое фосфорилирование, окислительное фосфорилирование (сопряжённое с транспортом электронов по дыхательной цепи) и субстратное фосфорилирование. В первых двух процессах преобразование поступившей с потоком электронов энергии в энергию фосфоэфирных связей АТФ осуществляет особый фермент — АТФ-синтетаза. Этот фермент присутствует во всех мембранах, участвующих в преобразовании энергии (мембраны бактерий, митохондрий и хлоропластов). АТФ-синтетаза катализирует присоединение неорганического фосфата (Фн) к АДФ, образование которого осуществляет аде-нилаткиназа
(АМФ + АТФ = 2 АДФ).
Активность АТФ-синтетазы можно обнаружить по обратной реакции гидролиза АТФ:
АТФ + Н20 = АДФ + Фн + Н+.
Благодаря обратимости реакции фосфорилирования, накопившийся АТФ может быть использован для создания протонного градиента, обеспечивающего энергией движение жгутиков и осмотическую работу. Энергия также направляется для обратного переноса электронов, необходимого для восстановления никотинамидадениндинуклеотида (НАД) при использовании бактериями неорганических доноров электронов (S03, N03, Fe2+ и др.).
Дыхательная электронтранспортная цепь (ЭТЦ) — система структурно и функционально связанных трансмембранных белков и переносчиков электронов. ЭТЦ позволяет запасти энергию, выделяющуюся в ходе окисления НАД∙Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану.
Дыхательная электронтранспортная цепь состоит из переносчиков электронов, которые передают электроны от субстратов на кислород. Расположение переносчиков определяется величиной их окислительно-восстановительного потенциала.
Таким образом, транспорт электронов в дыхательной электронтранспортной цепи
сопровождается трансмембранным переносом протонов. Возникающая разность потенциалов
по обеим сторонам внутренней мембране митохондрий используется для синтеза АТФ (окислительное фосфорилирование). В результате прохождения двух электронов по цепи образуется 3 молекулы АТФ.
Компоненты ЭТЦ: НАД(Ф) – зависимые дегидрогеназы, растворимые ферменты; Белки – содержащие железоероцентры, изменение валентности; Хиноны – жирорастворимые соединения, имеющие длинный терпеноидный хвост, связанный с хиноидным ядром, наиболее распространен убихинон; цитохромы – переносят только электроны за счет изменения валентности железа из +2 в +3; колплекс цитохромов – цитохромоксидаза, присутствие Си вместо железа.
Дыхательная цепь митохондрий содержит 4 основных комплекса и два небольших по молекулярной массе компонента – убихинон и цитохром. Комплекс 1 осуществляет перенос электронов от НАДН к убихинону. Комплекс 2 катализирует окисление сукцината убихиноном. Комплекс 3 переносит электроны от восстановленного убихинона к цитохрому. В комплексе 4 электроны переносятся от цитохрома к кислороды.
Фотохимические реакции фотосинтеза — это реакции, в которых энергия света преобразуется в энергию химических связей, и в первую очередь в энергию фосфорных связей АТФ. Именно АТФ является энергетической валютой клетки, обеспечивающей течение всех процессов. Одновременно под действием света происходит разложение воды, образуется восстановленный НАДФ и выделяется кислород. Энергия поглощенных квантов света стекается от сотен молекул пигментов ССК к фотохимическому реакционному центру, содержащему особую пару (димер) молекул хлорофилла а, которые характеризуются поглощением в наиболее длинноволновой части солнечного спектра и выполняют роль ловушки энергии возбуждения, блуждающего по пигментам ССК. Наряду с димером хлорофилла в фотосинтетический комплекс входят молекулы первичного и вторичного акцепторов электрона. Молекула хлорофилла, отдавая электрон первичному акцептору, окисляется. Электрон поступает в электронтранспортную цепь.
Совокупность светособирающего комплекса (ССК), фотохимического реакционного центра и связанных с ним молекул — переносчиков электрон составляет фотосистему. По современным представлениям фотосистема является важнейшим структурно-функциональным звеном фотосинтетического аппарата.
Различают два типа потока электронов — циклический и нециклический. При циклическом потоке электроны, переданные от молекулы хлорофилла первичному акцептору, возвращаются к ней обратно. При нециклическом потоке происходит фотоокисление воды и передача электрона от воды к НАДФ. Выделяемая в ходе окислительно-восстановительных реакций энергия частично используется на синтез АТФ. Процесс преобразования энергии света в энергию АТФ получил название фотосинтетического фосфорилирования (Арнон). Соответственно двум типам потока электронов различают циклическое и нециклическое фотофосфорилирование. При нециклическом потоке электронов принимают участие две фотосистемы. Как уже упоминалось, энергия квантов света стекается к молекуле пигмента П700, который входит в состав реакционного центра ФС I. Электрон в молекуле П700 переходит на более высокий энергетический уровень. Тенденция к потере электрона (окислению) выражена очень слабо. Однако при поглощении кванта света молекула П700переходит в возбужденное состояние и легко отдает электрон первичному акцептору (фотохимическая реакция). Энергия света изменяет распределение электронов в кольце молекулы хлорофилла П700 и делает один электрон доступным для переноса. При этом энергия квантов сначала переходит в энергию электронного возбуждения, а затем в энергию разделения зарядов.
После последовательной передачи четырех электронов от ВРК к П680+ происходит разложение
сразу двух молекул воды, сопровождающееся выделением одной молекулы кислорода и четырех
ионов водорода, которые попадают во внутритилакоидное пространство хлоропласта. Перенос электрона по цепи переносчиков от фотосистемы II к фотосистеме I сопровождается образованием АТФ из АДФ и неорганического Фн (АДФ + Фн —> АТФ). Суммарное уравнение процесса нециклического фотофосфорилирования может быть выражено следующим образом:
2НАДФ + 2Н20 + 2АДФ + 2Н3Р04 -> 2НАДФН + 2Н+ + 2АТФ + 02.
Таким образом, отличительными особенностями нециклического фотосинтетического фосфорилирования являются:1) окисление двух молекул воды;2) участие двух фотосистем;3) передача электронов от молекул воды (первичный донор) через электронтранспортную цепь на НАДФ (конечный акцептор). Продуктами процесса нециклического фотофосфорилирования являются восстановленный никотинамидадениндинуклеотидфосфат (НАДФН + Н+) и АТФ. Эти соединения в дальнейшем используются в темновой фазе фотосинтеза.
В циклическом процессе принимает участие лишь фотосистема I. В результате поглощения кванта света молекула П700 отдает возбужденный электрон сначала первичному переносчику, а затем от ферредоксина возвращается к П700. Суммарное уравнение циклического фотосинтетического фосфорилирования выглядит следующим образом:
АДФ + Н3Р04 + hv —> АТФ + Н20.
Существуют две гипотезы относительно механизма фосфорилирования (прямой механизм и косвенный). Согласно первой гипотезе фосфатная группа и АДФ связываются с ферментом в активном участке комплекса Fr Два протона перемещаются через канал по градиенту концентрации и соединяются с кислородом фосфата, образуя воду. Это делает фосфатную группу весьма реакционноспособной, и она присоединяется к АДФ с образованием АТФ. Согласно второй гипотезе, которая в последние годы получила экспериментальные подтверждения (косвенный механизм), АДФ и Фн соединяются в активном центре фермента спонтанно. Однако образовавшаяся АТФ прочно связана с ферментом, и для ее освобождения требуется энергия. Энергия доставляется протонами, которые, связываясь с ферментом (протонирование), изменяют его конформацию, после чего АТФ высвобождается. Продукты световой фазы, в которых аккумулирована энергия света, — АТФ и НАДФН Д. Арнон назвал ассимиляционной силой, поскольку они используются в темновую фазу, в ходе которой С02 восстанавливается до углеводов.
Синтез АТФ
Синтез АТФ осуществляется тремя способами: фотосинтетическое фосфорилирование, окислительное фосфорилирование (сопряжённое с транспортом электронов по дыхательной цепи) и субстратное фосфорилирование. В первых двух процессах преобразование поступившей с потоком электронов энергии в энергию фосфоэфирных связей АТФ осуществляет особый фермент — АТФ-синтетаза. Этот фермент присутствует во всех мембранах, участвующих в преобразовании энергии (мембраны бактерий, митохондрий и хлоропластов). АТФ-синтетаза катализирует присоединение неорганического фосфата (Фн) к АДФ, образование которого осуществляет аде-нилаткиназа
(АМФ + АТФ = 2 АДФ).
Активность АТФ-синтетазы можно обнаружить по обратной реакции гидролиза АТФ:
АТФ + Н20 = АДФ + Фн + Н+.
Благодаря обратимости реакции фосфорилирования, накопившийся АТФ может быть использован для создания протонного градиента, обеспечивающего энергией движение жгутиков и осмотическую работу. Энергия также направляется для обратного переноса электронов, необходимого для восстановления никотинамидадениндинуклеотида (НАД) при использовании бактериями неорганических доноров электронов (S03, N03, Fe2+ и др.).
Дыхательная электронтранспортная цепь (ЭТЦ) — система структурно и функционально связанных трансмембранных белков и переносчиков электронов. ЭТЦ позволяет запасти энергию, выделяющуюся в ходе окисления НАД∙Н и ФАДН2 молекулярным кислородом (в случае аэробного дыхания) или иными веществами (в случае анаэробного) в форме трансмембранного протонного потенциала за счёт последовательного переноса электрона по цепи, сопряжённого с перекачкой протонов через мембрану.
Дыхательная электронтранспортная цепь состоит из переносчиков электронов, которые передают электроны от субстратов на кислород. Расположение переносчиков определяется величиной их окислительно-восстановительного потенциала.
Таким образом, транспорт электронов в дыхательной электронтранспортной цепи
сопровождается трансмембранным переносом протонов. Возникающая разность потенциалов
по обеим сторонам внутренней мембране митохондрий используется для синтеза АТФ (окислительное фосфорилирование). В результате прохождения двух электронов по цепи образуется 3 молекулы АТФ.
Компоненты ЭТЦ: НАД(Ф) – зависимые дегидрогеназы, растворимые ферменты; Белки – содержащие железоероцентры, изменение валентности; Хиноны – жирорастворимые соединения, имеющие длинный терпеноидный хвост, связанный с хиноидным ядром, наиболее распространен убихинон; цитохромы – переносят только электроны за счет изменения валентности железа из +2 в +3; колплекс цитохромов – цитохромоксидаза, присутствие Си вместо железа.
Дыхательная цепь митохондрий содержит 4 основных комплекса и два небольших по молекулярной массе компонента – убихинон и цитохром. Комплекс 1 осуществляет перенос электронов от НАДН к убихинону. Комплекс 2 катализирует окисление сукцината убихиноном. Комплекс 3 переносит электроны от восстановленного убихинона к цитохрому. В комплексе 4 электроны переносятся от цитохрома к кислороды.
По типу питания живые организмы делятся на автотрофы, гетеротрофы и миксотрофы. Автотрофы (греч. αὐτός — сам + τροφ - пища) - организмы, которые самостоятельно способны синтезировать органические вещества из неорганических. Гетеротрофы (греч. ἕτερος - иной + τροφή - пища) - организмы, использующие для питания готовые органические вещества.
Наконец, миксотрофы (греч. μῖξις - смешение + τροφή - пища) - организмы, которые могут использовать как гетеротрофный, так и автотрофный способ питания. К примеру, эвглена зеленая на свету начинает фотосинтезировать, а в темноте питается гетеротрофно.

Фотосинтез
Фотосинтез (греч. φῶς - свет и σύνθεσις - синтез) - сложный химический процесс преобразования энергии квантов света в энергию химических связей. В результате фотосинтеза происходит синтез органических веществ из неорганических.

Этот процесс уникален и происходит только в растительных клетках, а также у некоторых бактерий. Фотосинтез осуществляется при участии хлорофилла (греч. χλωρός - зелёный и φύλλον - лист) - зеленого пигмента, окрашивающего органы растений в зеленый цвет. Существуют и другие вспомогательные пигменты, которые вместе с хлорофиллом выполняют светособирающую или светозащитную функции.
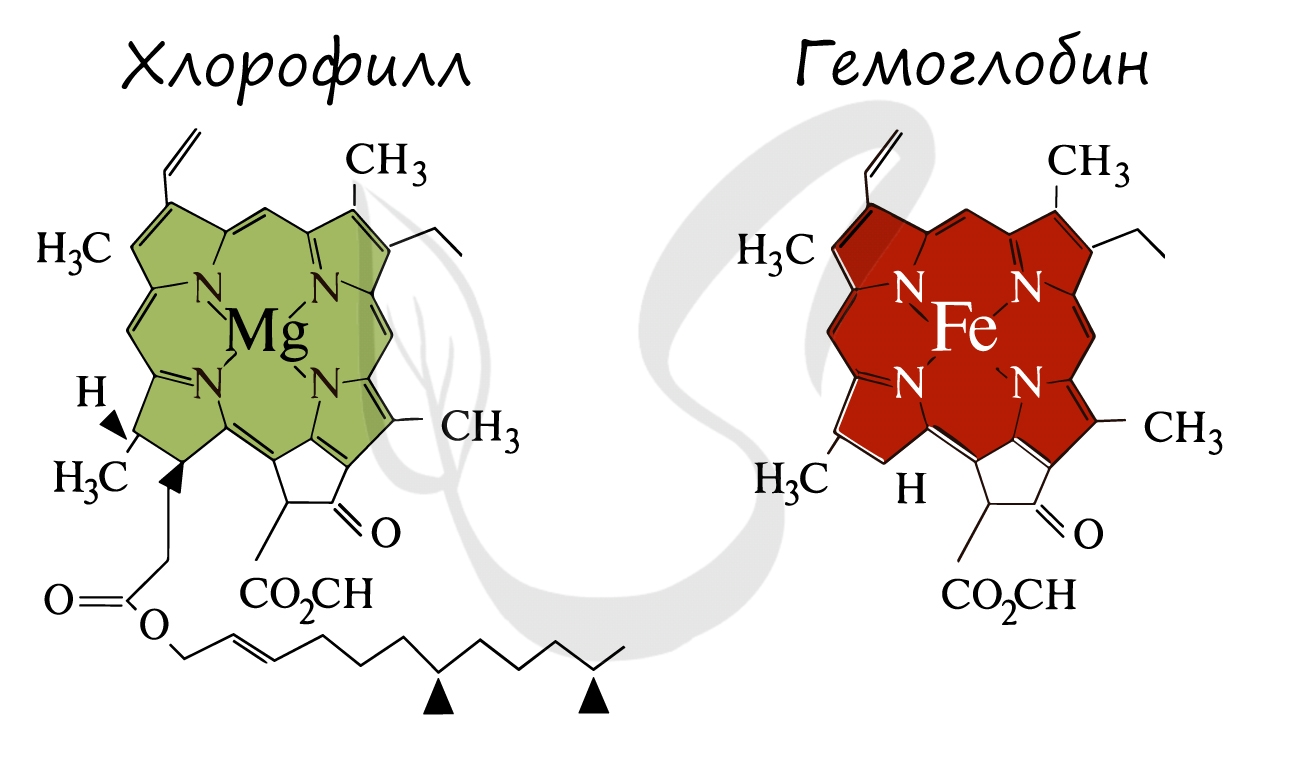
Ниже вы увидите сравнение строения хлорофилла и гемоглобина. Обратите внимание, что в центре молекулы хлорофилла находится ион Mg.
В высшей степени гениально значение процесса фотосинтеза подчеркнул русский ученый К.А. Тимирязев: "Все органические вещества, как бы они ни были разнообразны, где бы они ни встречались, в растении ли, в животном или человеке, прошли через лист, произошли от веществ, выработанных листом. Вне листа или, вернее, вне хлорофиллового зерна в природе не существует лаборатории, где бы выделялось органическое вещество. Во всех других органах и организмах оно превращается, преобразуется, только здесь оно образуется вновь из вещества неорганического"
Более подробно мы обсудим значение фотосинтеза в завершение этой статьи. Фотосинтез состоит из двух фаз: светозависимой (световой) и светонезависимой (темновой). Я рекомендую использовать названия светозависимая и светонезависимая, так как они способствуют более глубокому (и правильному!) пониманию фотосинтеза.
Светозависимая фаза (световая)
Эта фаза происходит только на свету на мембранах тилакоидов в хлоропластах. В ней принимают участие различные ферменты, белки-переносчики, молекулы АТФ-синтетазы и зеленый пигмент хлорофилл.
Хлорофилл выполняет две функции: поглощения и передачи энергии. При воздействии кванта света хлорофилл теряет электрон, переходя в возбужденное состояние. С помощью переносчиков электроны скапливаются с наружной поверхности мембраны тилакоидов, тем временем внутри тилакоида происходит фотолиз воды (разложение под действием света):
Гидроксид-ионы отдают лишний электрон, превращаясь в реакционно способные радикалы OH, которые собираются вместе и образуют молекулу воды и свободный кислород (это побочный продукт, который в дальнейшем удаляется в ходе газообмена).
Образовавшиеся при фотолизе воды протоны (H + ) скапливаются с внутренней стороны мембраны тилакоидов, а электроны - с внешней. В результате по обе стороны мембраны накапливаются противоположные заряды.
При достижении критической разницы, часть протонов проталкивается на внешнюю сторону мембраны через канал АТФ-синтетазы. В результате этого выделяется энергия, которая может быть использована для фосфорилирования молекул АДФ:
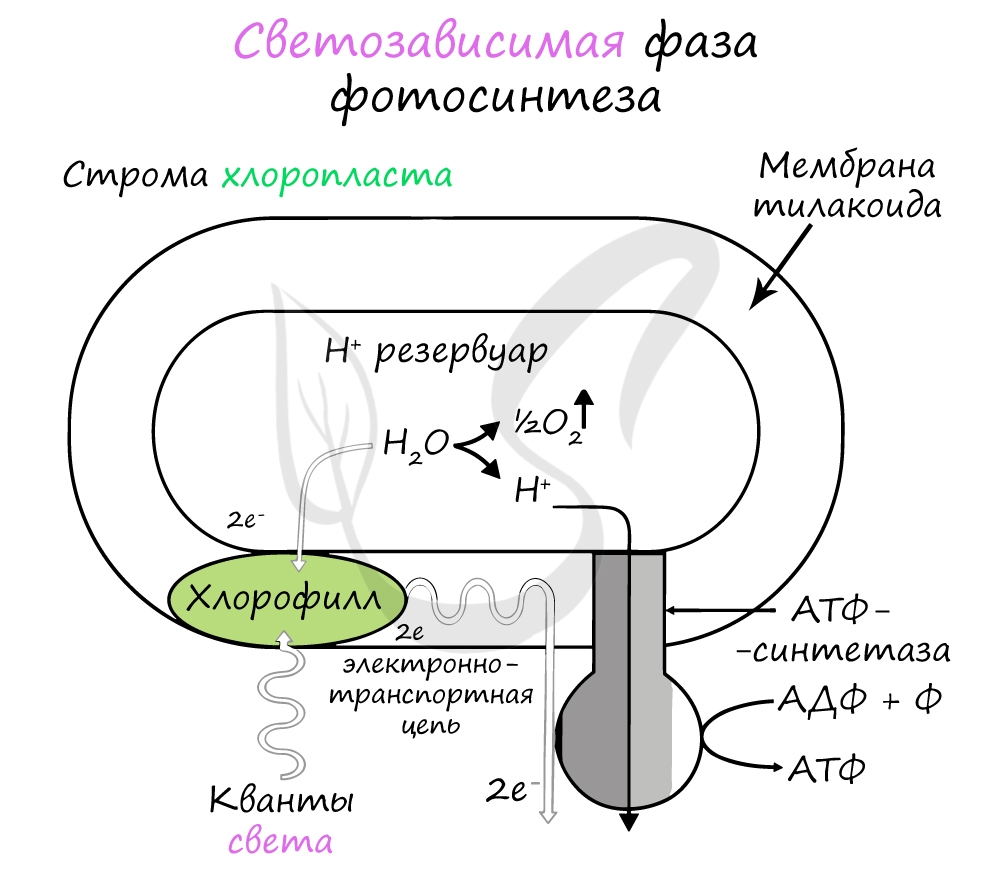
Протоны, попав на поверхность мембраны тилакоидов, соединяются с электронами и образуют атомарный водород, который используется для восстановления молекулы-переносчика НАДФ (никотинамиддинуклеотидфосфат). Благодаря этому окисленная форма - НАФД + превращается в восстановленную - НАДФ∗H2.
- Свободный кислород O2 - в результате фотолиза воды
- АТФ - универсальный источник энергии
- НАДФ∗H2 - форма запасания атомов водорода
Кислород удаляется из клетки как побочный продукт фотосинтеза, он совершенно не нужен растению. АТФ и НАДФ∗H2 в дальнейшем оказываются более полезны: они транспортируются в строму хлоропласта и принимают участие в светонезависимой фазе фотосинтеза.
Светонезависимая (темновая) фаза
Светонезависимая фаза происходит в строме (матриксе) хлоропласта постоянно: и днем, и ночью - вне зависимости от освещения.
При участии АТФ и НАДФ∗H2 происходит восстановление CO2 до глюкозы C6H12O6. В светонезависимой фазе происходит цикл Кальвина, в ходе которого и образуется глюкоза. Для образования одной молекулы глюкозы требуется 6 молекул CO2, 12 НАДФ∗H2 и 18 АТФ.
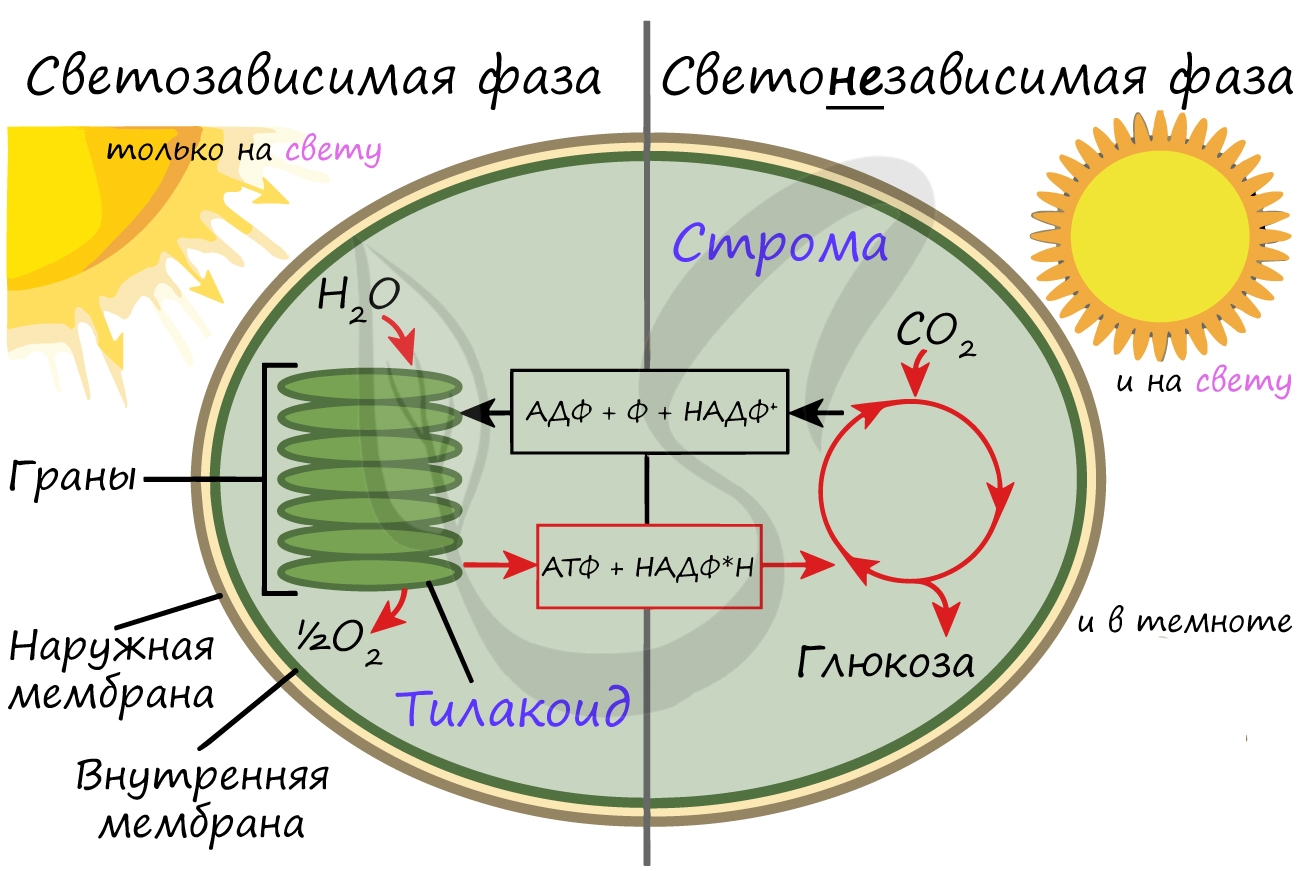
Таким образом, в результате темновой (светонезависимой) фазы фотосинтеза образуется глюкоза, которая в дальнейшем может быть преобразована в крахмал, служащий для запасания питательных веществ у растений.
Значение фотосинтеза
Значение фотосинтеза невозможно переоценить. Уверенно утверждаю: именно благодаря этому процессу жизнь на Земле приобрела такие чудесные и изумительные формы, какие мы видим вокруг себя: удивительные растения, прекрасные цветы и самые разнообразные животные.
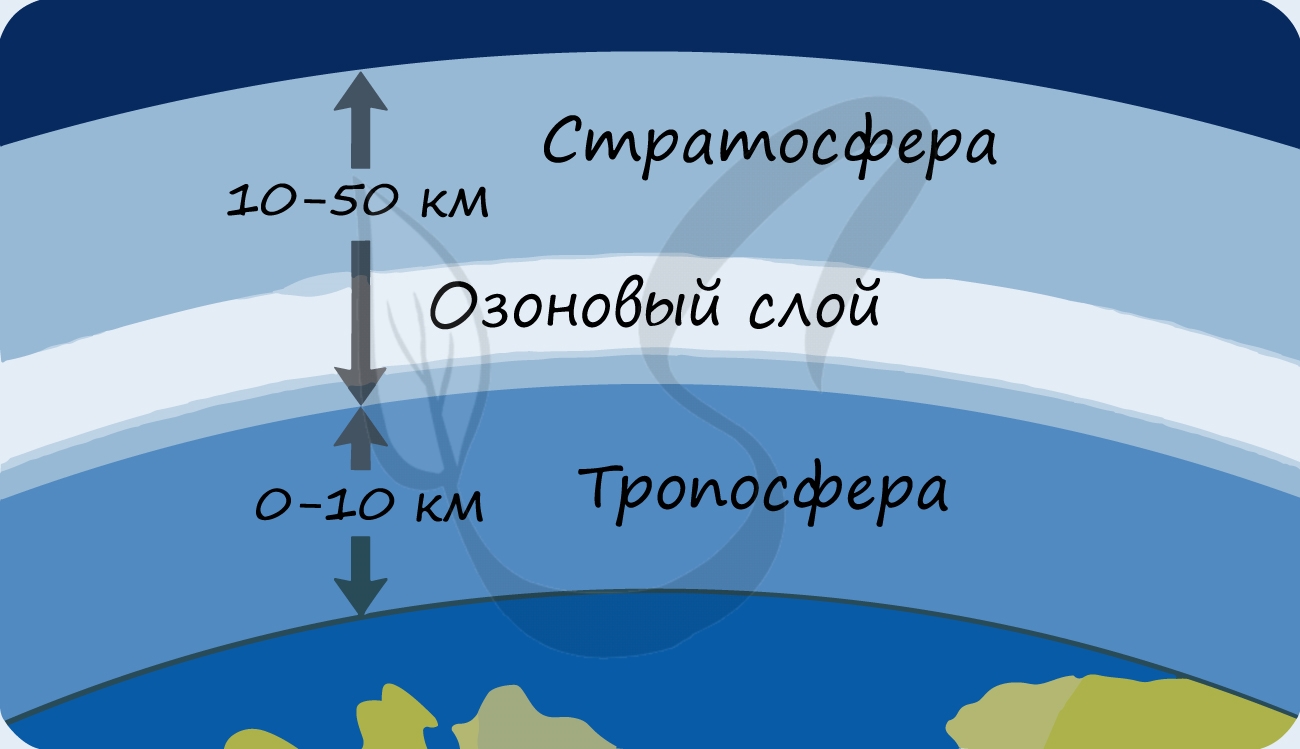
В разделе эволюции мы уже обсуждали, что изначально в составе атмосферы Земли не было кислорода: миллиарды лет назад его начали вырабатывать первые фотосинтезирующие бактерии - сине-зеленые водоросли (цианобактерии). Постепенно кислород накапливался, и со временем на Земле стало возможно аэробное (кислородное) дыхание. Возник озоновый слой, защищающий все живое на нашей планете от губительного ультрафиолета.
- Синтезируют органические вещества, являющиеся пищей для всего живого на планете
- Преобразуют энергию света в энергию химических связей, создают органическую массу
- Растения поддерживают определенный процент содержания O2 в атмосфере, очищают ее от избытка CO2
- Способствуют образованию защитного озонового экрана, поглощающего губительное для жизни ультрафиолетовое излучение

Хемосинтез (греч. chemeia – химия + synthesis - синтез)
Хемосинтез - автотрофный тип питания, который характерен для некоторых микроорганизмов, способных создавать органические вещества из неорганических. Это осуществляется за счет энергии, получаемой при окислении других неорганических соединений (железо- , азото-, серосодержащих веществ).
Хемосинтез был открыт русским микробиологом С.Н. Виноградским в 1888 году. Большинство хемосинтезирующих бактерий относится к аэробам, для жизни им необходим кислород.
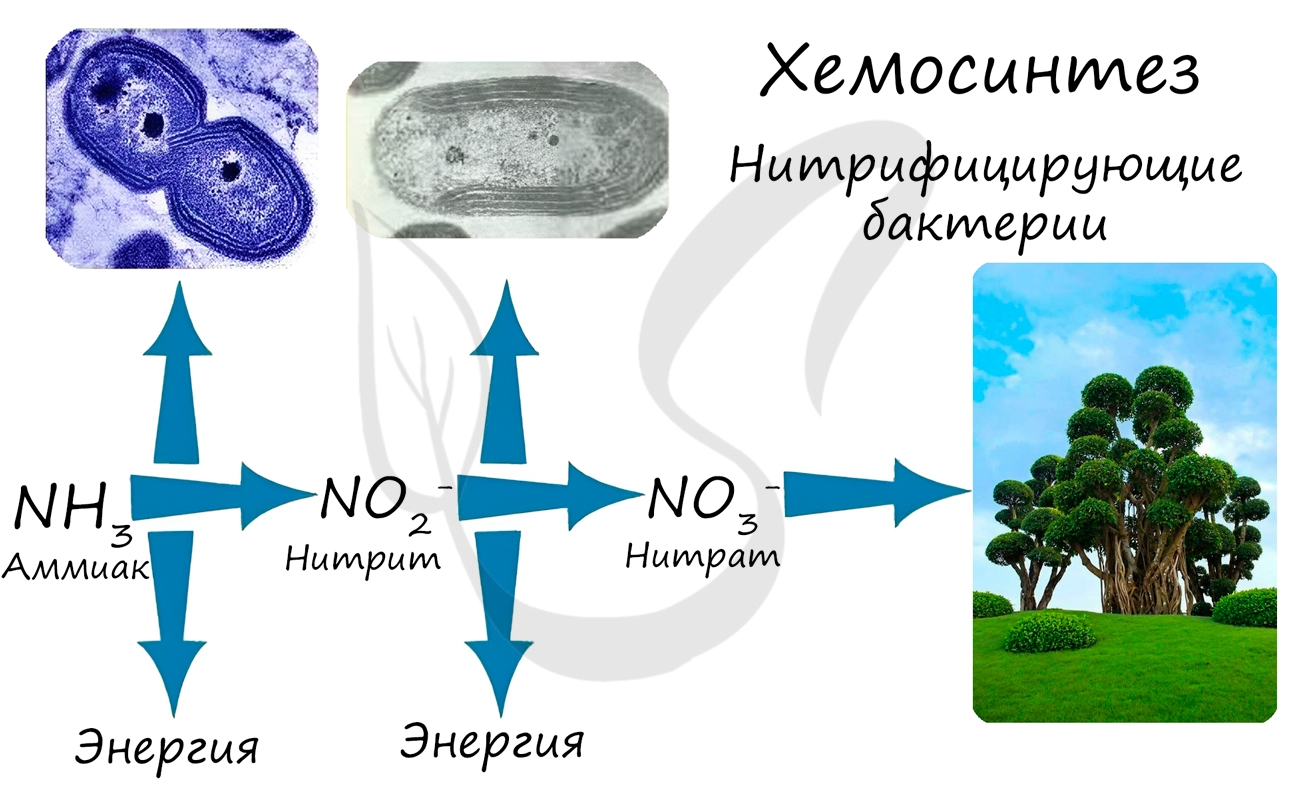
При окислении неорганических веществ выделяется энергия, которую организмы запасают в виде энергии химических связей. Так нитрифицирующие бактерии последовательно окисляют аммиак до нитрита, а затем - нитрата. Нитраты могут быть усвоены растениями и служат удобрением.
- Серобактерии - окисляют H2S --> S 0 --> (S +4 O3) 2- --> (S +6 O4) 2-
- Железобактерии - окисляют Fe +2 -->Fe +3
- Водородные бактерии - окисляют H2 --> H +1 2O
- Карбоксидобактерии - окисляют CO до CO2
Значение хемосинтеза
Хемосинтезирующие бактерии являются неотъемлемым звеном круговорота в природе таких элементов как: азот, сера, железо.
Нитрифицирующие бактерии обеспечивают переработку (нейтрализацию) ядовитого вещества - аммиака. Они также обогащают почву нитратами, которые очень важны для нормального роста и развития растений.
Усвоение нитратов происходит за счет клубеньковых бактерий на корнях бобовых растений, однако важно помнить, что клубеньковые (азотфиксирующие) бактерии, в отличие от нитрифицирующих бактерий, питаются гетеротрофно.
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.

Фотоси́нтез — фотохимические реакции или Фотохимия — это процесс образования углекислого газа и воды на свету при участии фотосинтетических пигментов (хлорофилл у растений, физиологии растений под фотосинтезом чаще понимается фотоавтотрофная функция — совокупность процессов поглощения, превращения и использования энергии квантов света в различных эндэргонических реакциях, в том числе превращения углекислого газа в органические вещества.
Различают оксигенный и аноксигенный типы фотосинтеза. Оксигенный гораздо более широко распространён, его осуществляют растения, цианобактерии и квантов света пигментами, их переход в возбуждённое состояние и передача энергии к другим молекулам фотосистемы. На втором этапе происходит разделение зарядов в реакционном центре, перенос электронов по фотосинтетической электронотранспортной цепи, что заканчивается синтезом сахаров и
Содержание
Пространственная локализация

Хлоропласты в клетках листа
Фотосинтез растений осуществляется в лист. В листе наиболее богата хлоропластами ткань эпидермис, однако большая его часть диффундирует в лист через С4 и CO2 происходит в строме.
Световая (светозависимая) стадия
В ходе световой стадии фотосинтеза образуется высокоэнергетические продукты: АТФ, служащий в клетке источником энергии, и НАДФН, использующийся как восстановитель. В качестве побочного продукта выделяется кислород. В общем роль световых реакций фотосинтеза заключается в том, что в световую фазу синтезируются молекула АТФ и молекулы-переносчики протонов, т.е. НАДФ Н2.
Фотохимическая суть процесса
Второе возбуждённое состояние наиболее высокоэнергетично, нестабильно и хлорофилл за 10 -12 сек переходит с него на первое, с потерей 100 кДж/моль энергии только в виде теплоты. Из первого синглетного и триплетного состояний молекула может переходить в основное с выделением энергии в виде света (флуоресценция) или тепла, с переносом энергии на другую молекулу, либо, поскольку электрон на высоком энергетическом уровне слабо связан с ядром, с переносов электрона на другое соединение.
Вторая возможность реализуется в светособирающих комплексах, первая — в реакционных центрах, где переходящий в возбужденное состояние под воздействием кванта света хлорофилл становится донором электрона ( окислительно-восстановительных потенциалов воды (для полуреакции H20 → O2 E0=+0,82 В) и НАДФ + (E0=-0,32 В). Хлорофилл при этом должен иметь в основном состоянии потенциал больший +0,82 В чтобы окислять воду, но при этом иметь в возбуждённом состоянии потенциал меньший чем −0,32 В чтобы восстанавливать НАДФ + . Одна молекула хлорофилла не может отвечать обоим требованиям. Поэтому сформировалось две фотосистемы и для полного проведения процесса необходимо два кванта света и два хлорофилла разных типов.
Светособирающие комплексы
Хлорофилл выполняет две функции: поглощения и передачу энергии. Более 90 % всего хлорофилла хлоропластов входит в состав светособирающих комплексов (ССК), выполняющих роль антенны, передающей энергию к реакционному центру фотосистем I или II. Помимо хлорофилла в ССК имеются водорослей и цианобактерий — нм. Передача сопровождается некоторыми потерями энергии (10 % от хлорофилла a к хлорофиллу b, 60 % от каратиноидов к хлорофиллу), из-за чего возможна только от пигмента с максимумом поглощения при меньшей длине волны к пигменту с большей. Именно в таком порядке взаимно локализуются пигменты ССК, причём наиболее длинноволновые хлорофиллы находятся в реакционных центрах. Обратный переход энергии невозможен.
ССК растений расположен в мембранах тилакоидов, у цианобактерий основная его часть вынесена за пределы мембран в прикреплённые к ним Основные компоненты электронтранспортной цепи
Фотосистема II
Фотосистема — совокупность ССК, фотохимического реакционного центра и переносчиков электрона. Светособирающий комплекс II содержит 200 молекул хлорофилла a, 100 молекул хлорофилла b, 50 молекул каротиноидов и 2 молекулы феофитина. Реакционный центр фотосистемы II представляет собой пигмент-белковый комплекс, расположенный в тилакоидных мембранах и окружённый ССК. В нём находится димер хлорофилла a с максимумом поглощения при 680 нм (П680). На него в конечном счёте передаётся энергия кванта света из ССК, в результате чего один из электронов переходит на более высокое энергетическое состояние, связь его с ядром ослабляется и возбуждённая молекула П680 становится сильным восстановителем (E0=-0,7 В).
Заполнение электронной вакансии в молекуле П680 происходит за счёт воды. В состав ФС II входит водоокисляющий комплекс, содержащий в активном центре ионы марганца в количестве 4 штук. Для образования одной молекулы кислорода требуется две молекулы воды, дающие 4 электрона. Поэтому процесс проводится в 4 такта и для его полного осуществления требуется 4 кванта света. Комплекс находится со стороны внутритилакоидного пространства и полученные 4 протона выбрасываются в него.
Таким образом, суммарный результат работы ФС II — это окисление 2 молекул воды с помощью 4 квантов света с образованием 4 протонов во внутритилакоидном пространстве и 2 восстановленных пластохинонов в мембране.
b6f или b/f-комплекс
Светособирающий комплекс I содержит примерно 200 молекул хлорофилла.
В реакционном центре первой фотосистемы находится димер хлорофилла a с максимумом поглощения при 700 нм (П700). После возбуждения квантом света он восстанавливает первичный акцептор — хлорофилл a, тот — вторичный ( ферридоксин-НАДФ-редуктазы .
Белок пластоцианин, восстановленный в b6f комплексе, транспортируется к реакционному центру первой фотосистемы со стороны внутритилакоидного пространства и передаёт электрон на окисленный П700.
Циклический и псевдоциклический транспорт электрона
Помимо полного нециклического пути электрона, описанного выше, обнаружены циклический и псевдоциклический.
Суть циклического пути заключается в том, что ферридоксин вместо НАДФ восстанавливает пластохинон, который переносит его назад на b6f комплекс. В результате образуется больший протонный градиент и больше АТФ, но не возникает НАДФН.
При псевдоциклическом пути ферридоксин восстанавливает кислород, который в дальнейшем превращается в воду и может быть использован в фотосистеме II. При этом также не образуется НАДФН.
Темновая стадия
В темновой стадии с участием АТФ и НАДФН происходит восстановление CO2 до глюкозы. Хотя свет не требуется для осуществления данного процесса, он участвует в его регуляции.
С3 — фотосинтез, цикл Кальвина
- Карбоксилирования
- Восстановления
- Регенерация акцептора CO2
На первой стадии к рибулозо-1,5-бифосфату присоединяется CO2 под действием фермента рибулозобисфосфат-карбоксилаза/оксигеназа ( С4 — фотосинтез
При низкой концентрации растворённого в строме CO2 Rubisco катализирует реакцию окисления рибулёзо-1,5-бифосфата и его распад на 3-фосфоглицериновую кислоту и САМ фотосинтез
При CAM (Crassulaceae acid metabolism) фотосинтезе происходит разделение ассимиляции CO2 и цикла Кальвина не в пространстве как у С4, а во времени. Ночью в вакуолях клеток по аналогичному вышеописанному механизму при открытых устьицах накапливается малат, днём при закрытых устьицах идёт цикл Кальвина. Этот механизм позволяет максимально экономить воду, однако уступает в эффективности и С4, и С3. Он оправдан при Значение фотосинтеза
Фотосинтез является главным входом неорганического углерода в биологический цикл. Весь кислород атмосферы биогенного происхождения и является его побочным продуктом. Формирование окислительной атмосферы полностью изменило состояние земной поверхности, сделало возможным появление дыхания, а в дальнейшем, после образования Изучение фотосинтеза
Хлорофиллы был впервые выделены в 1818 П. Ж. Пелетье и Ж. Кавенту. Разделить пигменты и изучить их по отдельности удалось М. С. Цвету с помощью созданного им метода К. А. Тимирязевым, он же, развивая положения Майера, показал что именно поглощенные позволяют повысить энергию системы, создав вместо слабых связей С-О и О-Н высокоэнергетические С-С (до этого считалось что в фотосинтезе используются жёлтые лучи, не поглощаемые пигментами листа). Сделано это было благодаря созданному им методу учёта фотосинтеза по поглощённому CO2, в ходе экспериментов по освещению растения светом разных длин волн (разного цвета) оказалось что интенсивность фотосинтеза совпадает со спектром поглощения хлорофилла.
Окислительно-восстановительную сущность фотосинтеза (как оксигенного, так и аноксигенного) постулировал Корнелис ван Ниль . Это означало что кислород в фотосинтезе образуется полностью из воды, что экспериментально подтвердил в 1941 А. П. Виноградов в опытах с изотопной меткой. В 1937 Роберт Хилл установил что процесс окисления воды (и выделения кислорода), а также ассимиляции CO2 можно разобщить. В 1954-1958 Д. Арнон установил механизм световых стадий фотосинтеза, а сущность процесса ассимиляции CO2 была раскрыта Мельвином Кальвином с использованием изотопов углерода в конце 1940-х, за эту работу в 1961 ему была присуждена Нобелевская премия.
В 1955 была выделена и очищена Rubisco. С4 фотосинтез был описан Ю. С. Карпиловым в 1960 и М. Д. Хэтчем и К. Р. Слэком в 1966.
Читайте также:
- Архангельский с и лекции по научной организации учебного процесса в высшей школе
- 3 сформулируйте 3 4 темы которые по вашему мнению будут актуальны для оэр в доу
- Польза молока для детей школьного возраста
- Информационный материал для родителей в детском саду день матери
- Совместные правила общения детей дома и в школе